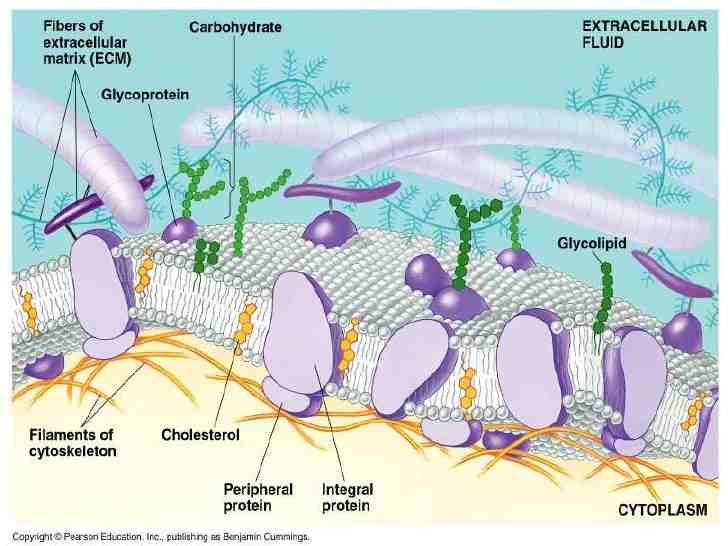
ساختار غشاء
غلاف میلین دارای کمترین پروتئین (۲۰%) و بیشترین لیپید (۷۵%) است. غشای داخلی میتوکندری دارای بیشترین پروتئین (۷۵%) و کمترین لیپید (۲۳%) و کربوهیدرات (۲%) است. بیشترین کربوهیدرات غشایی در اریتروسیت ها وجود دارد.
لیپیدهای غشاء:
اسفنگومیلین و فسفاتیدیل کولین اساسا در لایه خارجی غشاء و فسفاتیدیل اتانول آمین، فسفاتیدیل سرین، فسفاتیدیل اینوزیتول اغلب در لایه داخلی غشاء یافت میشوند.
ماکروفاژها حاوی گیرندههای فسفاتیدیل سرینی هستند که به سلولهای آشکار کننده فسفاتیدیل سرین متصل شده و سبب تخریب آن ها میگردند. افزایش سن گلبولهای قرمز به همراه آشکار شدن فسفاتیدیل سرین در سطح غشای پلاسمایی، نشانهای برای سیستم ماکروفاژی جهت خارج کردن آنها از سیستم گردش خون است. آشکار شدن فسفاتیدیل سرین در سلولهای قرمز در شرایط بیماری مثل مسمومیت با سرب و اورمی در بیماران مبتلا به نقص کلیوی، منجر به کاهش طول عمر سلولهای قرمز می شود. آنزیم سمبلاز مسئول انتقال فسفاتیدیل سرین از نیمه داخلی غشای پلاسمایی به نیمه بیرونی میباشد. سمبلاز در سلولهای سالم غیرفعال است. در ابتدای تحریک پلاکتها، فسفاتیدیل سرین به کمک آنزیم فلیپاز به میزان مختصر به سطح خارجی غشاء تغییر مکان داده و در اینجا فسفاتیدیل سرین، عوامل دخیل در انعقاد خون را فعال میکند.
غشاهای پلاسمایی سلولهای جانوری غنی از کلسترول بوده در حالی که غشای داخلی میتوکندری دارای کلسترول بسیار پایین میباشد. در سلولهای جانوری کلسترول تنظیم کننده کلیدی سیالیت غشاء بوده و در کنترل ساختارهای ریز غشای پلاسمایی شرکت می کند. افزایش کلسترول منجر به کاهش سیالیت غشاء شده و کاهش کلسترول میزان سیالیت را افزایش می دهد. به طور کلی سیالیت غشاء به عواملی چون میزان اسیدهای چرب غیر اشباع موجود در فسفولیپیدهای غشا، میزان کلسترول و درجه حرارت بستگی دارد.
در بیماری های کبدی مانند سیروز و همچنین در کم خونی سلول خاردار (آکانتوسیتوز) مقادیر کلسترول غشای گلبول های قرمز افزایش مییابد. در آبتا لیپوپروتئینمی محتوای اسفنگومیلین غشای سلولها افزایش و میزان فسفاتیدیل کولین کاهش مییابد و در نتیجه سیالیت غشای سلول.ها نیز دچار کاهش میشود.
پروتئینهای غشاء
 پروتئین های سراسری:
پروتئین های سراسری:
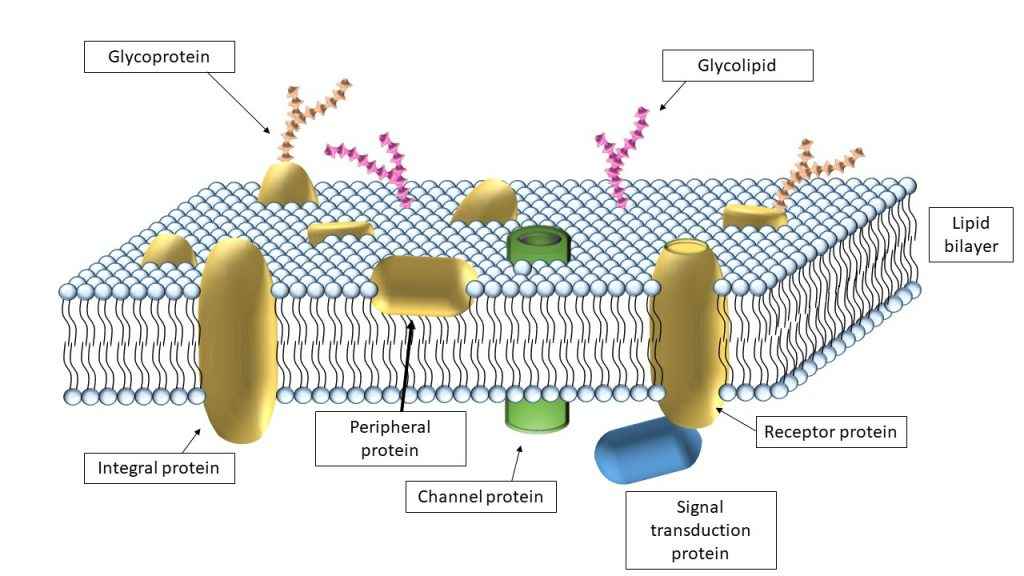 پروتئین های سراسری:
پروتئین های سراسری:پروتئین های غشایی سراسری ارتباط محکمی با لیپیدهای غشا برقرار کرده و تنها به وسیله ترکیباتی نظیر دترجنتها، حلالهای آلی، یا دناتورهکنندهها میتوان آنها را جدا کرد. اتصال محکم پروتئینهای سراسری به غشاها، نتیجه میانکنشهای آبگریز بین لیپیدهای غشایی و دُمَینهای آبگریز پروتئین میباشد. در همه پروتئینهای گذرنده از غشاء دارای، دُمَین گذرنده از غشاء دارای یک یا تعداد بیشتری مارپیچ آلفا یا صفحات چند تایی بتا تشکیل شده است.
باکتریورودوپسین یک پروتئین تک زنجیره گذرنده از غشا در یک نوع باکتری به نام هالوباکتریوم سالیناروم میباشد که دارای هفت ناحیه داخلی بسیار آبگریز بوده و هفت بار عرض غشای دو لایه را طی میکند. باکتریورودوپسین یک پمپ پروتئین وابسته به نور است که به صورت متراکم و با آرایش منظم در غشاء قرار گرفته است.
پروتئینهای محیطی
پروتئینهای غشایی محیطی از طریق میانکنشهای الکتروستاتیک و پیوند هیدروژنی با دُمَینهای آبدوست پروتئینهای سراسری و گروههای قطبی لیپیدهای غشایی، با غشاء در ارتباط هستند. پروتئینهای محیطی غشاء را میتوان با استفاده از محلولهایی با قدرت یونی بالا جدا نمود.
پروتئینهای لنگری متصل به لیپید
پروتئینهای گلیپه
از اتصال زنجیر پلی پپتیدی از طریق یک رابط فسفو اتانول آمینی و الیگو ساکاریدی به فسفاتیدیل اینوزیتول گلیکوزیله غشایی تشکیل می شوند. لذا به آن لنگر فسفاتیدیل اینوزیتول گلیکوزیله (GPI) گفته می شود.
پروتئین های آسیله:
با برقراری پیوند کوالان آمیدی یا استری بین ریشه جانبی آمینو اسیدهای سیستئین، سرین، ترئونین زنجیر پلی پپتیدی و گروه کربوکسیل اسید چرب مانند میریستیک اسید، پالمیتیک اسید، استئاریک اسید، اولئیک اسید تشکیل می شوند. میریستیک اسید توسط یک پیوند آمیدی به یک گلیسین در ناحیه N-ترمینال پروتئین متصل می شود.
پروتئین های پرنیله:
با برقراری پیوند سولفیدی یا تیواتری بین گروه سولفیدریل یک ریشه سیستئین در توالیCAAX-Cys در انتهای کربوکسیل زنجیر پلی پپتیدی و کربن انتهایی یک زنجیر پلی ایزوپرنوئیدی مانند فارنسیل و ژرانیل ژرانیل تشکیل می شود. X موجود در توالی مذکور نوع ایزوپرنوئید را مشخص می کند. گلوتامین، متیونین و سرین برای اتصال فارنسیل و لوسین برای اتصال ژرانیل ژرانیل مورد نیاز است.
پروتئین های لنگری آسیله و پرنیله در لایه داخلی غشاء و پروتئین های لنگری گلیپه در لایه خارجی غشاء وجود دارند.
کانال های غشاء:
کانال های پورین:
کانال های پورین دارای یک توالی گذار غشایی شامل رشته های بتا است که ساختمان بشکه-بتا را به همراه یک کانال پر از آب که قطری حدود ۶/۰ تا ۳ نانومتر دارد تشکیل می دهد. این ساختارها در غشای خارجی میتوکندری، برخی از باکتری ها و پلاستیدها وجود دارند. چندین پورین باکتریایی با ساختار بشکه-بتا وجود دارد که دارای ۱۶ رشته غیرموازی بوده و توسط پیوند هیدروژنی میان نزدیک ترین مولکول های مجاور در امتداد زنجیره به هم متصل شده اند.
کانال های سدیمی دریچه دار ولتاژی:
این کانال ها دو دریچه دارند: یکی در خارج که دریچه فعال سازی نام دارد و دیگری در داخل که دریچه غیر فعال سازی نامیده می شود. این کانال ها توسط تتردوتوکسین و ساکسی توکسین مهار می شوند.
کانال های پتاسیمی دریچه دار ولتاژی:
این کانال ها فقط یک دریچه فعال سازی در سطح داخل سلولی غشاء دارند. این کانال ها توسط دندروتوکسین مهار می شوند.
کانال های کلسیمی:
گیرنده های دی هیدروپیریدینی (DHP) از نوع کانال های کلسیمی دریچه دار ولتاژی می باشند که در غشای توبول های T عضله اسکلتی و قلبی وجود دارند. یک نوع از کانال کلسیمی دریچه دار لیگاندی در شبکه آندوپلاسمی/ شبکه سارکوپلاسمی (ER/SR) عضله اسکلتی و قلبی وجود دارد که گیرنده رایانودینی (RYR) نام دارد. کانال های RYR در عضله قلبی، به وسیله کلسیم وارد شده از طریق کانال های غشای پلاسمایی به داخل سلول باز می شوند. این نوع آزاد شدنCa++ ، رهاسازیCa++ القاء شده با Ca++ نامیده می شود. در مقابل، RYR عضله اسکلتی از طریق برهمکنش مستقیم پروتئین-پروتئین با کانال های Ca++ حساس به ولتاژ باز می شوند.
اختلالات کانال های یونی:
پارامیوتونی مادرزادی و فلج دوره ای هیپرکالمیک:
به دلیل جهش در کانال های سدیمی دریچه دار ولتاژی
سندرم QT طولانی:
به دلیل جهش در کانال های سدیمی و پتاسیمی دریچه دار ولتاژی
فلج دوره ای خانوادگی:
به دلیل جهش در کانال های سدیمی دریچه دار ولتاژی و کلسیمی دریچه دار ولتاژی
فلج دوره ای هیپوکالمیک:
به دلیل جهش در کانال های کلسیمی ولتاژی
هیپرترمی بدخیم:
به دلیل جهش جهش در گیرنده رایانودینی در عضله اسکلتی
سندرم میاستنی لامبرت-ایتون (LEMS):
یک بیماری خودایمنی است که در آن آنتی بادی ضد کانال های کلسیمی دریچه دار ولتاژی وجود دارد.
پروتئین های ناقل:
ATPase Na+ / K+:
این پمپ از دو زیر واحد تشکیل شده است: یک زیر واحد آلفا (۱۲ کیلو دالتون) که دارای ۸ مارپیچ عبور کننده از غشاء و نیز واجد جایگاهATPase در سمت سیتوزولی غشاء می باشد و یک زیر واحد بتا (۳۵ کیلو دالتون) که دارای یک مارپیچ عبور کننده از غشاء و به شکل گلیکوزیله می باشد. بخش خارج سلولی زیر واحد کوچک جایگاه اتصال استروئیدهای کاردیوتونیک نظیر دیژیتالیس (دیگوکسین و اوآبائین) است که این پمپ را مهار می نمایند. این پمپ سبب انتقال دو یون K+ به داخل سلول با اتصال به زیر واحد کوچک و سه یون Na+ به خارج سلول با اتصال به زیر واحد بزرگ می شود. اتصال سه یون Na+ در شکل دفسفریله انتقال دهنده و در داخل سلول و اتصال دو یونK+ در شکل فسفریله آن و در خارج سلول رخ می دهد.
Ca++ ATPase شبکه سارکوپلاسمی عضله (SERCA):
این پمپ دو یون Ca++ را بر خلاف گرادیان بزرگ غلظت، از سیتوزول به لومن شبکه سارکوپلاسمی جا به جا می کند و این عمل را به وسیله یک ناقل ناهمسو و در تبادل با دو یون H+ و به ازاء هیدرولیز یک ATPانجام می دهد. این ناقل توسط سطوح Ca++ سیتوپلاسمی و به واسطه کالمودولین که یک پروتئین متصل شونده بهCa++ است تنظیم می شود.
H+ ATPase:
به طور عمده در غشاء سلول های پاریتال معده و توبول های دیستال انتهایی و مجاری جمع کننده قشری کلیه ها وجود دارد.
ناقل ناهمسوی Na+/H+:
در غشای پلاسمایی تقریبا تمام سلول های بدن وجود دارد و در تنظیم pH داخل سلولی و مایعات دخالت دارد.
ناقل ناهمسوی Ca++/Na+:
در غشای سارکوپلاسمی فیبرهای عضلانی وجود دارد که از تمایل بالای Na+ برای ورود به داخل سلول در جهت خروج یون Ca++از سلول استفاده می کند.
ناقل ناهمسوی H+ K+ ATPase:
در سطح سلول های پاریتال معده وجود دارد و به عنوان یک پمپ، یون های H+ را به داخل معده ترشح می کند.
ناقلین غشای داخلی میتوکندری:
ناقل فسفات: فسفات را به همراه پروتون وارد ماتریکس می کند (ناقل فسفات همچنین می تواند به عنوان ناقل ناهمسوی فسفات-هیدروکسیل، فسفات را به ماتریکس و OH- را به خارج انتقال دهد).
ناقل منوکربوکسیلات: پیروات را به ماتریکس وOH- را به خارج انتقال می دهد.
ناقل دی کربوکسیلات: فسفات را به ماتریکس و مالات، سوکسینات و فومارات را به خارج انتقال می دهد.
ناقل تری کربوکسیلات: مالات را به ماتریکس و سیترات را به همراه H+ به خارج انتقال می دهد.
ناقل نوکلئوتید آدنینی: ADP را به ماتریکس وATP را به خارج انتقال می دهد.
ناقل آسپارتات-گلوتامات: گلوتامات را به ماتریکس و آسپارتات را به خارج انتقال می دهد.
ناقل مالات-آلفا کتوگلوتارات: مالات را به ماتریکس و آلفا کتوگلوتارات را به خارج انتقال می دهد.
اتصالات بین سلولی:
این اتصالات شامل اتصالات محکم یا کمربندی (از طریق رشته های اکتین)، اتصالات دسموزوم (با واسطه کادهرین ها و رشته های حد واسط مانند سیتوکراتین)، اتصالات همی دسموزوم و اتصلات منفذ دار نکسوس (از طریق پروتئین های کانکسون) می باشند. اتصالات محکم در جدار روده و رگ ها، دسموزوم ها در پوست و سلول های ماهیچه قلبی، و اتصالات منفذدار در ماهیچه صاف، ماهیچه قلبی و سلول های عصبی دیده می شوند.
چرا غشاء پلاسمایی به پروتئین های مختلف نیاز دارد؟
پروتئین های غشای پلاسمایی در چند گروه عملکردی بزرگ قرار می گیرند.

– برخی از پروتئین های غشاء، سلول را به ماده زمینه ای متصل نکه می دارند. برای مثال اینتگرینها به ماده زمینهای خارج از سلول و به طور همزمان به ریز رشتههای درون سلولی نیز اتصال دارند. بسیاری از سلولهای جانوری بوسیله ماتریکس خارج سلولی احاطه شدهاند که توسط خود آنها ترشح شده است. این ماتریکس شامل ژلی از پروتئین.های رشته ای و کربوهیدرات هاست.
مهم ترین پروتئین ساختاری ماتریکس خارج سلولی، کلاژن است که رشته های بسیار محکمی را بوجود می آورد. از دیگر گلیکوپروتئینهای موجود در ماتریکس خارج سلولی فیبرونکتینها هستند که به حفظ سازمان ماتریکس و نیز ایجاد اتصال سلول ها به آن کمک می کنند. فیبرونکتین ها به گیرنده های پروتئینی که از غشاء بیرون زده اند متصل می گردند. اینتگرینها پروتئینهایی هستند که به عنوان گیرندههای غشایی عمل می کنند. این پروتئین ها بسیاری ار مسیرهای پیام رسانی سلول را فعال می کنند تا اطلاعات را از سلول به ماتریکس خارج سلولی انتقال دهند. اینتگرین ها در حرکت سلولی و در سازمان دهی اسکلت سلولی اهمیت دارند، به طوری که سلول ها به این وسیله شکل معینی را به خود می گیرند. در بسیاری از سلول ها، اینتگرین ها ماتریکس خارج سلول را به ریز رشته های اسکلت سلولی داخلی وصل می کنند. وقتی سلول ها این اتصالات را نداشته باشند فرایند آپوپتوز صورت می گیرد. سلول های سرطانی ظاهراً نیازی به وصل شدن به ماتریکس خارج سلولی ندارند در واقع این خاصیت را از دست داده اند.
– بسیاری از پروتئین های غشا در انتقال مواد از غشاء نقش دارند. بسیاری از آنها کانالهایی را ایجاد می کنند که به طور انتخابی اجازه عبور یون ها یا مولکول های خاصی را می دهند. برخی دیگر از این پروتئین ها، پمپ هایی را می سازند که برای عبور فعالانه مواد حل شده از عرض غشا انرژی مصرف می کنند.
– برخی پروتئین های غشایی آنزیم هایی هستند که واکنش های نزدیک سطح سلول را کاتالیز می کنند.
– برخی دیگر از پروتئین های غشا، گیرنده هایی هستند که اطلاعات را از سلول های دیگر دریافت می کنند. برای مثال پیام های هورمونی را از سلول های درون ریز دریافت می کنند. این اطلاعات توسط سیستم تبدیل و انتقال پیام به درون سلول منتقل می شوند.
– برخی پروتئین های غشاء به صورت برچسب های شناسایی عمل می کنند که سایر سلول ها آن ها را تشخیص می دهند. سلول هایی که همدیگر را شناسایی می کنند ممکن است برای تشکیل بافت به هم متصل شوند. سلول های انسان گیرنده های شاخصی دارند که مشخصه آن ها به عنوان یک ویژگی منحصر به فرد است. برخی سلول ها پروتئین های سطحی یا آنتی ژن های سلول باکتری را به عنوان یک عامل خارجی، مورد شناسایی قرار می دهند.
– برخی دیگر از پروتئین های غشایی، اتصالات بین سلول های مجاور را بوجود می آورند. این پروتئین ها ممکن است به عنوان نقطه اتکایی برای عناصر تشکیل دهنده اسکلت سلولی نیز عمل کنند.

در مورد فصل ۱ مطالب بسیار مفیدی بود
عالیه👌
بسیار عالی بود
ممنون از مطلب بسیار آموزنده و جالب شما
سلام خسته نباشید مطلب آموزنده و بدردبخور برای همین امسال
مرسی از زحماتتون🌷
خسته نباشید به عوامل سایت عالی👌