سیستم کولینرژیک و هیستامینرژیک؛ نوروفارماکولوژی مولکولی؛ نوروساینس بالینی

ACETYLCHOLINE
Synthetic and Degradative Pathways
استیل کولین
مسیرهای سنتزی و تخریبی
ACh is synthesized in a reversible reaction in which an acetyl group is transferred from acetyl coenzyme A (COA) to choline by the enzyme choline acetyltransferase (ChAT) (6-16). Its synthesis is regulated by the rate-limiting availability of choline. Choline is transported into neuronal terminals, either in its free form or bound to membrane phospholipids, by distinct high-affinity and low-affinity transport mechanisms (6-17). Most ACh is synthesized in terminals, which are rich in ChAT, and also in mitochondria, which are the site of acetyl CoA synthesis.
ACh در یک واکنش برگشت پذیر سنتز میشود که در آن یک گروه استیل توسط آنزیم کولین استیل ترانسفراز (ChAT) از استیل کوآنزیم آ (COA) به کولین منتقل میشود (16-6). در دسترس بودن کولین به عنوان محدودکننده سرعت، سنتز آن را تنظیم میکند. کولین یا به شکل آزاد یا متصل به فسفولیپیدهای غشایی به پایانه های عصبی منتقل میشود که توسط مکانیسمهای انتقالی با میل ترکیبی زیاد و میل ترکیبی کم مجزایی صورت میگیرد (6-17). بیشتر ACh در پایانهها که غنی از ChAT است و همچنین در میتوکندریها که محل سنتز استیل CoA هستند، سنتز میشود.

6-16 Biosynthetic pathway of acetylcholine. Choline and acetyl coenzyme A produce acetylcholine in a reversible reaction catalyzed by choline acetyltransferase (CHAT). After it is synthesized, acetylcholine is rapidly metabolized by acetylcholinesterase into choline and free acetate. Subsequently both may be recycled to produce more acetylcholine Me = CH3 (methyl moiety).
6-16 مسیر بیوسنتزی استیل کولین. کولین و استیل کوآنزیم A در یک واکنش برگشت پذیر استیل کولین تولید میکنند که توسط استیل کولین ترانسفراز (CHAT) کاتالیز میشود. پس از سنتز، استیل کولین به سرعت توسط استیل کولین استراز به کولین و استات آزاد متابولیزه میشود. بعد از آن هر دو ممکن است بازیافت شوند تا استیل کولین Me = CH3 (بخش متیل) بیشتری تولید شود.

6-17 Model of a cholinergic synapse. Several interesting features of this synapse distinguish it from monoamine synapses. First, the most significant mechanism by which the action of acetylcholine is terminated is not by a (reuptake) transporter, but by a highly active enzyme, acetylcholinesterase (AChE). Second, the vesicular transporter for acetylcholine is distinct from the vesicular monoamine transporter (VMAT). Third, the membrane transporter does not return neurotransmitter to the presynaptic neuron but rather transports its metabolite (choline). Fourth, the presence of mitochondria is especially important because these organelles supply the acetyl coenzyme A necessary for acetylcholine synthesis. Finally, both G protein- coupled receptors, such as muscarinic subtypes M1 to M5, and ligand-gated ion channels, such as nicotinic receptors (nAChRs), may be present.
6-17 مدل سیناپس کولینرژیک. چندین ویژگی جالب این سیناپس آن را از سیناپسهای مونوآمین متمایز میکند. اول، مهم ترین مکانیسمی که توسط آن عمل استیل کولین خاتمه مییابد توسط یک ترانسپورتر (بازجذب) نیست، بلکه توسط یک آنزیم بسیار فعال یعنی استیل کولین استراز (AChE) است. دوم، ترانسپورتر برای استیل کولین از ترانسپورتر وزیکولی مونوآمین (VMAT) متمایز است. سوم، ترانسپورتر غشایی، نوروترانسمیتر را به نورون پیش سیناپسی باز نمیگرداند، بلکه متابولیت آن (کولین) را منتقل میکند. چهارم، حضور میتوکندریها از اهمیت ویژه ای برخوردار است زیرا این اندامکها استیل کوآنزیم A لازم برای سنتز استیل کولین را تامین میکنند. در نهایت، هر دو گیرندههای جفتشده با پروتئین G، مانند زیرگروههای موسکارینی M1 تا M5، و کانالهای یونی دریچهدار با لیگاند، مانند گیرندههای نیکوتین (nAChRs)، ممکن است وجود داشته باشند.
The synaptic actions of ACh are terminated by the enzyme acetylcholinesterase (AChE), which hydrolyzes Ach into acetate and choline (6-17). AChE is an extraordinarily efficient enzyme that is capable of hydrolyzing 1000 ACh molecules per second per molecule of enzyme. This enzyme occurs in the cytoplasm and in the outer cell membrane; thus, it can metabolize ACh both intracellularly and extracellularly. Anticholinesterases, which inhibit AChE, cause released ACh to accumulate extracellularly, producing excessive stimulation of ACh receptors throughout the nervous system. Reversible inhibitors, such as physostigmine and neostigmine, inactivate AChE for as many as 4 hours and are used clinically to treat glaucoma, myasthenia gravis (Chapter 12), and smooth muscle dysfunction of the bladder and intestines. Unlike physostigmine, neostigmine cannot enter the brain because it is a quaternary ammonium compound and thus is too highly charged to cross the blood-brain barrier (Chapter 2). Centrally acting anticholinesterases, such as tacrine and donepezil, are used to increase central ACh concentrations in patients with Alzheimer disease (see below). Irreversible anticholinesterases completely inhibit ACh breakdown. Restoration of AChE activity requires new AChE synthesis. These irreversible inhibitors are used as insecticides, and are highly toxic if ingested by humans. These agents were a major class of nerve gases. Just before World War II, first German and then Allied scientists began to explore potential uses for organophosphate agents in chemical warfare. Their efforts resulted in the development of several lethal compounds, including sarin, soman, and tabun, which can cause death within 5 minutes of exposure. The primary cause of death is respiratory failure, which typically is preceded by cognitive impairment and severe autonomic symptoms. Treatment for exposure to these toxins involves combined administration of a muscarinic receptor antagonist, such as atropine, with the AChE antagonist, pralidoxime, which paradoxically restores AChE function.
فعالیتهای سیناپسی ACh توسط آنزیم استیل کولین استراز (AChE) خاتمه مییابد، که Ach را به استات و کولین تجزیه میکند (6-17). AChE یک آنزیم فوق العاده کارآمد است که هر مولکول آنزیم قادر است در هر ثانیه 1000 مولکول ACh را تجزیه کند. این آنزیم در سیتوپلاسم و در غشای سلولی خارجی وجود دارد. بنابراین، میتواند ACH را هم به صورت درون سلولی و هم خارج سلولی متابولیزه کند. آنتی کولین استرازها که AChE را مهار میکنند باعث تجمع ACH آزاد شده در خارج سلولی میشوند و باعث تحریک بیش از حد گیرندههای ACh در سراسر سیستم عصبی میشوند. مهارکنندههای برگشتپذیر، مانند فیزوستیگمین و نئوستیگمین، AChE را تا 4 ساعت غیرفعال میکنند و از نظر بالینی برای درمان گلوکوم، میاستنی گراویس (فصل 12)، و اختلال عملکرد عضلات صاف مثانه و روده استفاده میشوند. بر خلاف فیزوستیگمین، نئوستیگمین نمیتواند وارد مغز شود زیرا یک ترکیب آمونیوم چهارتایی است و بنابراین برای عبور از سد خونی مغزی بسیار باردار است (فصل 2). آنتی کولین استرازهای با اثر مرکزی، مانند تاکرین و دونپزیل، برای افزایش غلظت ACH مرکزی در بیماران مبتلا به بیماری آلزایمر استفاده میشوند (به زیر مراجعه کنید). آنتی کولین استرازهای برگشت ناپذیر تجزیه ACh را کاملاً مهار میکنند. بازیابی فعالیت AChE نیازمند سنتز جدید AChE است. این مهارکنندههای برگشت ناپذیر به عنوان حشره کش استفاده میشوند و اگر توسط انسان بلعیده شوند بسیار سمی هستند. این عوامل دسته عمده ای از گازهای اعصاب بودند. درست قبل از جنگ جهانی دوم، ابتدا دانشمندان آلمانی و سپس دانشمندان متفقین شروع به کشف کاربردهای بالقوه عوامل ارگانوفسفره در جنگهای شیمیایی کردند. تلاشهای آنها منجر به تولید چندین ترکیب کشنده از جمله سارین، سومان و تابون شد که میتوانند در عرض 5 دقیقه پس از مواجهه باعث مرگ شوند. علت اصلی مرگ نارسایی تنفسی است که معمولاً با اختلالات شناختی و علائم شدید اتونوم پیش میآید. درمان قرار گرفتن در معرض این سموم شامل تجویز ترکیبی یک آنتاگونیست گیرنده موسکارینی، مانند آتروپین، با آنتاگونیست AChE، پرالیدوکسیم است که به طور متناقضی عملکرد AChE را بازیابی میکند.
Functional Anatomy
آناتومی کاربردی
ACh is synthesized by widely projecting neurons with cell bodies in the basal forebrain and brainstem (6-18) and by interneurons in striatum.
ACH توسط نورونهایی که جسم سلولی آنها در قاعده مغزپیشین و ساقه مغز (6-18) و اینترنورونهایی در جسم مخطط ساخته میشوند و دارای پروجکتینگ گستردهای هستند.
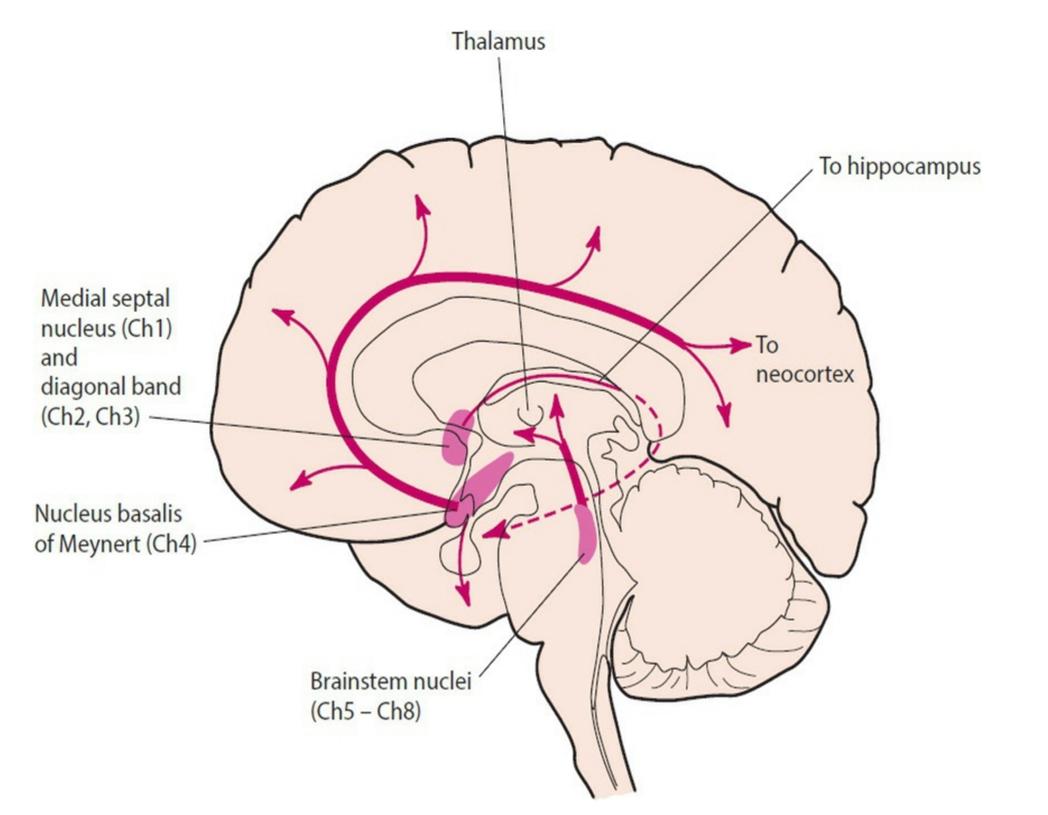
6-18 Cholinergic projections in the CNS. Eight small nuclei in the basal forebrain and brainstem, designated Ch1 to 8 (for cholinergic nuclei 1–8) in the rat, supply the cholinergic innervation in the brain. The basal forebrain cholinergic nuclei are composed of the medial septal nucleus (Ch1), the vertical nucleus of the diagonal band (Ch2), the horizontal limb of the diagonal band (Ch3), and the nucleus basalis of Meynert (Ch4). Brainstem cholinergic nuclei include the pedunculopontine nucleus (Ch5), the laterodorsal tegmental nucleus (Ch6), the medial habenula (Ch7), and the parabigeminal nucleus (Ch8).
6-18 پروجکتینگهای کولینرژیک در CNS. هشت هسته کوچک در قاعده مغز پیشین و ساقه مغز، که از Ch1 تا 8 (برای هستههای کولینرژیک 1-8) در موش صحرایی مشخص شدهاند، عصبدهی کولینرژیکی را در مغز تامین میکنند. هستههای کولینرژیک قاعده ای مغز پیشین از هسته سپتوم میانی (Ch1)، هسته عمودی نوار مورب (Ch2)، اندام افقی نوار مورب (Ch3) و هسته بازالیس ماینرت (Ch4) تشکیل شده اند. هستههای کولینرژیک ساقه مغز شامل هسته پدانکولوپونتین (Ch5)، هسته لاترودورسال تگمنتال (Ch6)، هابنولا میانی (Ch7) و هسته پارابیژمینال (Ch8) میباشد.
Widely projecting cholinergic neurons arise from eight nuclei that are clustered in two areas, the basal forebrain and the upper brainstem. The basal forebrain cholinergic nuclei are composed of the medial septal nucleus, the diagonal band, and the nucleus basalis of Meynert. The dense innervation of the sensory and limbic cortices arising predominantly from the diagonal band and nucleus basalis is thought to influence arousal, cortical responsiveness to sensory input, and emotional states. Cholinergic neurons of the diagonal band and the nucleus basalis also innervate regions of the hippocampus and cerebral cortex involved in learning and memory. Because these basal forebrain cholinergic neurons tend to undergo early and severe degeneration in Alzheimer disease, likely contributing to cognitive impairment, they have been investigated as therapeutic targets. These neurons are dependent on nerve growth factor (NGF) for trophic support (Chapter 8); accordingly, NGF and drugs aimed at NGF signaling pathways have been tested as treatments but have not proven successful. It is because of the loss of ACh in the hippocampus and cerebral cortex that AChE inhibitors have been used in Alzheimer disease. Unfortunately, they are only modestly efficacious; at best they delay symptomatic deterioration for a few months, but do not slow neurodegeneration and do not produce dramatic clinical improvement (Chapter 18).
نورونهای کولینرژیک بهطور گسترده از هشت هسته که در دو ناحیه، قاعده مغز پیشین و بالای ساقهی مغز جمع شدهاند، به وجود میآیند. هستههای کولینرژیک قاعده ای مغز پیشین از هسته سپتوم میانی، نوار مورب و هسته بازالیس ماینرت تشکیل شده اند. تصور میشود که عصبدهی فراوان قشر حسی و لیمبیک که عمدتاً از نوار مورب و هسته بازالیس ناشی میشود بر برانگیختگی، پاسخ قشری به ورودی حسی و حالات عاطفی تأثیر میگذارد. نورونهای کولینرژیک نوار مورب و هسته بازالیس نیز بخشهایی از هیپوکامپ و قشر مغز را که در یادگیری و حافظه دخیل هستند، عصب دهی میکنند. از آنجایی که این نورونهای کولینرژیک قاعده مغز پیشین در بیماری آلزایمر دچار تحلیل اولیه و شدید میشوند، آنها به عنوان اهداف درمانی مورد بررسی قرار گرفتهاند زیرا احتمالاً به اختلال شناختی کمک میکنند. این نورونها برای حمایت تغذیه ای به فاکتور رشد عصبی (NGF) وابسته هستند (فصل 8). بر این اساس، NGF و داروهایی که در مسیرهای سیگنال دهی NGF قرار دارند به عنوان درمان آزمایش شده اند اما موفقیت آمیز نبوده اند. به دلیل از بین رفتن ACh در هیپوکامپ و قشر مغز است که از مهارکنندههای AChE در بیماری آلزایمر استفاده شده است. متأسفانه، آنها فقط تا حدی مؤثر هستند. در بهترین حالت، آنها بدتر شدن علائم را برای چند ماه به تاخیر میاندازند، اما تخریب عصبی را کند نمیکنند و بهبود بالینی چشمگیری ایجاد نمیکنند (فصل 18).
The pedunculopontine nucleus and the laterodorsal tegmental nucleus within the brainstem innervate dopaminergic neurons of the VTA and play a role in reward. They also innervate other brainstem nuclei as well as thalamocortical circuits to control sleep and arousal (Chapter 13). While these nuclei, as well as the nucleus basalis, innervate the basal ganglia, the most significant cholinergic innervation of striatum comes from large cholinergic interneurons intrinsic to this region. These interneurons are critical components of the complex circuitry that underlies motor learning and motor control. As mentioned above, loss of dopaminergic input to the striatum in Parkinson disease leads to a profound motor disorder that can be partly counteracted by muscarinic cholinergic antagonists such as trihexyphenidyl and benztropine. These can be used in the early stages of the disease, or to supplement other drugs, such as L-dopa. Antimuscarinic agents also are used to treat the extrapyramidal side effects of antipsychotic drugs. Muscarinic cholinergic antagonists are believed to act at muscarinic receptors on striatal medium spiny neurons, where they partially compensate for the decreased dopaminergic signaling in striatal circuits (Chapter 18).
هسته پدانکولوپونتین و هسته لاترودورسال تگمنتال درون ساقه مغز نورونهای دوپامینرژیک VTA را عصب دهی میکنند و در پاداش نقش دارند. آنها همچنین سایر هستههای ساقه مغز و همچنین مدارهای تالاموکورتیکال را برای کنترل خواب و برانگیختگی عصبدهی میکنند (فصل 13). در حالی که این هستهها و همچنین هسته بازالیس، عقدههای قاعدهای را عصب دهی میکنند، مهم ترین عصب کولینرژیک جسم مخطط از نورونهای داخلی کولینرژیک اصلی بزرگ این ناحیه ناشی میشود. این بین نورونها اجزای حیاتی مدار پیچیده ای هستند که زمینه یادگیری حرکتی و کنترل حرکتی را تشکیل میدهند. همانطور که در بالا ذکر شد، از دست دادن ورودی دوپامینرژیک به جسم مخطط در بیماری پارکینسون منجر به یک اختلال حرکتی شدید میشود که میتواند تا حدی توسط آنتاگونیستهای کولینرژیک موسکارینی مانند تری هگزی فنیدیل و بنزتروپین خنثی شود. اینها یا مکمل سایر داروها مانند ال-دوپا را میتوان در مراحل اولیه بیماری استفاده کرد. از عوامل ضد موسکارینی نیز برای درمان عوارض جانبی خارج هرمی داروهای ضد روان پریشی استفاده میشود. اعتقاد بر این است که آنتاگونیستهای کولینرژیک موسکارینی در گیرندههای موسکارینی روی نورونهای خاردار محیط مخطط عمل میکنند، جایی که آنها تا حدی کاهش سیگنال دهی دوپامینرژیک در مدارهای جسم مخطط را جبران میکنند (فصل 18).
Storage and Release
ذخیره و آزادسازی
The cholinergic nerve-muscle synapse is the best studied synapse in neurobiology and has taught us an enormous amount about synaptic transmission as well as the mechanisms of synaptogenesis in both development and repair. Central cholinergic synapses differ in important regards, most significantly by being nerve-nerve synapses. In presynaptic terminals, ACh is concentrated in storage vesicles by a vesicular transporter (6-18) that can be inhibited by vesamicol, resulting in depletion of ACh from vesicles. Vesicles fuse with the presynaptic terminal membrane in response to depolarization, releasing ACh into the synapse. Several toxins interfere selectively with this vesicular release of ACh. Botulinum toxin A and tetanus toxin, which prevent ACh release from cholinergic motor neuron terminals, cause paralysis. Small injections of botulinum toxin (Botox) are used therapeutically to treat dystonias (by causing profound muscle relaxation) and also cosmetically to inhibit the movement of facial muscles that cause wrinkles. Black widow spider venom, which contains ɑ-latrotoxin, promotes massive vesicular release of ACh and a subsequent overstimulation of postsynaptic neurons. It exerts its effects in part by uncoupling Ca2+ signals from the release process. a-Latrotoxin stimulates exocytosis by binding to neurexins and latrophilins, critical components of nerve terminals (Chapter 3).
سیناپس عصب-عضله کولینرژیک بهترین سیناپس مورد مطالعه در نوروبیولوژی است و اطلاعات زیادی در مورد انتقال سیناپسی و همچنین مکانیسمهای سیناپتوژنز در توسعه و بازسازی به ما آموخته است. سیناپس های کولینرژیک مرکزی از جنبه های مهمی با هم تفاوت دارند که مهم ترین آنها سیناپس های عصبی-عصبی است. در پایانههای پیش سیناپسی، ACh توسط یک ترانسپورتر وزیکولی (6-18) در وزیکولهای ذخیره سازی غلیظ میشود که میتواند توسط وزامیکول مهار شود و منجر به تخلیه Ach از وزیکولها میشود. وزیکولها در پاسخ به دپلاریزاسیون با غشای انتهایی پیش سیناپسی ترکیب میشوند و ACH را در سیناپس آزاد میکنند. چندین سم به طور انتخابی از آزادسازی وزیکولی ACh جلوگیری میکنند. سم بوتولینوم A و سم کزاز که از ترشح ACh از پایانههای نورون حرکتی کولینرژیک جلوگیری میکنند، باعث فلج میشوند. تزریقهای کم سم بوتولینوم (بوتاکس) برای درمان دیستونی (با ایجاد شل شدگی شدید عضلانی) و همچنین از نظر زیبایی برای مهار حرکت عضلات صورت که باعث ایجاد چین و چروک میشوند، استفاده میشود. زهر عنکبوت بیوه سیاه، که حاوی آلفا-لاتروتوکسین است، باعث آزادسازی وزیکولی فراوان ACh و تحریک بیش از حد نورونهای پس سیناپسی میشود. اثرات خود را تا حدی بدون وابستگی به سیگنالهای کلسیم در فرآیند آزادسازی اعمال میکند. آلفا-لاتروتوکسین با اتصال به نوروکسینها و لاتروفیلینها، اجزای مهم پایانههای عصبی، اگزوسیتوز را تحریک میکند (فصل 3).
Because ACh is not returned to the presynaptic terminal by a membrane transporter, it cannot be recycled like the monoamines. Instead, choline, released by the action of AChE on ACh, is returned to cholinergic terminals by a high-affinity choline transporter, where it is used to synthesize new transmitter. Hemicholinium is a high-affinity inhibitor of the choline transporter.
از آنجایی که ACh توسط یک ترانسپورتر غشایی به پایانه پیش سیناپسی بازگردانده نمیشود، نمیتوان آن را مانند مونوآمینها بازیافت کرد. درعوض، کولین آزاد شده توسط AChE بر روی ACh، توسط یک انتقال دهنده کولین با میل ترکیبی بالا به پایانههای کولینرژیک بازگردانده میشود، جایی که برای سنتز ترنسمیتر جدید استفاده میشود. همیکولینیوم یک مهارکننده ترانسپورتر کولین است که میل ترکیبی زیادی به این ترانسپورتر دارد.
Acetylcholine Receptors
گیرندههای استیل کولین
ACh acts on two major types of receptors named for naturally occurring agonists: (1) muscarinic receptors, which are members of the GPCR superfamily, and (2) nicotinic receptors, which are members of the ligand- gated ion channel superfamily. Other members of the latter family include 5HT3 receptors, the ionotropic glutamate receptors, GABAA receptors, and glycine receptors. Structural similarities among these ligand- gated channels are represented in 6-19.
ACh روی دو نوع گیرنده اصلی عمل میکند؛ این براساس آگونیستهای طبیعی نامگذاری شدهاند: (1) گیرندههای موسکارینی، که جزء ابرخانواده GPCR هستند، و (2) گیرندههای نیکوتینی، که جزء خانواده کانالهای یونی دریچهدار لیگاندی هستند. سایر اعضای خانواده اخیر شامل گیرندههای 5HT3، گیرندههای یونوتروپیکی گلوتامات، گیرندههای گاباآ و گیرندههای گلیسین هستند. شباهتهای ساختاری بین این کانالهای دریچهدار لیگاندی در 6-19 نشان داده شده است.

6-19 Ligand-gated ion channels. Considerable structural similarity characterizes the neurotransmitter receptors in this superfamily. Each receptor subunit has an extracellular N terminus, four membrane-spanning domains (M1-M4), and a short extracellular C terminus. A single disulfide bond is conserved among all members of this family, with the exception of nicotinic α1 subunits, which possess two disulfide bonds.
6-19 کانالهای یونی دریچهدار لیگاندی. شباهت ساختاری قابل توجه گیرندههای انتقال دهنده عصبی را در این ابرخانواده مشخص میکند. هر زیر واحد گیرنده دارای یک پایانه N خارج سلولی، چهار دُمین عرض غشایی (M1-M4) و یک پایانه C خارج سلولی کوتاه است. یک پیوند دی سولفیدی در بین تمام اعضای این خانواده حفظ شده است، به استثنای زیر واحدهای نیکوتین α1 که دارای دو پیوند دی سولفیدی هستند.
Muscarinic receptors: Muscarinic receptors were named for their ability to bind muscarine, a compound derived from the poisonous mushroom Amanita muscaria. They are located peripherally in the autonomic nervous system (Chapter 9) and in the CNS. There are five subtypes, all expressed in brain, each with the canonical seven membrane-spanning domains that characterize GPCRs. Muscarinic receptors can be subgrouped based on their patterns of G protein coupling: M1, M3, and M5 receptors couple to Gq, and M2 and M4 receptors couple to G; 6-6. Electrophysiologic responses elicited by the activation of any given subtype can vary from one cell to another, depending on the second messenger systems and ion channels expressed in the cell. MJ, M3, and Ms receptors produce their effects by stimulating the phosphatidylinositol system. As with most other G; -linked receptors, M2 and M4 receptors elicit mostly inhibitory responses; they act by activating inwardly rectifying K channels, inhibiting voltage-gated Ca channels, and inhibiting adenylyl cyclase.
گیرندههای موسکارینی: گیرندههای موسکارینی به دلیل توانایی آنها در اتصال موسکارین نامگذاری شدند. موسکارین یک ترکیب مشتق شده از قارچ سمی Amanita muscaria است. آنها به صورت محیطی در سیستم عصبی خودمختار (فصل 9) و در CNS قرار دارند. پنج زیرگروه وجود دارد که همگی در مغز بیان میشوند و هر کدام دارای هفت حوزه متعارف غشایی هستند که GPCRها را مشخص میکنند. گیرندههای موسکارینی را میتوان بر اساس الگوهای جفت شدن پروتئین G زیر گروه بندی کرد: گیرندههای M1، M3 و M5 به Gq و گیرندههای M2 و M4 به G جفت میشوند. 6-6. پاسخهای الکتروفیزیولوژیک ناشی از فعالسازی هر زیرگروه میتواند از سلولی به سلول دیگر متفاوت باشد، بسته به سیستمهای پیامرسان دوم و کانالهای یونی بیانشده در سلول. گیرندههای MJ، M3 و Ms اثرات خود را با تحریک سیستم فسفاتیدیلینوزیتول ایجاد میکنند. مانند اکثر Gهای دیگر. گیرندههای مرتبط، گیرندههای M2 و M4 عمدتاً پاسخهای بازدارنده را ایجاد میکنند. آنها با فعال کردن کانالهای K اصلاحکننده درونی، مهار کانالهای کلسیم دارای ولتاژ و مهار آدنیلیل سیکلاز عمل میکنند.
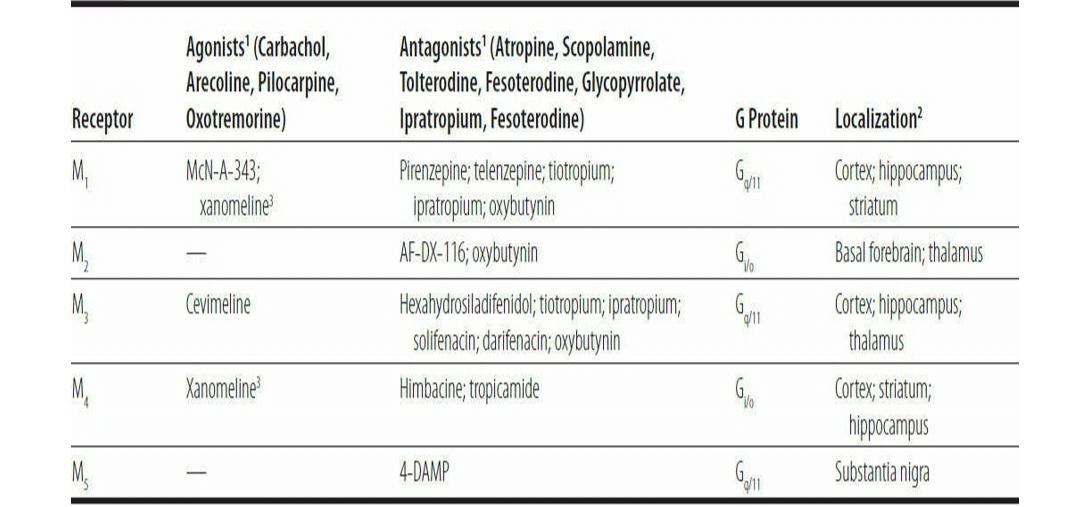
6-6 Muscarinic Cholinergic Receptors
گیرندههای کولینرژیکی موسکارینی
4-DAMP 4-diphenylacetoy-N-methylpiperidine; Mot-343,4-13-chlorophenylcarbamylony2-buty
‘An increasing number of agonists and antagonists selective for certain muscarinic receptor subtypes have become available in recent years. Examples of nonspecific or general agents are given in parentheses. See Chapter 9 for the use of many of these drugs in targeting the autonomic nervous system.
2mRNA expression for these receptors has been determined in rat brain; the distribution of each subtype may differ in human tissue.
Yannis positive lasteric modular with same specificity for M1, and M4 receptors.
M1, M3, and M4 receptors are primarily located in the cerebral cortex and hippocampus, where their activity may mediate some of the effects of ACh on learning and memory. In the striatum, M1 and M4 subtypes occur in abundance and mediate cholinergic signaling in extrapyramidal motor circuits and in responses to rewards. M2 receptors are concentrated in the basal forebrain, the site of several cholinergic nuclei, where they act as autoreceptors to control ACh synthesis and release. The M5 receptor is the least abundant of the muscarinic receptors and is expressed at low levels throughout brain.
General muscarinic agonists include several alkaloids, such as muscarine, pilocarpine, and arecoline, and many synthetic compounds, such as carbachol and oxotremorine. Centrally acting agonists produce arousal as well as peripheral effects such as excessive salivation and sweating. Arecoline is an ingredient of betel leaves, which are chewed in some Asian cultures to produce mild increases in arousal. Muscarinic cholinergic agonists are used clinically to treat glaucoma and urinary retention, and to ameliorate the dry mouth that characterizes Sjögren syndrome, a disorder of autoimmune degeneration of the salivary glands (Chapter 9). An increasing number of subtype-selective muscarinic receptor agonists and antagonists are being developed for clinical use; most target the autonomic nervous system and regulate peripheral organ function, although interest continues in the possible use of such agents to enhance cognitive function (Chapter 14).
گیرندههای M1، M3 و M4 عمدتاً در قشر مغز و هیپوکامپ قرار دارند، جایی که فعالیت آنها ممکن است برخی از اثرات ACH بر یادگیری و حافظه را واسطه کند. در جسم مخطط، زیرگروههای M1 و M4 به وفور رخ میدهند و سیگنالهای کولینرژیک را در مدارهای موتور خارج هرمیو در پاسخ به پاداشها واسطه میکنند. گیرندههای M2 در پیشمغز قاعدهای، محل چندین هسته کولینرژیک، متمرکز شدهاند، جایی که بهعنوان گیرندههای خودکار برای کنترل سنتز و انتشار ACh عمل میکنند. گیرنده M5 کمترین فراوانی در بین گیرندههای موسکارینی است و در سطوح پایین در سراسر مغز بیان میشود. آگونیستهای موسکارینی عمومیشامل چندین آلکالوئید مانند موسکارین، پیلوکارپین و آرکولین و بسیاری از ترکیبات مصنوعی مانند کارباکول و اکسوترمورین هستند. آگونیستهای با اثر مرکزی باعث برانگیختگی و همچنین اثرات محیطی مانند ترشح بیش از حد بزاق و تعریق میشوند. آرکولین یکی از اجزای برگ فوفل است که در برخی از فرهنگهای آسیایی جویده میشود تا باعث افزایش خفیف در برانگیختگی شود. آگونیستهای کولینرژیک موسکارینی از نظر بالینی برای درمان گلوکوم و احتباس ادرار، و برای بهبود خشکی دهان که مشخصه سندرم شوگرن، یک اختلال انحطاط خودایمنی غدد بزاقی است، استفاده میشوند (فصل 9). تعداد فزاینده ای از آگونیستها و آنتاگونیستهای گیرنده موسکارینی انتخابی فرعی برای استفاده بالینی در حال توسعه هستند. بیشتر آنها سیستم عصبی خودمختار را هدف قرار میدهند و عملکرد اندامهای محیطی را تنظیم میکنند، اگرچه علاقه به استفاده احتمالی از چنین عواملی برای تقویت عملکرد شناختی ادامه دارد (فصل 14).
Prototypical muscarinic antagonists include atro-pine and scopolamine. Neither of these is subtype- selective. Atropine is a derivative of belladonna, whose name (beautiful woman) reflects its historical cosmetic use as a pupillary dilator. Belladonna is produced by the deadly nightshade plant, named because ingestion of excessive amounts is dangerous. Indeed, any excess use of antimuscarinic drugs produces cognitive impairment, and at higher doses delirium (Chapter 14), along with tachycardia and other dangerous autonomic symptoms. As stated earlier, muscarinic antagonists, such as benztropine, are commonly used to treat Parkinsonian symptoms induced by first-generation antipsychotic drugs. Many psychotropic drugs, including TCAs and low-potency antipsychotic drugs (eg, chlorpromazine and thioridazine), have muscarinic antagonist properties, as do many first-generation histamine H1 antagonists (antihistamines) used to treat allergies and as over-the-counter hypnotic agents. In addition to expected autonomic effects, such as dry mouth, constipation, and urinary retention, these drugs can cause delirium. The latter is seen most commonly in geriatric patients treated with high doses of these agents or when combinations of drugs that share anticholinergic properties are used.
آنتاگونیستهای موسکارینی اولیه شامل آترو کاج و اسکوپولامین هستند. هیچکدام از اینها زیرمجموعه انتخابی نیستند. آتروپین از مشتقات بلادونا است که نام آن (زن زیبا) نشان دهنده استفاده تاریخی آرایشی آن به عنوان گشادکننده مردمک است. بلادونا توسط گیاه کشنده شب بو تولید میشود که به این دلیل نامگذاری شده است که مصرف بیش از حد آن خطرناک است. در واقع، هر گونه استفاده بیش از حد از داروهای ضد موسکارینی باعث اختلال شناختی و در دوزهای بالاتر هذیان میشود (فصل 14)، همراه با تاکی کاردی و سایر علائم خطرناک اتونوم. همانطور که قبلا گفته شد، آنتاگونیستهای موسکارینی، مانند بنزتروپین، معمولاً برای درمان علائم پارکینسون ناشی از داروهای ضد روان پریشی نسل اول استفاده میشوند. بسیاری از داروهای روانگردان، از جمله TCAها و داروهای ضد روان پریشی کم قدرت (مانند کلرپرومازین و تیوریدازین)، دارای خواص آنتاگونیست موسکارینی هستند، مانند بسیاری از آنتاگونیستهای هیستامین H1 نسل اول (آنتی هیستامینها) که برای درمان آلرژیها و داروهای هیپنوتیزم بدون نسخه استفاده میشوند. . علاوه بر اثرات اتونوم مورد انتظار مانند خشکی دهان، یبوست و احتباس ادرار، این داروها میتوانند باعث هذیان شوند. مورد دوم بیشتر در بیماران سالمندی دیده میشود که با دوزهای بالای این عوامل درمان میشوند یا زمانی که ترکیبی از داروهایی که خواص آنتی کولینرژیک مشترک دارند استفاده میشود.
Nicotinic receptors Nicotinic ACh receptors (nAChRs) are located at the neuromuscular junction, autonomic ganglia, adrenal medulla, and CNS. These ligand-gated ion channels were first characterized based on their ability to bind nicotine isolated from the tobacco plant Nicotiana tabacum. Their activation by ACh leads to the rapid influx of Na* and Cat and subsequent cellular depolarization. A prominent feature of AChRs is their very rapid desensitization.
Unlike desensitization of GPCRs, which requires the actions of other proteins (Chapter 4), desensitization of nAChRs is an intrinsic property of the receptors themselves. The same may hold true for other iono-tropic receptors. The latter type of desensitization may be similar to inactivation of voltage-gated Na+ channels (Chapter 2).
گیرندههای نیکوتینی گیرندههای نیکوتینی ACh (nAChRs) در محل اتصال عصبی عضلانی، گانگلیونهای اتونوم، مدولای آدرنال و CNS قرار دارند. این کانالهای یونی دارای لیگاند ابتدا بر اساس توانایی آنها در اتصال نیکوتین جدا شده از گیاه تنباکو Nicotiana tabacum مشخص شدند. فعال شدن آنها توسط ACh منجر به هجوم سریع Na* و Cat و متعاقب آن دپلاریزاسیون سلولی میشود. یکی از ویژگیهای برجسته AChRها حساسیت زدایی بسیار سریع آنهاست. بر خلاف حساسیت زدایی از GPCRها، که به اعمال سایر پروتئینها نیاز دارد (فصل 4)، حساسیت زدایی از nAChRها یک ویژگی ذاتی خود گیرندهها است. همین امر ممکن است در مورد سایر گیرندههای یونی-گرمسیری نیز صادق باشد. نوع دوم حساسیت زدایی ممکن است شبیه غیرفعال کردن کانالهای Na+ دردار ولتاژ باشد (فصل 2).
nAChRs expressed in neurons and those expressed in muscle differ in their subunit composition. nAChRs are composed of five subunits organized around a central pore (6-20). Most receptors are composed of heterologous subunits, of which at least 12 have been identified to date. Eight are classified as a subunits (α2- a) and three as ẞ subunits (B2-B4); a1 and ẞ1 subunits are expressed in muscle. Like other members of the ligand-gated ion channel family, each subunit has four membrane-spanning domains and disulfide loops 20. The second transmembrane (M2) region of each subunit in each pentameric receptor lines the pore of the channel to regulate its gating properties.
nAChRهای بیان شده در نورونها و آنهایی که در عضله بیان میشوند در ترکیب زیر واحد خود متفاوت هستند. nAChRها از پنج زیر واحد تشکیل شده اند که در اطراف یک منفذ مرکزی سازماندهی شده اند (6-20). اکثر گیرندهها از زیر واحدهای هترولوگ تشکیل شده اند که حداقل 12 مورد از آنها تا به امروز شناسایی شده است. هشت مورد به عنوان یک زیر واحد (α2-a) و سه زیر واحد به عنوان زیرواحد ẞ (B2-B4) طبقه بندی میشوند. زیر واحدهای a1 و ẞ1 در عضله بیان میشوند. مانند سایر اعضای خانواده کانالهای یونی دریچه ای لیگاند، هر زیر واحد دارای چهار حوزه پوشاننده غشاء و حلقههای دی سولفیدی 20 است. دومین ناحیه گذر غشایی (M2) هر زیر واحد در هر گیرنده پنتامری، منافذ کانال را برای تنظیم ویژگیهای دروازه ای آن میپوشاند.
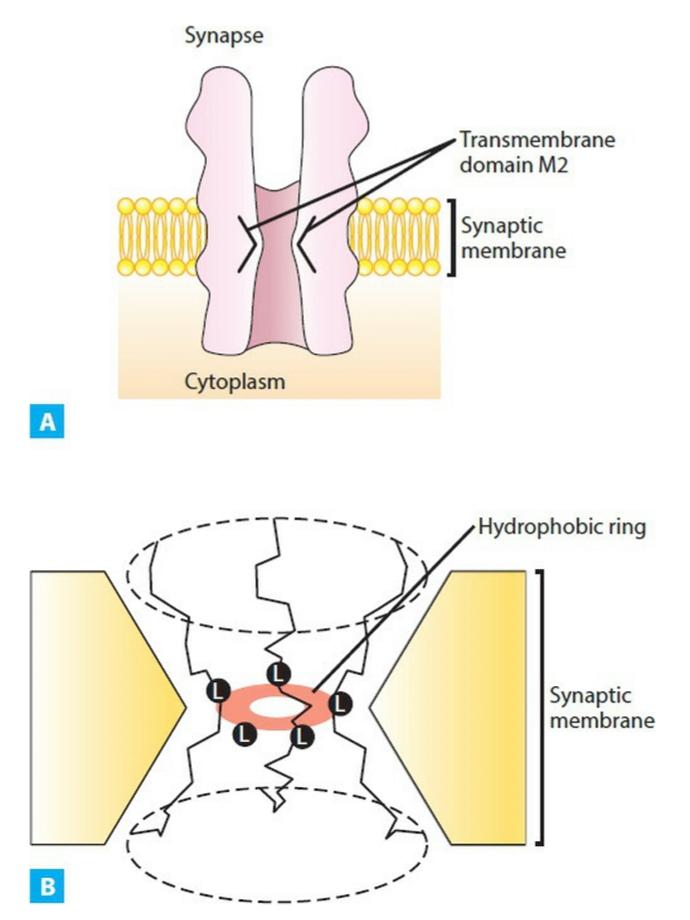
6-20 Model of the nicotinic acetylcholine receptor. Crystallized synaptic membranes were examined with an electron microscope to predict the three-dimensional structure of this receptor. A. A cylindrical membrane-embedded structure with a central pore is shown. The second transmembrane domain (M2) of each subunit lines the pore and bends inward to block ion flow through the channel. B. A highly conserved leucine residue (L) in the M2 bend of each subunit is believed to protrude into the pore to form a tight hydro- phobic ring, which may act as a barrier to the flow of hydrated ions across the channel. (Adapted with permission from Unwin N: Nicotinic acetylcholine receptor at 9 A resolution. J Mol Biol. 1993;229(4):1101- 1124)
6-20 مدل گیرنده نیکوتین استیل کولین. غشاهای سیناپسی متبلور با میکروسکوپ الکترونی برای پیش بینی ساختار سه بعدی این گیرنده مورد بررسی قرار گرفتند. الف. یک ساختار استوانه ای تعبیه شده در غشاء با منفذ مرکزی نشان داده شده است. دومین حوزه گذر غشایی (M2) هر زیر واحد، منافذ را میپوشاند و به سمت داخل خم میشود تا جریان یون را از طریق کانال مسدود کند. ب. اعتقاد بر این است که یک باقیمانده لوسین بسیار حفاظت شده (L) در خم M2 هر زیر واحد به داخل منافذ بیرون زده و یک حلقه آبگریز محکم ایجاد میکند، که ممکن است به عنوان مانعی برای جریان یونهای هیدراته در سراسر کانال عمل کند. (اقتباس شده با مجوز Unwin N: گیرنده استیل کولین نیکوتینی با وضوح 9 A. J Mol Biol. 1993;229(4):1101-1124)
Most neuronal nAChRs contain both a and ẞ sub-units, and their stoichiometry is such that the ratio of these subunits typically is two to three. However, some nAChRs are homomeric; a7 subunits, for example, are capable of forming functional channels in vitro without the presence of other subunits. Several major types of nAChR complexes have been identified based on their affinities for certain toxins; for example, one population of receptors (ie, a7 homomeric receptors) binds a-bungarotoxin with high affinity and nicotine with low affinity, whereas most other populations bind nicotine with high affinity and are insensitive to bungarotoxin.
اکثر nAChRهای عصبی حاوی هر دو زیر واحد a و ẞ هستند و استوکیومتری آنها به گونه ای است که نسبت این زیر واحدها معمولاً دو به سه است. با این حال، برخی از nAChRها همومریک هستند. به عنوان مثال، زیر واحدهای a7 قادر به تشکیل کانالهای عملکردی در شرایط آزمایشگاهی بدون حضور زیر واحدهای دیگر هستند. چندین نوع اصلی از کمپلکسهای nAChR بر اساس تمایل آنها به سموم خاص شناسایی شدهاند. به عنوان مثال، یک جمعیت از گیرندهها (یعنی گیرندههای همومریک a7) a-bungarotoxin را با میل ترکیبی بالا و نیکوتین را با میل ترکیبی کم متصل میکنند، در حالی که اکثر جمعیتهای دیگر به نیکوتین با میل ترکیبی بالا متصل میشوند و به بونگاروتوکسین حساس نیستند.
Only a small number of ligands bind at nAChRs. Curare blocks muscular and, to a lesser extent, neuronal nAChRs. Succinylcholine, a paralytic agent that is used clinically during anesthesia, acts as a weak partial agonist; it mildly activates nAChRs and subsequently induces their prolonged desensitization. Several blocking agents, including hexamethonium and mecamyla-mine, inhibit nAChRs that mediate neurotransmission in both sympathetic and parasympathetic ganglia (Chapter 9). Very few antagonists that are selective for subtypes of neuronal nAChRs have been developed. Perhaps the most selective antagonist available is methyllycaconitine, which preferentially antagonizes a7 homomeric receptor complexes. Recently, varenicline, a partial agonist at a4/ẞ2 nAChRs, has been introduced to promote smoking cessation (Chapter 16). The selective agonist epibatidine binds with high affinity at nicotinic receptors but has been used only for experimental purposes (eg, as an antinociceptive agent).
فقط تعداد کمیاز لیگاندها در nAChRs متصل میشوند. Curare nAChRهای عضلانی و تا حدی عصبی را مسدود میکند. سوکسینیل کولین، یک عامل فلج کننده که به صورت بالینی در طول بیهوشی استفاده میشود، به عنوان یک آگونیست نسبی ضعیف عمل میکند. nAChRها را به طور ملایم فعال میکند و متعاقبا باعث حساسیت زدایی طولانی مدت آنها میشود. چندین عامل مسدودکننده، از جمله هگزامتونیوم و مکامیلا مین، nAChRs را که واسطه انتقال عصبی در عقدههای سمپاتیک و پاراسمپاتیک هستند، مهار میکنند (فصل 9). تعداد بسیار کمیآنتاگونیست انتخابی برای زیرگروههای nAChRهای عصبی ایجاد شده اند. شاید انتخابی ترین آنتاگونیست موجود متیل لیکاکونیتین باشد که ترجیحاً با کمپلکسهای گیرنده همومریک a7 مخالفت میکند. اخیراً، وارنیکلین، آگونیست نسبی در a4/ẞ2 nAChRs، برای ترویج ترک سیگار معرفی شده است (فصل 16). آگونیست انتخابی اپیباتیدین با میل ترکیبی بالایی به گیرندههای نیکوتین متصل میشود، اما فقط برای اهداف تجربی (مثلاً به عنوان یک عامل ضد درد) استفاده شده است.
As their name implies, nAChRs bind nicotine, which is a highly addictive substance (Chapter 16). The normal role of nicotinic receptors, however, appears to lie in arousal, attention, and memory. The widespread use of tobacco by people with schizophrenia has raised the questions of whether nicotine is being used to self- medicate cognitive impairments. Some epidemiologic studies indicate that smoking reduces the likelihood of developing two neurodegenerative diseases, Alzheimer disease and Parkinson disease. Unfortunately, the use of nicotine or tobacco products is fraught with complications, most notably addiction and the diverse ill effects of inhaling or chewing tobacco (including cardiovascular disease, respiratory disease, and diverse cancers). Nevertheless, interest remains high in the pharmaceutical industry to develop more selective nicotinic agents for a range of medicinal purposes including the enhancement of cognition (Chapter 14).
همانطور که از نام آنها پیداست، nAChRها نیکوتین را که یک ماده بسیار اعتیادآور است، متصل میکنند (فصل 16). نقش طبیعی گیرندههای نیکوتین، با این حال، به نظر میرسد در برانگیختگی، توجه و حافظه نهفته است. استفاده گسترده از تنباکو توسط افراد مبتلا به اسکیزوفرنی این سؤال را ایجاد کرده است که آیا نیکوتین برای خوددرمانی اختلالات شناختی استفاده میشود یا خیر. برخی از مطالعات اپیدمیولوژیک نشان میدهد که سیگار احتمال ابتلا به دو بیماری تخریب کننده عصبی، بیماری آلزایمر و بیماری پارکینسون را کاهش میدهد. متأسفانه، استفاده از نیکوتین یا محصولات تنباکو مملو از عوارضی است که مهمترین آنها اعتیاد و اثرات نامطلوب مختلف استنشاق یا جویدن تنباکو (از جمله بیماریهای قلبی عروقی، بیماریهای تنفسی و سرطانهای مختلف) است. با این وجود، علاقه در صنعت داروسازی برای توسعه عوامل نیکوتین انتخابی بیشتر برای طیف وسیعی از اهداف دارویی از جمله تقویت شناخت (فصل 14) همچنان بالاست.
HISTAMINE
هیستامین
Synthetic and Degradative Pathways
مسیرهای سنتز و تخریب
Histamine is produced in one step by decarboxylation of the amino acid histidine by histidine decarboxylase (6-21). Histidine also can be decarboxylated by AADC, which, as previously discussed, is involved in the synthesis of DA, NE, and 5HT. Histamine is metabolized into methyl histidine by histamine methyltransferase. Alternatively, diamine oxidase may convert the neurotransmitter into imidazole acetaldehyde.
هیستامین در یک مرحله با دکربوکسیلاسیون اسید آمینه هیستیدین توسط هیستیدین دکربوکسیلاز تولید میشود (6-21). هیستیدین همچنین میتواند توسط AADC دکربوکسیله شود، که همانطور که قبلاً بحث شد، در سنتز DA، NE و 5HT نقش دارد. هیستامین توسط هیستامین متیل ترانسفراز به متیل هیستیدین متابولیزه میشود. از طرف دیگر، دیامین اکسیداز ممکن است انتقال دهنده عصبی را به ایمیدازول استالدئید تبدیل کند.

6-21 Biosynthetic pathway of histamine. Histamine is produced from the decarboxylation of the amino acid histidine and is subsequently metabolized into one of three products. Diamine oxidase converts histamine into imidazole acetaldehyde, and histamine methyltransferase converts the neurotransmitter into its other major metabolite, methylhistamine. Acetyl histamine is a minor metabolite.
6-21 مسیر بیوسنتزی هیستامین. هیستامین از دکربوکسیلاسیون اسید آمینه هیستیدین تولید میشود و متعاقباً به یکی از سه محصول متابولیزه میشود. دیامین اکسیداز هیستامین را به ایمیدازول استالدئید تبدیل میکند و هیستامین متیل ترانسفراز انتقال دهنده عصبی را به دیگر متابولیت اصلی خود یعنی متیل هیستامین تبدیل میکند. استیل هیستامین یک متابولیت جزئی است.
Functional Anatomy
آناتومیکاربردی
Within the brain, histamine is synthesized exclusively by neurons with their cell bodies in the tuber- omammillary nucleus (TMN) that lies within the posterior hypothalamus (PH) (6-22). There are approximately 64,000 histaminergic neurons per side in humans. These cells project throughout the brain and spinal cord. Areas that receive especially dense projections include the cerebral cortex, hippocampus, neostriatum, nucleus accumbens, amygdala, and hypothalamus.
در داخل مغز، هیستامین منحصراً توسط نورونها با بدن سلولی آنها در هسته غدهامیلاری (TMN) که در هیپوتالاموس خلفی (PH) قرار دارد، سنتز میشود (6-22). تقریباً 64000 نورون هیستامینرژیک در هر طرف در انسان وجود دارد. این سلولها در سراسر مغز و نخاع پخش میشوند. نواحی که برجستگیهای بسیار متراکمیدریافت میکنند شامل قشر مغز، هیپوکامپ، نئوستریتوم، هسته اکومبنس، آمیگدال و هیپوتالاموس است.

6-22 Histaminergic projections in the CNS. Histaminergic neurons are located solely in the tuberomammillary nucleus of the hypothalamus and give rise to widespread projections.
6-22 برآمدگی هیستامینرژیک در CNS. نورونهای هیستامینرژیک تنها در هسته توبروممیلاری هیپوتالاموس قرار دارند و باعث برآمدگیهای گسترده میشوند.
The TMN is innervated by other widely projecting systems including monoaminergic and orexinergic neurons. Histaminergic neurons project back, in turn, to monoamine and cholinergic neurons involved in arousal, attention, learning, and memory 6-23. While the best characterized function of the histamine system in the brain is regulation of sleep and arousal, histamine is also involved in learning and memory through its direct actions on the cerebral cortex, hippo-campus, and amygdala, and indirectly by its effects on cholinergic and monoamine neurons. It also appears that histamine is involved in the regulation of feeding and energy balance. The firing rate of histaminergic neurons in the TMN is strongly correlated with states of arousal. Firing is fastest during periods of wakefulness and slower during sleep; indeed, these cells fall silent during slow-wave sleep (Chapter 13).
TMN توسط سایر سیستمهای به طور گسترده ای از جمله نورونهای مونوآمینرژیک و اورکسینرژیک عصب دهی میشود. نورونهای هیستامینرژیک به نوبه خود به نورونهای مونوآمین و کولینرژیک درگیر در برانگیختگی، توجه، یادگیری و حافظه 6-23 برمیگردند. در حالی که بهترین عملکرد مشخص شده سیستم هیستامین در مغز تنظیم خواب و برانگیختگی است، هیستامین همچنین از طریق اعمال مستقیم خود بر روی قشر مغز، پردیس هیپو و آمیگدال، و به طور غیرمستقیم با اثرات آن بر کولینرژیک، در یادگیری و حافظه نیز نقش دارد. و نورونهای مونوآمین همچنین به نظر میرسد که هیستامین در تنظیم تغذیه و تعادل انرژی نقش دارد. سرعت شلیک نورونهای هیستامینرژیک در TMN به شدت با حالتهای برانگیختگی در ارتباط است. شلیک در زمان بیداری سریعتر و در هنگام خواب کندتر است. در واقع، این سلولها در طول خواب موج آهسته ساکت میشوند (فصل 13).

6-23 Afferent and efferent connections of the tuberomammillary nucleus. The diagram shows possible connections of histamine neurons involved in sleep and arousal, feeding, and cognition. All histamine in the brain and spinal cord is produced by the tuberomammillary nucleus (TMN) of the hypothalamus (see 6-22). The TMN is under inhibitory control of GABAergic neurons from the ventrolateral peroptic area (VLPO). There is evidence that endocannabinoids activate TMN neurons. Histaminergic neurons influence learning and memory both by direct projections to the cerebral cortex and hippocampus and by stimulating the cholinergic basal forebrain, which innervates these structures. Histaminergic neurons also project to regions of the brain involved in feeding. BLA, basolateral amygdala; NBM, nucleus basalis of Meynert. (Reproduced with permission from Passani MB, Giannoni P, Bucherelli C, Baldi E, Blandina P. Histamine in the brain: beyond sleep and memory. Biochem Pharmacol. 2007;73(8):1113-1122.)
6-23 اتصالات آوران و وابران هسته توبرومامیلاری. این نمودار اتصالات احتمالی نورونهای هیستامینی را نشان میدهد که در خواب و برانگیختگی، تغذیه و شناخت نقش دارند. تمام هیستامین در مغز و نخاع توسط هسته توبروممیلاری (TMN) هیپوتالاموس تولید میشود (6-22 را ببینید). TMN تحت کنترل مهاری نورونهای GABAergic از ناحیه پراپتیک بطنی جانبی (VLPO) است. شواهدی وجود دارد که اندوکانابینوئیدها نورونهای TMN را فعال میکنند. نورونهای هیستامینرژیک بر یادگیری و حافظه تأثیر میگذارند، هم از طریق برآمدگی مستقیم به قشر مغز و هیپوکامپ و هم با تحریک پیش مغز پایه کولینرژیک، که این ساختارها را عصب میکند. نورونهای هیستامینرژیک نیز به مناطقی از مغز که درگیر تغذیه هستند، نفوذ میکنند. BLA، آمیگدال قاعده جانبی. NBM، هسته بازالیس Meynert. (تکثیر شده با مجوز از Passani MB، Giannoni P، Bucherelli C، Baldi E، Blandina P. هیستامین در مغز: فراتر از خواب و حافظه. Biochem Pharmacol. 2007؛ 73(8):1113-1122.)
In the periphery, histamine is released from mast cells, where it plays a role in inflammatory, allergic, pruritic, and algesic responses. Histamine released in the stomach activates H2 receptors to cause release of acid.
در محیط اطراف، هیستامین از ماست سلها آزاد میشود، جایی که در پاسخهای التهابی، آلرژیک، خارش و آلژزیک نقش دارد. هیستامین آزاد شده در معده گیرندههای H2 را فعال میکند تا باعث آزاد شدن اسید شود.
Storage, Release, and Reuptake
ذخیره، آزادسازی و بازجذب
The mechanisms underlying the storage, release, and reuptake of histamine remain poorly defined (6-24). It is presumed that histamine is concentrated in synaptic vesicles, yet the vesicular transporter responsible for such localization has not been determined; it is speculated that VMAT2 may mediate this function. When histamine is released from nerve terminals in response to electrical stimulation, it acts on presynaptic and postsynaptic histamine receptors. Histamine is metabolized by the cytoplasmic enzyme, histamine N- methyltransferase. It is not clear how released histamine might be transported into neurons if this is, indeed, the major mechanism of inactivation. The OCT-2 and -3 can transport histamine when expressed in cell culture. Whether it functions as a significant histamine transporter in the brain is not yet established.
مکانیسمهای زیربنایی ذخیره، آزادسازی و بازجذب هیستامین به خوبی تعریف نشده است (6-24). فرض بر این است که هیستامین در وزیکولهای سیناپسی متمرکز است، اما ناقل تاولی که مسئول چنین محلی سازی است مشخص نشده است. حدس زده میشود که VMAT2 ممکن است واسطه این عملکرد باشد. هنگامیکه هیستامین در پاسخ به تحریک الکتریکی از پایانههای عصبی آزاد میشود، روی گیرندههای هیستامین پیش سیناپسی و پس سیناپسی اثر میگذارد. هیستامین توسط آنزیم سیتوپلاسمی، هیستامین N- متیل ترانسفراز متابولیزه میشود. مشخص نیست که چگونه هیستامین آزاد شده ممکن است به سلولهای عصبی منتقل شود، اگر این مکانیسم اصلی غیرفعال سازی باشد. OCT-2 و -3 میتوانند هیستامین را در کشت سلولی منتقل کنند. اینکه آیا به عنوان یک ناقل هیستامین قابل توجه در مغز عمل میکند، هنوز مشخص نشده است.

6-24 Model of a histaminergic synapse. Presynaptic and postsynaptic molecular entities involved in the synthesis, release, signaling, and reuptake of histamine are shown. Histamine can be metabolized by MAO (shown intracellularly) as well as by histamine methyltransferase and diamine oxidase (not shown). The transporter responsible for the reuptake of histamine has not yet been identified with certainty. See (6-6) for abbreviations.
6-24 مدل سیناپس هیستامینرژیک. موجودیتهای مولکولی پیش سیناپسی و پس سیناپسی درگیر در سنتز، آزادسازی، سیگنال دهی و بازجذب هیستامین نشان داده شده اند. هیستامین میتواند توسط MAO (در داخل سلولی نشان داده شده است) و همچنین توسط هیستامین متیل ترانسفراز و دیامین اکسیداز (نشان داده نشده) متابولیزه شود. ناقل مسئول بازجذب هیستامین هنوز با قطعیت شناسایی نشده است. برای اختصارات (6-6) را ببینید.
Histamine Receptors
گیرندههای هیستامین
Four histamine receptors (H1-H4) have been identified; all are members of the GPCR superfamily (6-7) . H1 receptors couple to Gq, and H2 receptors couple to G. The H3 receptor is thought to couple to G; and to act both as an inhibitory autoreceptor and as a hetero-receptor that regulates the release of other neurotransmitters from nerve terminals. H3 receptors have their highest distribution in the striatum, nucleus accumbens, and cerebral cortex. Antagonists and inverse agonists are being explored as potential therapeutic agents to increase alertness and enhance cognition, although clinical trials have thus far been discouraging. H4 receptors appear to play a role in inflammation in peripheral tissues. They are expressed at low levels in the brain.
چهار گیرنده هیستامین (H1-H4) شناسایی شده است. همه اعضای ابرخانواده GPCR هستند (6-7). گیرندههای H1 با Gq و گیرندههای H2 با G جفت میشوند. تصور میشود گیرنده H3 با G جفت میشود. و هم به عنوان یک گیرنده بازدارنده و هم به عنوان یک گیرنده هترو که آزاد شدن سایر انتقال دهندههای عصبی را از پایانههای عصبی تنظیم میکند. گیرندههای H3 بیشترین توزیع خود را در جسم مخطط، هسته اکومبنس و قشر مغز دارند. آنتاگونیستها و آگونیستهای معکوس به عنوان عوامل درمانی بالقوه برای افزایش هوشیاری و تقویت شناخت مورد بررسی قرار میگیرند، اگرچه آزمایشات بالینی تاکنون دلسرد کننده بوده است. به نظر میرسد گیرندههای H4 در التهاب بافتهای محیطی نقش دارند. آنها در سطوح پایین در مغز بیان میشوند.
6-7 The Histamine Receptor Family
خانواده گیرنده هیستامینی

1Selective agonists or antagonists.
آنتاگونیست یا آگونیست انتخابی
Many drugs block H1 receptors, either as an unwanted side effect (eg, the TCA doxepin and the antipsychotic drug clozapine) or as a desired mechanism as in the case of many allergy medications such as diphenhydramine. CNS-penetrant H1 receptor antagonists are sedating. Newer antihistamines used as allergy medications, such as loratadine and terfe-nadine, are nonsedating because they do not cross the blood-brain barrier. Centrally acting antihistamines are the active ingredient in many over-the-counter sleep aids. Most agents that are selective for the histamine H2 receptor do not cross the blood-brain barrier. These agents such as cimetidine and ranitidine inhibit the secretion of gastric acid.
بسیاری از داروها گیرندههای H1 را مسدود میکنند، یا به عنوان یک عارضه جانبی ناخواسته (به عنوان مثال، دوکسپین TCA و داروی ضد روان پریشی کلوزاپین) یا به عنوان مکانیسم مورد نظر مانند بسیاری از داروهای آلرژی مانند دیفن هیدرامین. آنتاگونیستهای گیرنده H1 نافذ CNS آرام بخش هستند. آنتی هیستامینهای جدیدتر که به عنوان داروهای آلرژی استفاده میشوند، مانند لوراتادین و ترفنادین، آرام بخش نیستند زیرا از سد خونی مغزی عبور نمیکنند. آنتی هیستامینهای با اثر مرکزی، ماده فعال بسیاری از داروهای خواب بدون نسخه هستند. اکثر عواملی که برای گیرنده هیستامین H2 انتخابی هستند از سد خونی مغزی عبور نمیکنند. این عوامل مانند سایمتیدین و رانیتیدین ترشح اسید معده را مهار میکنند.
OREXINS (HYPOCRETINS)
اورکسین (هیپوکرتین)
Unlike the other neurotransmitters released by widely projecting systems in the brain, the orexins (also known as hypocretins) are peptides. Orexin A and B (or hypocretin 1 and 2) are produced in the lateral hypothalamic area (LHA) and PH (6-25) and bind to two receptors that, like nearly all neuropep-tide receptors, are G protein-linked: orexin receptors 1 and 2 (OX1 and OX2). OX1 is coupled via Gq, whereas OX2 is coupled via Gio and in some cases by Gq. Like all neuropeptides (Chapter 7), the orexins are synthesized by transcription and translation and packaged in dense core vesicles for release. Differences in vesicles and release mechanisms between small molecule and peptide neurotransmitters are discussed in Chapter 7.
بر خلاف سایر انتقال دهندههای عصبی آزاد شده توسط سیستمهای به طور گسترده در مغز، اورکسینها (همچنین به عنوان هیپوکرتین شناخته میشوند) پپتید هستند. اورکسین A و B (یا هیپوکرتین 1 و 2) در ناحیه هیپوتالاموس جانبی (LHA) و PH (25-6) تولید میشوند و به دو گیرنده متصل میشوند که مانند تقریباً همه گیرندههای نوروپپ تاید، به پروتئین G مرتبط هستند: orexin. گیرندههای 1 و 2 (OX1 و OX2). OX1 از طریق Gq جفت میشود، در حالی که OX2 از طریق Gio و در برخی موارد توسط Gq جفت میشود. مانند همه نوروپپتیدها (فصل 7)، اورکسینها با رونویسی و ترجمه سنتز میشوند و در وزیکولهای هسته متراکم برای آزادسازی بسته بندی میشوند. تفاوت در وزیکولها و مکانیسمهای آزادسازی بین انتقال دهندههای عصبی مولکول کوچک و پپتیدی در فصل 7 مورد بحث قرار گرفته است.
Orexin neurons project to and activate monoaminergic and cholinergic neurons involved in the maintenance of a long “awake” period. Lack of orexin, possibly caused by degeneration of orexinergic neurons, produces narcolepsy (Chapter 13). Orexin neurons are regulated by peripheral mediators that carry information about energy balance, including glucose, leptin, and ghrelin. They also receive inputs from limbic structures. Orexin neurons are, therefore, in a position not only to regulate sleep-wake cycles but also to respond to significant environmental and metabolic signals. Accordingly, orexin plays a role in the regulation of energy homeostasis, reward, and perhaps more generally in emotion. There is a great deal of interest in generating orexin receptor antagonists, both OX1/2 dual antagonists (eg, suvorexant) and selective OX2 antagonists for the treatment of insomnia. Conversely, the possibility that orexin agonists might promote wakefulness awaits testing in clinical trials.
نورونهای اورکسین به نورونهای مونوآمینرژیک و کولینرژیک درگیر در حفظ یک دوره طولانی “بیداری” میپردازند و آنها را فعال میکنند. فقدان اورکسین، که احتمالاً به دلیل انحطاط نورونهای اورکسینرژیک ایجاد میشود، باعث ایجاد نارکولپسی میشود (فصل 13). نورونهای اورکسین توسط واسطههای محیطی که اطلاعات مربوط به تعادل انرژی از جمله گلوکز، لپتین و گرلین را حمل میکنند، تنظیم میشوند. آنها همچنین ورودیها را از ساختارهای لیمبیک دریافت میکنند. بنابراین، نورونهای اورکسین در موقعیتی هستند که نه تنها چرخههای خواب و بیداری را تنظیم میکنند، بلکه به سیگنالهای محیطی و متابولیکی قابل توجهی پاسخ میدهند. بر این اساس، اورکسین در تنظیم هموستاز انرژی، پاداش و شاید به طور کلی در احساسات نقش دارد. علاقه زیادی به تولید آنتاگونیستهای گیرنده اورکسین، هر دو آنتاگونیست دوگانه OX1/2 (مانند سوورکسانت) و آنتاگونیستهای انتخابی OX2 برای درمان بی خوابی وجود دارد. برعکس، این احتمال که آگونیستهای اورکسین ممکن است بیداری را تقویت کنند در انتظار آزمایش در آزمایشات بالینی است.
کلیک کنید «SELECTED READING»
Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci. 2005; 28:403-450.
Beaudoin-Gobert M, Sgambato-Faure V. Serotonergic pharmacology in animal models: from behavioral disorders to dyskinesia. Neuropharmacology. 2014; 81C: 15-30.
Blakely RD, Edwards RH. Vesicular and plasma membrane transporters for neurotransmitters. Cold Spring Harb Perspect Biol. 2012; 4: a005595.
Dani JA. Overview of nicotinic receptors and their roles in the central nervous system. Biol Psychiatry. 2001; 49:166-174.
Dayan P, Huys QJM. Serotonin in affective control. Annu Rev Neuurosci. 2009; 32:95–126.
Digby GJ, Shirey JK, Conn PJ. Allosteric activators of muscarinic receptors as novel approaches for treatment of CNS disorders. Mol Biosyst. 2010; 6:1345–1354.
Drenan RM, Lester HA. Insights into the neurobiology of the nicotinic cholinergic system and nicotine addiction from mice expressing nicotinic receptors harboring gain-of-function mutations. Pharmacol Rev. 2012; 64:869-879.
Fields HL, Hjelmstad GO, Margolis EB, et al. Ventral tegmental area neurons in learned appetitive behavior and positive reinforcement. Annu Rev Neurosci. 2007; 30:289–316.
Gonzalez-Maeso J, Weisstaub, NV, Zhou MM, et al. Hallucinogens recruit specific cortical 5-HT2A receptor-mediated signaling pathways to affect behavior. Neuron. 2007; 53:439–452.
Haas H, Panula P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat Rev Neurosci. 2003; 4:121–130.
Jones CK, Byun N, Bubser M. Muscarinic and nicotinic acetylcholine receptor agonists and allosteric modulators for the treatment of schizophrenia. Neuropsychopharmacology. 2012; 37:16-42.
Lefkowitz RJ. Arrestins come of age: a personal historical perspective. Prog Mol Biol Transl Sci. 2013; 118:3-18.
Mieda M, Sakurai T. Overview of orexin/hypocretin system. Prog Brain Res. 2012; 198:5–14. Mieda M, Sakurai T. Orexin (hypocretin) receptor agonists and antagonists for treatment of sleep disorders. Rationale for development and current status. CNS Drugs. 2013; 27:83-90.
Missale C, Nash SR, Robinson SW, et al. Dopamine receptors: from structure to function. Physiol Rev. 1998; 78:189-225.
Passani MB, Giannoni P, Bucherelli C, et al. Histamine in the brain: beyond sleep and memory. Biochem
Pharmacol. 2007; 73:1113-1122.
Ramsay RR. Inhibitor design for monoamine oxidases. Curr Pharm Des. 2013; 19:2529-2539.
Rasmussen SGF, DeVree BT, Zou YZ, et al. Crystal structure of the b2 adrenergic receptor-Gs protein complex. Nature. 2011; 477:549-555.
Reiter E, Ahn S, Shukla AK, Lefkowitz RJ. Molecular mechanism of ẞ-arrestin-biased agonism at seven- transmembrane receptors. Annu Rev Pharmacol Toxicol. 2012; 52:179-197.
Roeper J. Dissecting the diversity of midbrain dopa-mine neurons. Trends Neurosci. 2013; 36:336-342. Schmitt KC, Rothman RB, Reith ME. Nonclassical pharmacology of the dopamine transporter: atypical inhibitors, allosteric modulators, and partial substrates. J Pharmacol Exp Ther. 2013; 346:2-10.
Schultz W. Updating dopamine reward signals. Curr Opin Neurobiol. 2013; 23:229–238.
Seifert R, Strasser A, Schneider EH, Neumann D, Dove S, Buschauer A. Molecular and cellular analysis of human histamine receptor subtypes. Trends Pharmacol Sci. 2013; 34:33–58.
Thompson AJ. Recent developments in 5-HT3 receptor pharmacology. Trends Pharmacol Sci. 2013; 34:100-
109.
Wacker D, Wang C, Katritch V, et al. Structural features for functional selectivity at serotonin receptors. Science. 2013; 340:615-619.
Wess J. Muscarinic acetylcholine receptor knockout mice: novel phenotypes and clinical implications. Annu Rev Pharmacol Toxicol. 2004; 44:423-450.
Williams DK, Wang J, Papke RL. Positive allosteric modulators as an approach to nicotinic acetylcho-line receptor-targeted therapeutics: advantages and limitations. Biochem Pharmacol. 2011; 82:915–930.
CV استاد گرانقدر آقای دکتر علی شهبازی

 ورود/ ثبت نام با جیمیل
ورود/ ثبت نام با جیمیل
