
ترجمه و بازنویسی علمی فصول کتاب «نوروآناتومی بالینی» | کتاب درسی جامع ساختارهای عصبی (ویرایش دوم)
کتاب درسی نوروآناتومی بالینی، ویرایش دوم. Textbook of Clinical Neuroanatomy (2nd ed.) یکی از منابع معتبر در حوزه نوروآناتومی بالینی (Clinical Neuroanatomy) است که با رویکردی کاربردی، ساختارها و مسیرهای سیستم عصبی (Nervous System) را بهصورت نظاممند و بالینی شرح میدهد.
این ترجمه آموزشی، حاصل تلاش تیم علمی آیندهنگاران مغز به سرپرستی داریوش طاهری است و با هدف ارتقای آموزش علوم اعصاب (Neuroscience) در زبان فارسی، با حفظ دقت علمی، به زبانی روان تدوین شده است.
در ۲۰ فصل این مجموعه، ساختارهایی چون مغز (Brain)، نخاع (Spinal Cord)، اعصاب مغزی (Cranial Nerves)، سیستم لیمبیک (Limbic System)، مسیرهای حسی و حرکتی (Sensory and Motor Pathways)، مخچه (Cerebellum)، دیانسفالون (Diencephalon) و سیستم عصبی خودمختار (Autonomic Nervous System) بررسی میشوند.
این مجموعه برای دانشجویان پزشکی، رزیدنتها و متخصصان علاقهمند به نوروساینس بالینی طراحی شده و تلفیقی از مفاهیم بنیادی، نکات بالینی و نمودارهای عملکردی را ارائه میدهد.
با سپاس از همراهی شما
تیم آیندهنگاران مغز | سرپرستی: داریوش طاهری
18. Special Senses and their Neural Pathways
18. حواس ویژه و مسیرهای عصبی آنها
There are five types of special senses, viz. (a) sense of smell, (b) sense of vision, (c) sense of sound/ hearing, (d) sense of balance, and (e) sense of taste. The special senses have highly specialized receptors which provide specific information about the environment, i.e. they respond to only one type of stimulus. These receptors are located close to the brain and are well protected within the skull. Their responses are more complex but well coordinated within the brain. The special senses: (a) of smell and taste depend on chemoreceptors, (b) of vision on photoreceptors, and (c) of sound and balance on mechanoceptors.
پنج نوع حس ویژه وجود دارد، یعنی (الف) حس بویایی، (ب) حس بینایی، (ج) حس شنوایی/شنوایی، (د) حس تعادل و (ه) حس چشایی. حواس ویژه گیرندههای بسیار تخصصی دارند که اطلاعات خاصی در مورد محیط ارائه میدهند، یعنی فقط به یک نوع محرک پاسخ میدهند. این گیرندهها در نزدیکی مغز قرار دارند و به خوبی در جمجمه محافظت میشوند. پاسخهای آنها پیچیدهتر اما به خوبی در مغز هماهنگ است. حواس ویژه: (الف) بویایی و چشایی به گیرندههای شیمیایی، (ب) بینایی به گیرندههای نوری و (ج) صدا و تعادل به گیرندههای مکانیکی وابسته هستند.
Olfactory System
In lower vertebrates and many mammals such as dog the sense of smell is highly developed and they are called macrosmatic. In humans the sense of smell is less developed, hence they are called microsmatic.
سیستم بویایی
در مهرهداران پست و بسیاری از پستانداران مانند سگ، حس بویایی بسیار توسعهیافته است و به آنها ماکروسماتیک میگویند. در انسان حس بویایی کمتر توسعهیافته است، از این رو به آنها میکروسماتیک میگویند.
The olfactory system transmits sense of smell from olfactory epithelium of nasal mucosa to the olfactory cortex of the brain. The following structures are included in the olfactory system:
• Olfactory epithelium and olfactory nerves
• Olfactory bulb, tract and striae
• Olfactory cortical areas.
سیستم بویایی، حس بویایی را از اپیتلیوم بویایی مخاط بینی به قشر بویایی مغز منتقل میکند. ساختارهای زیر در سیستم بویایی گنجانده شدهاند:
• اپیتلیوم بویایی و اعصاب بویایی
• پیاز بویایی، مسیر بویایی و استریا
• نواحی قشر بویایی.
Olfactory Epithelium and Olfactory Nerves
The olfactory epithelium is a specialised nasal epithelium which lines the superior one-third of the nasal cavity including the roof (Fig. 18.1). It consists of three types of cells: (a) olfactory receptor cells, (b) supporting cells, and (c) progenitor/basal cells (Fig. 18.2).
اپیتلیوم بویایی و اعصاب بویایی
اپیتلیوم بویایی یک اپیتلیوم بینی تخصصی است که یک سوم فوقانی حفره بینی از جمله سقف آن را میپوشاند (شکل 18.1). این اپیتلیوم از سه نوع سلول تشکیل شده است: (الف) سلولهای گیرنده بویایی، (ب) سلولهای پشتیبان، و (ج) سلولهای پیشساز/پایه (شکل 18.2).

FIG. 18.1 Location of olfactory epithelium in the nasal cavity.
شکل ۱۸.۱ محل قرارگیری اپیتلیوم بویایی در حفره بینی.
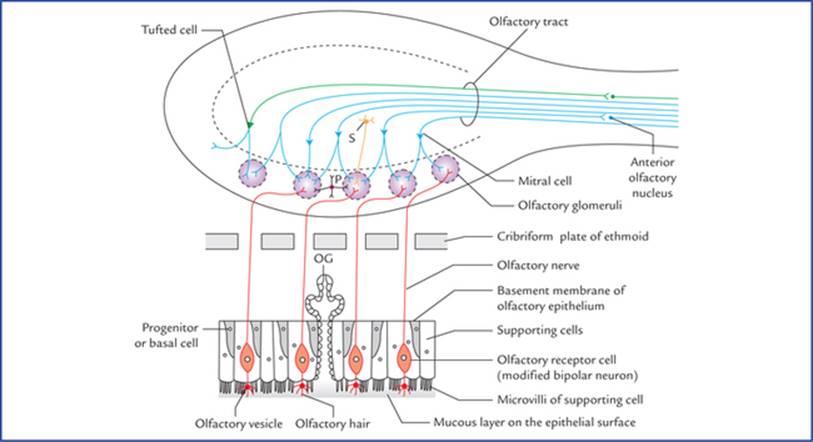
FIG. 18.2 Cellular components of olfactory epithelium and the olfactory bulb. (S = stellate cell, P = periglomerular cell, OG = olfactory glands (of Bowman).)
شکل ۱۸.۲ اجزای سلولی اپیتلیوم بویایی و پیاز بویایی. (S = سلول ستارهای، P = سلول پریگلومرولار، OG = غدد بویایی (بومن).)
The olfactory receptor cells are, in fact, the modified bipolar neurons and lie vertically between the supporting cells. Their dendrites extend as naked processes towards the free surface of the epithelium, where they end by forming bulbous enlargements called olfactory vesicles. These vesicles possess cilia—olfactory hairs which lie in a thin mucous film covering the epithelial surface. The supporting cells present many microvilli towards the free surface of the epithelium. The mucous film covering the olfactory epithelium is secreted by the olfactory glands (of Bowman) and the microvilli of the supporting cells.
سلولهای گیرنده بویایی در واقع نورونهای دوقطبی اصلاحشده هستند و به صورت عمودی بین سلولهای پشتیبان قرار دارند. دندریتهای آنها به صورت زوائد برهنه به سمت سطح آزاد اپیتلیوم امتداد مییابند، جایی که با تشکیل برآمدگیهای پیازی شکل به نام وزیکولهای بویایی به پایان میرسند. این وزیکولها دارای مژک هستند – موهای بویایی که در یک لایه مخاطی نازک قرار دارند که سطح اپیتلیوم را میپوشاند. سلولهای پشتیبان دارای میکروویلیهای زیادی به سمت سطح آزاد اپیتلیوم هستند. لایه مخاطی پوشاننده اپیتلیوم بویایی توسط غدد بویایی (بومن) و میکروویلیهای سلولهای پشتیبان ترشح میشود.
The air-borne molecules enter the nasal cavity and are dissolved in this mucous film. The odour producing molecules thus released, bound to the receptor cells. The olfactory cilia react by depolarizing and initiating action potentials in the receptor cells.
مولکولهای هوا وارد حفره بینی میشوند و در این لایه مخاطی حل میشوند. مولکولهای تولیدکننده بو که بدین ترتیب آزاد میشوند، به سلولهای گیرنده متصل میشوند. مژکهای بویایی با دپلاریزه کردن و ایجاد پتانسیلهای عمل در سلولهای گیرنده واکنش نشان میدهند.
As mentioned above, the olfactory receptors of olfactory epithelium are actually neurons. This is the only site in the body where neurons are exposed to the surface of the body.
همانطور که در بالا ذکر شد، گیرندههای بویایی اپیتلیوم بویایی در واقع نورون هستند. این تنها مکانی در بدن است که نورونها در معرض سطح بدن قرار دارند.
The receptor cells continuously degenerate and are renewed by the progenitor cells. The ability to regenerate primary sensory neurons is unique to mammalian nervous system.
سلولهای گیرنده به طور مداوم تخریب میشوند و توسط سلولهای پیشساز تجدید میشوند. توانایی بازسازی نورونهای حسی اولیه منحصر به سیستم عصبی پستانداران است.
The axons of receptor cells are fine unmyelinated fibres. They ascend and collect to form about 20 bundles called olfactory nerves. The olfactory nerves pass through the foramina of cribriform plate of ethmoid bone to enter the anterior cranial fossa where they terminate in the olfactory bulb.
آکسونهای سلولهای گیرنده، فیبرهای نازک بدون میلین هستند. آنها بالا میروند و جمع میشوند تا حدود 20 دسته به نام اعصاب بویایی تشکیل دهند. اعصاب بویایی از طریق سوراخهای صفحه مشبک استخوان اتموئید عبور میکنند تا وارد حفره جمجمهای قدامی شوند و در آنجا در پیاز بویایی خاتمه یابند.
Olfactory Bulb, Tract and Striae (Figs 18.3–18.5)
The olfactory bulb is a flattened oval mass of grey matter lying just above the cribriform plate. It consists of an outer cortical zone and an inner medullary zone.
پیاز بویایی، مسیر بویایی و خطوط بویایی (شکلهای ۱۸.۳ تا ۱۸.۵)
پیاز بویایی یک توده بیضی شکل مسطح از ماده خاکستری است که درست بالای صفحه مشبک قرار دارد. این ناحیه از یک ناحیه قشری بیرونی و یک ناحیه مغزی داخلی تشکیل شده است.
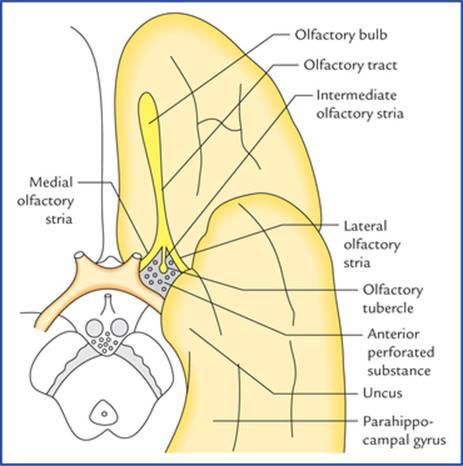
FIG. 18.3 Some of the olfactory structures related to the anterior part of the base of the brain.
شکل ۱۸.۳ برخی از ساختارهای بویایی مربوط به قسمت قدامی قاعده مغز.

FIG. 18.4 Structures on the inferior aspect of the brain in the area surrounding the optic nerves, chiasma, optic tracts, and interpeduncular fossa. Many of these structures are related to olfactory and limbic system.
شکل ۱۸.۴ ساختارهایی در سطح تحتانی مغز در ناحیه اطراف اعصاب بینایی، کیاسما، راههای بینایی و حفره بین پایهای. بسیاری از این ساختارها مربوط به سیستم بویایی و لیمبیک هستند.

FIG. 18.5 Olfactory pathway.
شکل ۱۸.۵ مسیر بویایی.
The cortical zone contains olfactory glomeruli and nerve cells which form most of the prominent cellular component of the olfactory bulb. The nerve cells of the olfactory bulb comprise (Fig. 18.2):
• Mitral cells.
• Tufted cells (analogous to but smaller than the mitral cells).
• Stellate (granule) cells, and
• Periglomerular cells.
ناحیه قشری شامل گلومرولهای بویایی و سلولهای عصبی است که بیشتر اجزای سلولی برجسته پیاز بویایی را تشکیل میدهند. سلولهای عصبی پیاز بویایی شامل موارد زیر هستند (شکل ۱۸.۲):
• سلولهای میترال.
• سلولهای تافتدار (مشابه سلولهای میترال اما کوچکتر از آنها).
• سلولهای ستارهای (گرانول) و
• سلولهای پریگلومرولار.
The medullary zone consists of nerve fibres of the olfactory tract. A small group of nerve cells situated at the transitional zone between the olfactory bulb and olfactory tract constitute the anterior olfactory nucleus.
ناحیه بصل النخاع از رشتههای عصبی مجرای بویایی تشکیل شده است. گروه کوچکی از سلولهای عصبی که در ناحیه انتقالی بین پیاز بویایی و مجرای بویایی قرار دارند، هسته بویایی قدامی را تشکیل میدهند.
Olfactory bulb continues posteriorly as olfactory tract (Figs. 18.3 and 18.4). When traced posteriorly the olfactory tract divides into medial and lateral olfactory striae. The point of bifurcation is expanded and forms the olfactory trigone. The medial and lateral olfactory striae are intimately related to the anterior perforated substance and form its anteromedial and anterolateral boundaries respectively. An intermediate stria is sometimes present. It extends from the centre of trigone to anterior perforated substance where it sinks into the base of olfactory tubercle, which is a small elevation of anterior perforated substance immediately caudal to the olfactory trigone.
پیاز بویایی به صورت مجرای بویایی در عقب ادامه مییابد (شکلهای 18.3 و 18.4). وقتی از عقب ردیابی شود، مجرای بویایی به دو بخش میانی و جانبی تقسیم میشود. نقطه انشعاب گسترش یافته و مثلث بویایی را تشکیل میدهد. بخشهای میانی و جانبی مجرای بویایی ارتباط نزدیکی با ماده سوراخدار قدامی دارند و به ترتیب مرزهای قدامی-داخلی و قدامی-جانبی آن را تشکیل میدهند. گاهی اوقات یک بخش میانی نیز وجود دارد. این عصب از مرکز تریگون تا ماده سوراخدار قدامی امتداد مییابد و در آنجا به پایه توبرکل بویایی فرو میرود، که یک برآمدگی کوچک از ماده سوراخدار قدامی است که بلافاصله بعد از تریگون بویایی قرار دارد.
Posterolaterally the anterior perforated substance is related to the uncus while posteromedially it is bounded by a small bundle of fibres called diagonal band of Broca.
در قسمت خلفی-خارجی، ماده سوراخدار قدامی به استخوان زیرین (Uncus) مرتبط است، در حالی که در قسمت خلفی-داخلی توسط یک دسته کوچک از الیاف به نام نوار مورب بروکا محدود میشود.
Olfactory Cortical Areas
The primary olfactory cortex lies between the anterior perforated substance and the uncus. The entorhinal area comprising uncus and anterior part of the parahippocampal gyrus is often termed secondary olfactory cortex.
نواحی قشر بویایی
قشر بویایی اولیه بین ماده سوراخدار قدامی و آنکوس قرار دارد. ناحیه آنتورینال که شامل آنکوس و قسمت قدامی شکنج پاراهیپوکامپ است، اغلب قشر بویایی ثانویه نامیده میشود.
Neural Pathways for Sense of Olfaction (Fig. 18.5)
The axons of the olfactory receptor cells carrying olfactory sensations from olfactory epithelium (first order sensory neurons) collect to form small bundles (olfactory nerves) which pass through the cribriform plate of the ethmoid bone, to enter the olfactory bulb where they terminate in the olfactory glomeruli. The glomeruli are formed by the axons of receptor cells and dendrites of mitral and tufted cells (these cells form the second order sensory neurons in the olfactory pathway).
مسیرهای عصبی برای حس بویایی (شکل ۱۸.۵)
آکسونهای سلولهای گیرنده بویایی که حس بویایی را از اپیتلیوم بویایی (نورونهای حسی درجه یک) منتقل میکنند، جمع میشوند و دستههای کوچکی (اعصاب بویایی) را تشکیل میدهند که از صفحه مشبک استخوان اتموئید عبور میکنند تا وارد پیاز بویایی شوند و در آنجا در گلومرولهای بویایی خاتمه یابند. گلومرولها توسط آکسونهای سلولهای گیرنده و دندریتهای سلولهای میترال و تافت تشکیل میشوند (این سلولها نورونهای حسی درجه دو را در مسیر بویایی تشکیل میدهند).
Each glomerulus receives impulses from about 26,000 receptor cells and passes this information to the olfactory cortical areas through mitral and tufted cells.
هر گلومرول تکانههایی را از حدود ۲۶۰۰۰ سلول گیرنده دریافت میکند و این اطلاعات را از طریق سلولهای میترال و تافت به نواحی قشری بویایی منتقل میکند.
The most of the axons of mitral cells form the lateral olfactory stria and run to the primary olfactory cortex on the same side which is located between the anterior perforated substance and the uncus on the inferomedial surface of the temporal lobe. The details are shown in Figure 18.4. The others run via intermediate olfactory stria to connect with the olfactory tubercle, and hence with the limbic system.
بیشتر آکسونهای سلولهای میترال، نوارهای بویایی جانبی را تشکیل میدهند و به قشر بویایی اولیه در همان طرف که بین ماده سوراخدار قدامی و آنکوس در سطح تحتانی-میانی لوب گیجگاهی قرار دارد، میروند. جزئیات در شکل 18.4 نشان داده شده است. بقیه از طریق نوارهای بویایی میانی به غده بویایی و در نتیجه به سیستم لیمبیک متصل میشوند.
The axons of the tufted cells run in the medial olfactory stria and cross the midline in the anterior commissure to form synapses with the granule cells in the opposite olfactory bulb.
آکسونهای سلولهای تافتدار در استریای بویایی میانی امتداد مییابند و از خط میانی در رابط قدامی عبور میکنند تا با سلولهای گرانول در پیاز بویایی مقابل سیناپس تشکیل دهند.
It should be noted that in contrast to all other sensory pathways, the fibres of second order sensory neurons reach the primary olfactory cortex directly without relay in one of the thalamic nuclei or its equivalent nucleus like lateral geniculate body.
لازم به ذکر است که برخلاف سایر مسیرهای حسی، فیبرهای نورونهای حسی مرتبه دوم مستقیماً و بدون رله در یکی از هستههای تالاموس یا هسته معادل آن مانند جسم زانویی جانبی به قشر بویایی اولیه میرسند.
The entorhinal area (Brodmann’s area 28) receives few or no tract fibres directly but receives fibres profusely from primary olfactory cortex, hence it is sometimes called secondary olfactory cortex.
ناحیه انتورینال (ناحیه برودمن ۲۸) فیبرهای کمی یا هیچ فیبری را مستقیماً دریافت نمیکند، اما فیبرها را به وفور از قشر بویایی اولیه دریافت میکند، از این رو گاهی اوقات قشر بویایی ثانویه نامیده میشود.
Unique features of the olfactory pathway
• The olfactory pathway consists of only two neurons whereas other conscious sensory pathways consist of at least three neurons.
• Olfactory impulses are transmitted directly to the cerebral cortex without relay in the thalamus.
ویژگیهای منحصر به فرد مسیر بویایی
• مسیر بویایی تنها از دو نورون تشکیل شده است، در حالی که سایر مسیرهای حسی آگاهانه حداقل از سه نورون تشکیل شدهاند.
• تکانههای بویایی مستقیماً به قشر مغز منتقل میشوند، بدون اینکه در تالاموس تقویت شوند.
Clinical Correlation
• A tumour (viz. meningioma) in the floor of anterior cranial fossa may compress the olfactory bulb and tract and interfere with or cause loss of smell to the same side only. It is therefore important to test for smell through each nostril.
همبستگی بالینی
• تومور (مثل مننژیوم) در کف حفره جمجمهای قدامی ممکن است پیاز و مجرای بویایی را فشرده کند و باعث اختلال یا از دست دادن بویایی فقط در همان سمت شود. بنابراین آزمایش بو از طریق هر سوراخ بینی مهم است.
• The sectioning of anterior commissure impairs the sense of smell.
• برش رابط قدامی حس بویایی را مختل میکند.
• The sense of smell contributes significantly to the sense of taste. The sensation of delicious food is in fact an appreciation of aromas through the olfactory system. Persons who have lost their sense of smell, often complain of loss of taste.
• حس بویایی به طور قابل توجهی در حس چشایی نقش دارد. احساس غذای خوشمزه در واقع درک عطرها از طریق سیستم بویایی است. افرادی که حس بویایی خود را از دست دادهاند، اغلب از از دست دادن طعم شکایت دارند.
This phenomenon can be demonstrated by pinching one’s nose to close the nasal passages, while trying to taste something with olfaction blocked, it becomes difficult for a person to distinguish between the taste of a piece of apple and a piece of potato.
این پدیده را میتوان با گرفتن بینی برای بستن مجاری بینی نشان داد، در حالی که فرد سعی میکند چیزی را با حس بویایی مسدود شده بچشد، تشخیص طعم یک تکه سیب و یک تکه سیبزمینی برای فرد دشوار میشود.
• The receptor cells themselves are the first order sensory neurons.
• سلولهای گیرنده، نورونهای حسی درجه یک هستند.
• The receptor cells are exposed to the surface of the body.
• سلولهای گیرنده در معرض سطح بدن قرار دارند.
Visual System
The vision (sight) is the most important special sense in humans. The visual system transmits sense of vision (sight) from retina of eyeball to the visual cortex. The following structures are included in the visual system:
سیستم بینایی
بینایی (یا حس بینایی) مهمترین حس ویژه در انسان است. سیستم بینایی، حس بینایی (بینایی) را از شبکیه چشم به قشر بینایی منتقل میکند. ساختارهای زیر در سیستم بینایی گنجانده شدهاند:
• Retina
• Optic nerve
• Optic chiasma
• Optic tract
• Lateral geniculate body
• Optic radiation (geniculocalcarine tract), and
• Visual cortex
• شبکیه
• عصب بینایی
• کیاسمای بینایی
• راه بینایی
• جسم زانویی جانبی
• تشعشعات بینایی (راه ژنیکولوکالکارینی) و
• قشر بینایی
Retina
The retina forms the inner photosensitive coat of the eyeball. It consists of two layers, an inner neural layer and an outer layer of pigment epithelium (Fig. 18.6).
شبکیه
شبکیه لایه داخلی حساس به نور کره چشم را تشکیل میدهد. این لایه از دو لایه تشکیل شده است، یک لایه عصبی داخلی و یک لایه بیرونی اپیتلیوم رنگدانهای (شکل 18.6).

FIG. 18.6 Three basic layers of retina and their constituent cells. The arrow (on the left side) indicates the direction of light falling on the retina. It is important to note that several rods and cones converge on a single bipolar neuron and several bipolar neurons activate one ganglion cell. The one-to-one relationship between rods and cones, bipolar neurons and ganglion cells shown in this figure is only for the sake of simplicity.
شکل ۱۸.۶ سه لایه اساسی شبکیه و سلولهای تشکیلدهنده آنها. فلش (در سمت چپ) جهت نور تابیده شده به شبکیه را نشان میدهد. لازم به ذکر است که چندین میله و مخروط روی یک نورون دوقطبی همگرا میشوند و چندین نورون دوقطبی یک سلول گانگلیونی را فعال میکنند. رابطه یک به یک بین میلهها و مخروطها، نورونهای دوقطبی و سلولهای گانگلیونی که در این شکل نشان داده شده است، فقط برای سادگی است.
Neural layer
The neural layer contains three basic layers of cells:
• An outer layer of rod and cone cells
• A middle layer of bipolar cells
• An inner layer of ganglion cells
لایه عصبی
لایه عصبی شامل سه لایه اساسی از سلولها است:
• یک لایه بیرونی از سلولهای میلهای و مخروطی
• یک لایه میانی از سلولهای دوقطبی
• یک لایه داخلی از سلولهای گانگلیون
The other cells are association neurons and neuroglial cells.
سایر سلولها، نورونهای ارتباطی و سلولهای نوروگلیال هستند.
Rod and cone cells
The rods and cones (around 150 million in number) are modified neurons and serve as photoreceptors. Both consist of an outer and inner portion, the former being light sensitive and contains photopigments, rhodopsin in case of rods and iodopsin in case of cones which convert the stimulus of light into a nerve impulse. The outer portion is rod-shaped in case of rods and cone-shaped in case of cones, hence the names rods and cones. The inner portions of rods and cones are slender and are termed rod and cone fibres respectively. The cones respond better to the bright light and are responsible for visual acuity and colour vision. They are most numerous in the central region of the retina. Rods on the other hand, predominate in the peripheral part of the retina. They respond to poor light and are important for peripheral vision.
سلولهای میلهای و مخروطی
میلهها و مخروطها (حدود ۱۵۰ میلیون عدد) نورونهای اصلاحشده هستند و به عنوان گیرندههای نوری عمل میکنند. هر دو از یک بخش بیرونی و داخلی تشکیل شدهاند که بخش بیرونی حساس به نور است و حاوی رنگدانههای نوری، رودوپسین در مورد میلهها و یدوپسین در مورد مخروطها است که محرک نور را به یک تکانه عصبی تبدیل میکند. بخش بیرونی در مورد میلهها میلهای شکل و در مورد مخروطها مخروطی شکل است، از این رو نامهای میلهای و مخروطی به آنها داده شده است. بخشهای داخلی میلهها و مخروطها باریک هستند و به ترتیب فیبرهای میلهای و مخروطی نامیده میشوند. مخروطها به نور روشن بهتر پاسخ میدهند و مسئول حدت بینایی و دید رنگی هستند. آنها بیشتر در ناحیه مرکزی شبکیه وجود دارند. از سوی دیگر، میلهها در قسمت محیطی شبکیه غالب هستند. آنها به نور کم پاسخ میدهند و برای دید محیطی مهم هستند.
Bipolar cells
Bipolar cells are bipolar neurons, interposed between the photoreceptor cells and the ganglion cells.
سلولهای دوقطبی
سلولهای دوقطبی، نورونهای دوقطبی هستند که بین سلولهای گیرنده نوری و سلولهای گانگلیونی قرار گرفتهاند.
Ganglion cells
Ganglion cells are large multipolar neurons forming the last retinal link in the visual pathway. The axons of ganglion cells form a layer of nerve fibres adjacent to the vitreous humour. These fibres converge towards the rounded area (optic disc) from all directions where they pierce the choroid and sclera, about 3 or 4 mm to the nasal side of the posterior pole of the eyeball and constitute the optic nerve.
سلولهای گانگلیون
سلولهای گانگلیون، نورونهای چندقطبی بزرگی هستند که آخرین حلقه شبکیه در مسیر بینایی را تشکیل میدهند. آکسونهای سلولهای گانگلیون، لایهای از الیاف عصبی را در مجاورت زجاجیه تشکیل میدهند. این الیاف از همه جهات به سمت ناحیه گرد (دیسک بینایی) همگرا میشوند، جایی که مشیمیه و صلبیه را سوراخ میکنند، حدود ۳ یا ۴ میلیمتر به سمت بینی قطب خلفی کره چشم و عصب بینایی را تشکیل میدهند.
The optic disc, about 1.5 mm in diameter, is slightly (3 mm) medial to the posterior pole. It is insensitive to light as it has no sensory receptors (blind spot). It represents the point of exit of the optic nerve fibres. Its central part is pierced by central artery of retina and tributaries of central vein of retina. The optic disc may be regarded as a window to the brain and its examination is an important step in the diagnosis and prognosis of diseases with vascular and neurological implications.
دیسک بینایی، با قطر حدود ۱.۵ میلیمتر، کمی (۳ میلیمتر) نسبت به قطب خلفی داخلیتر است. به دلیل نداشتن گیرندههای حسی (نقطه کور)، نسبت به نور حساس نیست. این نقطه، محل خروج الیاف عصب بینایی است. قسمت مرکزی آن توسط شریان مرکزی شبکیه و شاخههای ورید مرکزی شبکیه سوراخ میشود. دیسک بینایی را میتوان به عنوان دریچهای به مغز در نظر گرفت و بررسی آن گامی مهم در تشخیص و پیشآگهی بیماریهایی با پیامدهای عروقی و عصبی است.
The macula lutea is a yellowish oval area of 6 mm in diameter near the centre of the posterior part of the retina. It is in line with the visual axis. The name macula lutea or yellow spot is derived from the presence of yellow pigment (xanthophyll) among the nerve cells in this region.
لکه زرد (ماکولا لوتئا) ناحیهای بیضیشکل مایل به زرد با قطر ۶ میلیمتر است که در نزدیکی مرکز قسمت خلفی شبکیه قرار دارد. این ناحیه در راستای محور بینایی است. نام لکه زرد یا ماکولا لوتئا از وجود رنگدانه زرد (گزانتوفیل) در بین سلولهای عصبی این ناحیه گرفته شده است.
A small depression in the centre of the macula lutea is called fovea centralis. The fovea is about 1.5 mm in diameter and is separated from the edge of optic disc by a distance of 3 mm. The visual acuity is maximum at the fovea (i.e. clearest vision). The fovea is believed to contain only cone receptors.
یک فرورفتگی کوچک در مرکز ماکولا لوتئا، حفره مرکزی نامیده میشود. قطر حفره حدود ۱.۵ میلیمتر است و از لبه دیسک بینایی با فاصله ۳ میلیمتر جدا میشود. حداکثر حدت بینایی در حفره بینایی است (یعنی واضحترین دید). اعتقاد بر این است که حفره بینایی فقط حاوی گیرندههای مخروطی است.
Clinical Correlation
The retina can be examined directly through an ophthalmoscope and following features are seen in its posterior part (fundus): (a) optic disc/optic papilla/ blind spot, (b) central artery of retina emerging from optic disc, (c) tributaries of central vein of retina converging towards the optic disc, (d) macula lutea, and (e) fovea centralis.
همبستگی بالینی
شبکیه را میتوان مستقیماً از طریق افتالموسکوپ معاینه کرد و ویژگیهای زیر در قسمت خلفی آن (فوندوس) مشاهده میشود: (الف) دیسک بینایی/پاپیلای بینایی/نقطه کور، (ب) شریان مرکزی شبکیه که از دیسک بینایی خارج میشود، (ج) شاخههای ورید مرکزی شبکیه که به سمت دیسک بینایی همگرا میشوند، (د) ماکولا لوتئا، و (ه) حفره مرکزی.
N.B. The optic disc looks pink in colour due to the presence of capillary vessels. In optic atrophy the disc looks white as the capillary vessels disappear.
توجه: دیسک بینایی به دلیل وجود رگهای مویرگی صورتی رنگ به نظر میرسد. در آتروفی بینایی، دیسک با ناپدید شدن رگهای مویرگی، سفید به نظر میرسد.
Pigment epithelium
Pigment epithelium consists of a single layer of cells containing melanin pigment. Pigment epithelium reinforces the light absorbing proportion of the choroid to reduce the scattering of light within the eye.
اپیتلیوم رنگدانهای
اپیتلیوم رنگدانهای از یک لایه سلول حاوی رنگدانه ملانین تشکیل شده است. اپیتلیوم رنگدانهای، نسبت جذب نور مشیمیه را تقویت میکند تا پراکندگی نور در داخل چشم را کاهش دهد.
Clinical Correlation
While pigment epithelium is firmly attached to the choroid but it is not so firmly attached to the neural layer of retina. The pigment epithelium of retina develops from an outer layer and the neural layer from the inner layer of the optic cup. As a result there remains a potential space between the two layers. Therefore the detachment of retina which may follow a blow on the eye, consists of separation of neural layer from pigment epithelium, i.e. at the junction of two layers of optic cup.
همبستگی بالینی
در حالی که اپیتلیوم رنگدانهای محکم به مشیمیه متصل است، اما به لایه عصبی شبکیه چندان محکم متصل نیست. اپیتلیوم رنگدانهای شبکیه از یک لایه بیرونی و لایه عصبی از لایه داخلی جام بینایی ایجاد میشود. در نتیجه، بین دو لایه فضای بالقوهای باقی میماند. بنابراین، جدا شدن شبکیه که ممکن است پس از ضربه به چشم رخ دهد، شامل جداسازی لایه عصبی از اپیتلیوم رنگدانهای، یعنی در محل اتصال دو لایه جام بینایی است.
The visual field and retinal quadrants
When one looks straight ahead with eyes fixed, that part of external world which can be seen with each eye is called visual field of that eye. Thus it is the area within which an object can be seen while the eye fixes on a spot of light or object. Laterally it extends up to 104 degree and on nasal side 65 degree. In front there is a cone-shaped area in which the visual fields of two eyes overlap. Therefore, area seen by one eye and that seen by both the eyes is more or less same except a small area that can be seen only by the eye of that side (Fig. 18.7).
میدان بینایی و ربعهای شبکیه
وقتی کسی با چشمان ثابت مستقیم به جلو نگاه میکند، آن بخش از دنیای بیرون که با هر چشم دیده میشود، میدان بینایی آن چشم نامیده میشود. بنابراین، این ناحیهای است که در آن میتوان یک شیء را در حالی که چشم روی یک نقطه نور یا شیء ثابت شده است، دید. این میدان در طرفین تا 104 درجه و در سمت بینی تا 65 درجه امتداد دارد. در جلو، ناحیهای مخروطی شکل وجود دارد که در آن میدانهای بینایی دو چشم همپوشانی دارند. بنابراین، ناحیهای که توسط یک چشم دیده میشود و ناحیهای که توسط هر دو چشم دیده میشود، کم و بیش یکسان است، به جز یک ناحیه کوچک که فقط توسط چشم آن طرف دیده میشود (شکل 18.7).
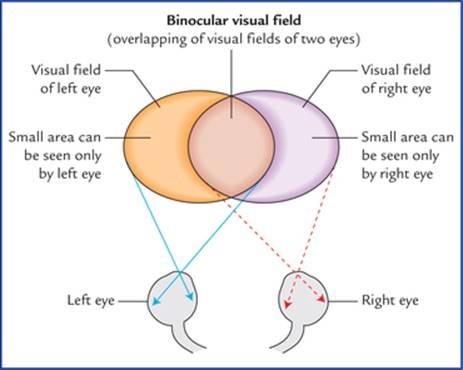
FIG. 18.7 Visual fields of the two eyes and binocular visual field.
شکل ۱۸.۷ میدانهای بینایی دو چشم و میدان بینایی دو چشمی.
For the sake of convenience of description, the visual field is conventionally divided into right and left halves. Each half is further divided into an upper and a lower half, so that visual field is described to consist of four quadrants (Fig. 18.8).
برای راحتی توصیف، میدان بینایی به طور مرسوم به نیمههای راست و چپ تقسیم میشود. هر نیمه نیز به نیمههای بالایی و پایینی تقسیم میشود، به طوری که میدان بینایی به صورت چهار ربع توصیف میشود (شکل ۱۸.۸).

FIG. 18.8 Scheme to show the projection of retina on the lateral geniculate body and the visual cortex. The details areshown only for the right halves of the two retinae.
شکل ۱۸.۸ طرحی برای نشان دادن تصویر شبکیه بر روی جسم زانویی جانبی و قشر بینایی. جزئیات فقط برای نیمههای راست دو شبکیه نشان داده شده است.
In a similar manner the retina is also divided into four quadrants (Fig. 18.8). First each retina is divided into nasal and temporal halves by a vertical line passing through the fovea centralis. Then a horizontal line also passing through the fovea, divides each half of retina into upper and lower quadrants. The macular area (responsible for most acute vision) is represented separately from the peripheral parts of the retina. Light rays can enter the eye only through the pupil and since they travel in straight lines, it is obvious that objects of temporal field of vision are perceived by the nasal half of the retina whereas those in the nasal half are perceived by the temporal half of the retina.
به طور مشابه، شبکیه نیز به چهار ربع تقسیم میشود (شکل ۱۸.۸). ابتدا هر شبکیه توسط یک خط عمودی که از گودی مرکزی عبور میکند، به نیمههای بینی و گیجگاهی تقسیم میشود. سپس یک خط افقی که آن هم از گودی مرکزی عبور میکند، هر نیمه شبکیه را به ربعهای بالایی و پایینی تقسیم میکند. ناحیه ماکولا (مسئول اکثر بیناییهای دقیق) به طور جداگانه از قسمتهای محیطی شبکیه نمایش داده میشود. پرتوهای نور فقط میتوانند از طریق مردمک وارد چشم شوند و از آنجایی که در خطوط مستقیم حرکت میکنند، بدیهی است که اشیاء میدان دید گیجگاهی توسط نیمه بینی شبکیه درک میشوند، در حالی که اشیاء در نیمه بینی توسط نیمه گیجگاهی شبکیه درک میشوند.
Central projection of retinal areas
The following general rules concerning central projection of retina should be carefully understood before discussing the components of the optic pathway:
تصویر مرکزی نواحی شبکیه
قبل از بحث در مورد اجزای مسیر بینایی، باید قوانین کلی زیر در مورد تصویر مرکزی شبکیه به دقت درک شود:
• Fibres from right halves of two retinae terminate in the right lateral geniculate body and those from left halves terminate in the left lateral geniculate body. The visual information from lateral geniculate body of each side is relayed into the visual cortex of the corresponding side.
• فیبرهای نیمه راست دو شبکیه به جسم زانویی جانبی راست و فیبرهای نیمه چپ به جسم زانویی جانبی چپ ختم میشوند. اطلاعات بینایی از جسم زانویی جانبی هر طرف به قشر بینایی طرف مربوطه منتقل میشود.
• Fibres from peripheral parts of retina end in the ventral two-thirds of the lateral geniculate body and those from the macula into the dorsal one-third of lateral geniculate body. Fibres from upper quadrants end in the medial half and those from lower quadrants in the lateral half of the geniculate body.
• فیبرهای قسمتهای محیطی شبکیه به دو سوم شکمی جسم زانویی جانبی و فیبرهای ماکولا به یک سوم پشتی جسم زانویی جانبی ختم میشوند. فیبرهای ربعهای بالایی به نیمه میانی و فیبرهای ربعهای پایینی به نیمه جانبی جسم زانویی ختم میشوند.
• The fibres from medial half of the lateral geniculate body relay into the upper lip of the calcarine sulcus and those from the lateral half into the lower lip of calcarine sulcus of the occipital lobe. Peripheral and macular quadrants maintain the same relationship as that in the lateral geniculate body, i.e. peripheral quadrants occupy the anterior two-third of visual cortex and macular quadrants occupy the posterior one-third of visual cortex.
• فیبرهای نیمه داخلی جسم زانویی جانبی به لب بالایی شیار کالکارین و فیبرهای نیمه جانبی به لب پایینی شیار کالکارین لوب پسسری منتقل میشوند. ربعهای محیطی و ماکولا همان رابطهای را که در جسم زانویی جانبی وجود دارد، حفظ میکنند، یعنی ربعهای محیطی دو سوم قدامی قشر بینایی و ربعهای ماکولا یک سوم خلفی قشر بینایی را اشغال میکنند.
• The portion of lateral geniculate body and visual cortex receiving fibres from macula (only 6 mm in diameter) are disproportionately larger.
• بخشی از جسم زانویی جانبی و قشر بینایی که فیبرها را از ماکولا دریافت میکنند (فقط ۶ میلیمتر قطر دارند) به طور نامتناسبی بزرگتر هستند.
• Thus specific points on the retina project to the specific points in the lateral geniculate body and similarly specific points in the lateral geniculate body are projected to the specific points in the visual cortex. Therefore, point to point relationship is maintained between the retinae and the visual cortex.
• بنابراین نقاط خاصی روی شبکیه به نقاط خاصی در جسم زانویی جانبی منعکس میشوند و به طور مشابه نقاط خاصی در جسم زانویی جانبی به نقاط خاصی در قشر بینایی منعکس میشوند. بنابراین، رابطه نقطه به نقطه بین شبکیه و قشر بینایی حفظ میشود.
Figure 18.8 represents the above mentioned rules.
شکل ۱۸.۸ قوانین ذکر شده در بالا را نشان میدهد.
Optic Nerve, Optic Chiasma and Optic Tract
The optic nerve comes out by piercing the sclera 3 mm medial to the posterior pole of the eyeball. In man it contains about one million fibres, which are the axons of ganglion cells of retina. It passes through the optic canal to reach the middle cranial fossa, where it approaches and joins its counterpart of the other side to form the optic chiasma above the roof of pituitary fossa. Posteriorly the chiasma divides into two optic tracts which after winding round the cerebral peduncles end in the lateral geniculate body of the corresponding side. The optic nerve is about 4 cm in length. Throughout its course the nerve is surrounded by the extensions of the meninges, i.e. pia, arachnoid and dura mater. The meningeal extensions around the nerve fuse with the scleral coat of the eyeball. The subarachnoid space around the nerve is continuous with that of brain. The fibres arising from macula are numerous and form the papillomacular bundle.
عصب بینایی، کیاسمای بینایی و مسیر بینایی
عصب بینایی با سوراخ کردن صلبیه در فاصله ۳ میلیمتری داخلی قطب خلفی کره چشم، خارج میشود. در انسان، این عصب حاوی حدود یک میلیون فیبر است که آکسونهای سلولهای گانگلیونی شبکیه هستند. این عصب از کانال بینایی عبور میکند تا به حفره جمجمهای میانی برسد، جایی که به همتای خود در طرف دیگر نزدیک شده و به آن میپیوندد تا کیاسمای بینایی را در بالای سقف حفره هیپوفیز تشکیل دهد. در قسمت خلفی، کیاسمای بینایی به دو مسیر بینایی تقسیم میشود که پس از پیچیدن دور پایههای مغزی، در جسم زانویی جانبی طرف مربوطه به پایان میرسند. عصب بینایی حدود ۴ سانتیمتر طول دارد. در طول مسیر خود، عصب توسط امتدادهای مننژ، یعنی نرمشامه، عنکبوتیه و سختشامه احاطه شده است. امتدادهای مننژی در اطراف عصب با پوشش صلبیه کره چشم ترکیب میشوند. فضای زیر عنکبوتیه اطراف عصب، در امتداد فضای مغز قرار دارد. فیبرهای منشعب از ماکولا متعدد هستند و دسته پاپیلوماکولار را تشکیل میدهند.
The fibres of optic nerve arising in the nasal half of each retina (nasal fibres) enter the optic tract of the opposite side after crossing in the optic chiasma.
فیبرهای عصب بینایی که از نیمه بینی هر شبکیه (فیبرهای بینی) منشأ میگیرند، پس از عبور از کیاسمای بینایی، وارد مجرای بینایی طرف مقابل میشوند.
It is to be noted that immediately after crossing, these fibres loop forward for a short distance in the optic nerve. Therefore, a lesion affecting optic nerve just in front of optic chiasma leads to temporal field defect in opposite eye and blindness on the side of lesion. Fibres from temporal half of each retina (temporal fibres) enter the optic tract of the same side. Thus due to the decussation of nasal fibres of optic nerves in the optic chiasma, each optic tract contains the temporal fibres of the same side and nasal fibres of the opposite side.
لازم به ذکر است که بلافاصله پس از عبور، این فیبرها برای مسافت کوتاهی در عصب بینایی به سمت جلو حلقه میزنند. بنابراین، ضایعهای که عصب بینایی را درست در جلوی کیاسمای بینایی تحت تأثیر قرار میدهد، منجر به نقص میدان گیجگاهی در چشم مقابل و نابینایی در سمت ضایعه میشود. فیبرهای نیمه گیجگاهی هر شبکیه (فیبرهای گیجگاهی) وارد مجرای بینایی همان طرف میشوند. بنابراین به دلیل تقاطع فیبرهای بینی اعصاب بینایی در کیاسمای بینایی، هر مجرای بینایی شامل فیبرهای گیجگاهی همان طرف و فیبرهای بینی طرف مقابل است.
Lateral Geniculate Body
The structure of lateral geniculate body has already been discussed on page 131. This body (or nucleus) is the site of termination of all the optic nerve fibres except the few which reach and relay in the pretectal region and superior colliculus of the midbrain. These fibres form an afferent limb of the pupillary light reflex. Recent studies show that geniculate body receives afferents from the visual cortex also.
جسم زانویی جانبی
ساختار جسم زانویی جانبی قبلاً در صفحه ۱۳۱ مورد بحث قرار گرفته است. این جسم (یا هسته) محل پایان تمام فیبرهای عصب بینایی به جز تعداد کمی است که به ناحیه پرهتکتال و کولیکولوس فوقانی مغز میانی میرسند و در آنجا رله میشوند. این فیبرها یک شاخه آوران رفلکس نور مردمک را تشکیل میدهند. مطالعات اخیر نشان میدهد که جسم زانویی آورانهایی را از قشر بینایی نیز دریافت میکند.
Geniculocalcarine Tract
Fibres arising from cells of lateral geniculate body constitute the geniculocalcarine tract. First these fibres pass through the retrolentiform part of the internal capsule, then they fan out to form the optic radiation which end in the visual cortex (area 17, 18 and 19) of the occipital lobe. Some of the geniculocalcarine fibres loop forward over the temporal (inferior) horn of lateral ventricle before curving posteriorly towards the occipital lobe and constitute the temporal loop (Meyer’s loop) of geniculocalcarine tract. Fibres forming this loop arise from the lateral part of the lateral geniculate body.
راه ژنیکولوکالکارین
فیبرهایی که از سلولهای جسم زانویی جانبی منشأ میگیرند، راه ژنیکولوکالکارین را تشکیل میدهند. ابتدا این فیبرها از قسمت خلفی کپسول داخلی عبور میکنند، سپس به صورت بادبزنی بیرون میزنند و تابش نوری را تشکیل میدهند که در قشر بینایی (ناحیه ۱۷، ۱۸ و ۱۹) لوب پسسری خاتمه مییابد. برخی از فیبرهای ژنیکولوکالکارین قبل از اینکه به سمت خلف به سمت لوب پسسری خم شوند، به سمت جلو و روی شاخ گیجگاهی (تحتانی) بطن جانبی حلقه میزنند و حلقه گیجگاهی (حلقه مایر) راه ژنیکولوکالکارین را تشکیل میدهند. فیبرهای تشکیلدهنده این حلقه از قسمت جانبی جسم زانویی خارجی منشأ میگیرند.
Visual Cortex
Visual cortex includes primary and secondary visual areas. These are described in detail on pages 152 and 153.
قشر بینایی
قشر بینایی شامل نواحی بینایی اولیه و ثانویه است. این نواحی به تفصیل در صفحات ۱۵۲ و ۱۵۳ شرح داده شدهاند.
Neural Pathway for Vision (Fig. 18.9)
The neural pathway for vision is a three order neuronal pathway.
مسیر عصبی بینایی (شکل ۱۸.۹)
مسیر عصبی بینایی یک مسیر نورونی سهگانه است.

FIG. 18.9 The visual pathway and sites of various lesions affecting it. The black areas of inserts on the right side indicate the visual field defects: 1 = complete blindness of right eye, 2 = bitemporal hemianopia, 3 = right nasal hemianopia, 4&5 = left homonymous hemianopia, 6 = homonymous hemianopia with macular sparing, 7 = contralateral inferior quadrantic hemianopia, 8 = contralateral superior quadrantic hemianopia.
شکل ۱۸.۹ مسیر بینایی و محل ضایعات مختلف مؤثر بر آن. نواحی سیاه درجها در سمت راست، نقایص میدان بینایی را نشان میدهند: ۱ = نابینایی کامل چشم راست، ۲ = همیآنوپی دو گیجگاهی، ۳ = همیآنوپی بینی راست، ۴ و ۵ = همیآنوپی همنام چپ، ۶ = همیآنوپی همنام با حفظ ماکولا، ۷ = همیآنوپی ربع تحتانی طرف مقابل، ۸ = همیآنوپی ربع فوقانی طرف مقابل.
The first order sensory (primary) neurons carrying visual sensations are bipolar cells of the retina. Their dendrites synapse with photoreceptors (rods and cones) and axons with the dendrites of ganglion cells.
نورونهای حسی (اولیه) درجه اول که حامل حس بینایی هستند، سلولهای دوقطبی شبکیه هستند. دندریتهای آنها با گیرندههای نوری (میلهای و مخروطی) و آکسونها با دندریتهای سلولهای گانگلیونی سیناپس میکنند.
The second order sensory (secondary) neurons are the multipolar neurons (generally referred by most authors as ganglion cells) whose axons run along the optic nerve to the optic chiasma, where fibres from nasal half of retina cross to the opposite side and travel through the opposite optic tract to terminate in the opposite lateral geniculate body. The fibres from temporal half remain uncrossed in the optic chiasma and run in optic tract of the same side to terminate in the ipsilateral geniculate body. The fibres from macula lutea behaves in the exactly same manner.
نورونهای حسی (ثانویه) مرتبه دوم، نورونهای چندقطبی (که عموماً توسط اکثر نویسندگان به عنوان سلولهای گانگلیون نامیده میشوند) هستند که آکسونهای آنها در امتداد عصب بینایی تا کیاسمای بینایی امتداد دارند، جایی که فیبرهای نیمه بینی شبکیه به طرف مقابل عبور میکنند و از طریق دستگاه بینایی مقابل حرکت میکنند تا در جسم زانویی جانبی مقابل خاتمه یابند. فیبرهای نیمه گیجگاهی در کیاسمای بینایی به صورت متقاطع باقی میمانند و در دستگاه بینایی همان طرف امتداد مییابند تا در جسم زانویی همان طرف خاتمه یابند. فیبرهای ماکولا لوتئا نیز دقیقاً به همین شکل رفتار میکنند.
The cell bodies of third order sensory (tertiary) neurons are located in the lateral geniculate body. Their axons form the optic radiations which project to the visual cortex.
جسم سلولی نورونهای حسی درجه سوم (سوم) در جسم زانویی جانبی قرار دارند. آکسونهای آنها پرتوهای نوری را تشکیل میدهند که به قشر بینایی تابیده میشوند.
Clinical Correlation
The blindness is conventionally described with reference to the field of vision rather than with reference to the parts of retina. Loss of vision in one-half of the visual field (right or left) is termed hemianopia. If same halves of visual fields are affected in both the eyes, it is called homonymous hemianopia. On the other hand if different halves of visual fields are affected, the term heteronymous hemianopia is used.
همبستگی بالینی
نابینایی معمولاً با اشاره به میدان بینایی توصیف میشود، نه با اشاره به بخشهای شبکیه. از دست دادن بینایی در نیمی از میدان بینایی (راست یا چپ) همیآنوپی نامیده میشود. اگر نیمههای یکسانی از میدانهای بینایی در هر دو چشم تحت تأثیر قرار گیرند، همیآنوپی همنام نامیده میشود. از سوی دیگر، اگر نیمههای مختلف میدانهای بینایی تحت تأثیر قرار گیرند، از اصطلاح همیآنوپی ناهمنام استفاده میشود.
The visual field defects caused by lesions affecting different parts of the visual pathway are described in Figure 18.9.
نقصهای میدان بینایی ناشی از ضایعاتی که بخشهای مختلف مسیر بینایی را تحت تأثیر قرار میدهند، در شکل 18.9 شرح داده شده است.
1. Injury of optic nerve of one side will result in total blindness in the eye of that side.
1. آسیب عصب بینایی یک طرف منجر به نابینایی کامل در چشم آن طرف میشود.
2. Damage of central part of optic chiasma (as may occur from pituitary tumour) interrupts the crossing fibres from nasal halves of two retinae, and results in bitemporal heteronymous hemianopia (tunnel vision).
2. آسیب به قسمت مرکزی کیاسمای بینایی (همانطور که ممکن است در اثر تومور هیپوفیز رخ دهد) فیبرهای عبوری از نیمههای بینی دو شبکیه را قطع میکند و منجر به همیآنوپی هترونیموس دو گیجگاهی (دید تونلی) میشود.
3. Destruction of lateral part of optic chiasma on one side (as may occur from aneurysm of internal carotid artery) produces a unilateral nasal hemianopia on the side of lesion.
3. تخریب قسمت جانبی کیاسمای بینایی در یک طرف (همانطور که ممکن است در اثر آنوریسم شریان کاروتید داخلی رخ دهد) باعث همیآنوپی بینی یک طرفه در سمت ضایعه میشود.
4 & 5. Complete destruction of optic tract (4), geniculocalcarine tract (5) on one side results in blindness in opposite half of field of vision (i.e. nasal field of ipsilateral eye and temporal field of contralateral eye). This type of defect is described as right or left homonymous hemianopia, depending upon the side of lesion.
4 و 5. تخریب کامل مسیر بینایی (4)، مسیر ژنیکولوکالکارین (5) در یک طرف منجر به نابینایی در نیمه مخالف میدان دید (یعنی میدان بینی چشم همان طرف و میدان گیجگاهی چشم مقابل) میشود. این نوع نقص، بسته به سمت ضایعه، به عنوان همیآنوپی هومونیموس راست یا چپ توصیف میشود.
6. Lesions involving visual cortex produce blindness in opposite half of field of vision (homonymous hemianopia), with sparing of macular vision.
6. ضایعاتی که قشر بینایی را درگیر میکنند، باعث نابینایی در نیمه مخالف میدان دید (همیآنوپی همنام) میشوند، با حفظ دید ماکولا.
7. Destruction of upper tip of calcarine sulcus on one side produces visual loss in lower quadrant of opposite field of vision (contralateral inferior quadrantic anopia).
7. تخریب نوک بالایی شیار کالکارین در یک طرف باعث از دست دادن بینایی در ربع تحتانی میدان دید مخالف میشود (آنوپی ربع تحتانی طرف مقابل).
8. Destruction of lower lip of calcarine sulcus on one side produces visual loss in superior quadrant of opposite field of vision (contralateral superior quadrantic anopia).
8. تخریب لب تحتانی شیار کالکارین در یک طرف باعث از دست دادن بینایی در ربع فوقانی میدان دید مخالف میشود (آنوپی ربع فوقانی طرف مقابل).
N.B.
• A temporal lobe lesion involving temporal loop (also called Meyer’s loop) of geniculocalcarine tract on one side causes defect in superior quadrant of opposite field of vision (superior quadrantic anopia).
• Lesions anterior to lateral geniculate body also interrupts the fibres responsible for the pupillary light reflex.
توجه:
• ضایعه لوب گیجگاهی که حلقه گیجگاهی (که حلقه مایر نیز نامیده میشود) از مسیر ژنیکولوکالکارینی را در یک طرف درگیر میکند، باعث نقص در ربع فوقانی میدان دید مقابل میشود (آنوپی ربع فوقانی).
• ضایعات قدامی جسم زانویی جانبی نیز فیبرهای مسئول رفلکس نوری مردمک را قطع میکنند.
Visual Reflexes
Light reflex or pupillary reflex
When light is shown to one eye, normally the pupils of both the eyes constrict. The constriction of pupil upon which the light is shown is termed direct light reflex and the constriction of the pupil of the other eye (even though no light fell upon that eye) is termed indirect or consensual light reflex.
رفلکسهای بینایی
رفلکس نوری یا رفلکس مردمک
وقتی نور به یک چشم تابیده میشود، معمولاً مردمکهای هر دو چشم تنگ میشوند. انقباض مردمک که نور به آن تابیده میشود، رفلکس نوری مستقیم نامیده میشود و انقباض مردمک چشم دیگر (حتی اگر هیچ نوری به آن چشم نیفتد) رفلکس نوری غیرمستقیم یا توافقی نامیده میشود.
Pathway of the reflex (Fig. 18.10)
Afferent limb: The visual impulses travel through the optic nerve, optic chiasma and optic tract to terminate in the pretectal nucleus. Then the impulses from pretectal nucleus pass to the parasympathetic nuclei (Edinger-Westphal nuclei) of the IIIrd cranial nerve of both sides.
مسیر رفلکس (شکل 18.10)
عضو آوران: تکانههای بینایی از طریق عصب بینایی، کیاسمای بینایی و مجرای بینایی حرکت میکنند تا در هسته پرهتکتال خاتمه یابند. سپس تکانهها از هسته پرهتکتال به هستههای پاراسمپاتیک (هستههای ادینگر-وستفال) عصب جمجمهای سوم هر دو طرف منتقل میشوند.

FIG. 18.10 Pathways for light reflex (direct and consensual).
شکل ۱۸.۱۰ مسیرهای رفلکس نور (مستقیم و توافقی).
Efferent limb: The impulses from the Edinger-Westphal nucleus pass bilaterally along the oculomotor nerve to the ciliary ganglion. The postganglionic parasympathetic fibres travel along the short ciliary nerves to supply the sphincter pupillae muscle of the iris leading to constriction of pupils of the both eyes.
شاخه وابران: تکانههای هسته ادینگر-وستفال به صورت دو طرفه در امتداد عصب اوکولوموتور به گانگلیون مژگانی منتقل میشوند. فیبرهای پاراسمپاتیک پسگانگلیونی در امتداد اعصاب مژگانی کوتاه حرکت میکنند تا عضله اسفنکتر مردمک عنبیه را تغذیه کنند و منجر به انقباض مردمک هر دو چشم شوند.
The afferent and efferent limbs of light reflex are depicted in Flowchart 18.1.
شاخههای آوران و وابران رفلکس نور در فلوچارت ۱۸.۱ نشان داده شده است.

FLOWCHART 18.1 Light reflex.
نمودار جریان ۱۸.۱ رفلکس نور.
Accommodation reflex
When the eyes are focused from a distant to a near object, three reactions take place in the eyes, viz.
رفلکس تطابق
هنگامی که چشمها از یک شیء دور به یک شیء نزدیک متمرکز میشوند، سه واکنش در چشمها رخ میدهد، از جمله:
• There is constriction of the pupils due to action of sphincter pupillae to restrict the light waves to the thickest central part of the lens.
• There is thickening of the lenses due to contraction of the ciliary muscles, to increase the refractive index of the lenses.
• There is convergence of both the eyeballs, due to the contraction of medial recti, to bring about the convergence of the ocular axes.
• انقباض مردمکها به دلیل عملکرد مردمکهای اسفنکتر برای محدود کردن امواج نور به ضخیمترین قسمت مرکزی عدسی.
• ضخیم شدن عدسیها به دلیل انقباض عضلات مژگانی، برای افزایش ضریب شکست عدسیها.
• همگرایی هر دو کره چشم، به دلیل انقباض عضلات راست داخلی، برای ایجاد همگرایی محورهای چشمی.
These three reactions together constitute the accommodation or near reflex.
این سه واکنش در کنار هم، رفلکس تطابق یا رفلکس نزدیک را تشکیل میدهند.
Pathways of the accommodation reflex
Afferent limb: The visual impulses travel through the optic nerve, optic chiasma, optic tract, lateral geniculate body, and the optic radiation to the visual cortex.
مسیرهای رفلکس تطابق
عضو آوران: تکانههای بینایی از طریق عصب بینایی، کیاسمای بینایی، دستگاه بینایی، جسم زانویی جانبی و تابش بینایی به قشر بینایی میرسند.
From visual cortex the impulses reach the ‘frontal eye field’ through association fasciculus, the superior longitudinal fasciculus.
از قشر بینایی، تکانهها از طریق فاسیکولوس ارتباطی، فاسیکولوس طولی فوقانی، به «میدان چشم جلویی» میرسند.
Efferent limb: From ‘frontal eye field’ the corticonuclear fibres project to the oculomotor nerve nuclei of both the sides. From here the impulses travel through the oculomotor nerves to supply, all the three muscles, viz. sphincter pupillae, ciliary muscle and medial rectus of the eyeballs whose active action is needed for accommodation.
عضو وابران: از «میدان چشم جلویی»، فیبرهای قشر هستهای به هستههای عصب حرکتی چشم در هر دو طرف امتداد مییابند. از اینجا، تکانهها از طریق اعصاب حرکتی چشم به هر سه عضله، یعنی مردمک اسفنکتر، عضله مژگانی و عضله راست داخلی کره چشم که عمل فعال آنها برای تطابق لازم است، میروند.
The afferent and efferent limbs of accommodation reflex are depicted in Flowchart 18.2.
عضوهای آوران و وابران رفلکس تطابق در فلوچارت 18.2 نشان داده شده است.

FLOWCHART 18.2 Accommodation reflex.
نمودار جریان ۱۸.۲ رفلکس تطابق.
N.B.
• Since all the muscles responsible for accommodation are supplied of oculomotor nerve, the oculomotor nerve is often termed nerve of accommodation.
• Accommodation reflex is bilateral.
توجه:
• از آنجایی که تمام عضلات مسئول تطابق از عصب حرکتی چشم تغذیه میشوند، عصب حرکتی چشم اغلب عصب تطابق نامیده میشود.
• رفلکس تطابق دو طرفه است.
Clinical Correlation
Since accommodation is an active process and involves a muscular effort, watching of television or reading for long period may be tiring and may produce headache due to muscular fatigue.
همبستگی بالینی
از آنجایی که تطابق یک فرآیند فعال است و شامل تلاش عضلانی میشود، تماشای تلویزیون یا مطالعه برای مدت طولانی ممکن است خستهکننده باشد و به دلیل خستگی عضلانی باعث سردرد شود.
Auditory System
The hearing is second in importance among the special senses in humans after the vision. The auditory system transmits the stimulus responsible for hearing from the hearing receptor in the internal ear (also called spiral organ or organ of Corti) to the auditory area of the cerebral cortex.
سیستم شنوایی
شنوایی پس از بینایی، دومین حس مهم در بین حواس ویژه انسان است. سیستم شنوایی، محرک مسئول شنوایی را از گیرنده شنوایی در گوش داخلی (که به آن اندام مارپیچی یا اندام کورتی نیز گفته میشود) به ناحیه شنوایی قشر مغز منتقل میکند.
The following structures are included in the auditory system:
• Organ of Corti (hearing receptor).
• Dorsal and ventral cochlear nuclei.
• Trapezoid body and its nuclei.
• Lateral lemniscus.
• Inferior colliculus.
• Medial geniculate body.
• Auditory radiation.
• Auditory cortex.
ساختارهای زیر در سیستم شنوایی گنجانده شدهاند:
• اندام کورتی (گیرنده شنوایی).
• هستههای حلزونی پشتی و شکمی.
• جسم ذوزنقهای و هستههای آن.
• لبه خارجی.
• کالیکولوس تحتانی.
• جسم زانویی داخلی.
• تشعشع شنوایی.
• قشر شنوایی.
Organ of Corti (Hearing Receptor) (Fig. 18.11)
Organ of Corti is located in the cochlear duct. The cells inside the cochlear duct are highly modified to form a structure called organ of Corti. The organ of Corti consists of:
اندام کورتی (گیرنده شنوایی) (شکل ۱۸.۱۱)
اندام کورتی در مجرای حلزونی گوش قرار دارد. سلولهای داخل مجرای حلزونی به شدت تغییر شکل یافتهاند تا ساختاری به نام اندام کورتی را تشکیل دهند. اندام کورتی شامل موارد زیر است:

FIG. 18.11 Spiral organ (organ of Corti).
شکل ۱۸.۱۱ اندام مارپیچی (اندام کورتی).
• Two rod (pillar) cells called rods of Corti stand on the basilar membrane and project into the scala media. The triangular interval between the two rods the tunnel of Corti is filled with perilymph.
• دو سلول میلهای (ستونی) به نام میلههای کورتی روی غشای پایه قرار دارند و به داخل اسکالا مدیا بیرون زدهاند. فاصله مثلثی بین دو میله، تونل کورتی، پر از پری لنف است.
• Specialized sensory cells called hair cells, are arranged in a single row internal to the inner rod, and in 3 or 4 rows external to the outer rod. From the upper surface of the hair cells project tiny cilia called stereocilia. The tips of the cilia are embedded within the gelatinous shelf called tectorial membrane, which is attached at one end to the limbus and at the other end to the Hensen’s cells. The afferent fibres of cochlear nerve form synaptic contact with the plasma membrane of the hair cells.
• سلولهای حسی تخصصی به نام سلولهای مویی، در یک ردیف داخلی به سمت میله داخلی و در ۳ یا ۴ ردیف خارجی به سمت میله خارجی قرار گرفتهاند. از سطح بالایی سلولهای مویی، مژکهای کوچکی به نام استریوسیلیا بیرون زدهاند. نوک مژکها در داخل قفسه ژلاتینی به نام غشای تکتوریال قرار گرفتهاند که از یک طرف به لیمبوس و از طرف دیگر به سلولهای هنسن متصل است. فیبرهای آوران عصب حلزونی با غشای پلاسمایی سلولهای مویی تماس سیناپسی برقرار میکنند.
• The phalangeal cells (supporting cells of Deiter) form cap-like investments around the bases of hair cells. The supporting Hensen’s cells are elongated columnar cells and lie outside the external row of outer hair cells, succeeded more laterally by cubical cells of Claudius.
• سلولهای فالانژیال (سلولهای پشتیبان دیتر) در اطراف پایه سلولهای مویی، پوششهای کلاه مانندی تشکیل میدهند. سلولهای هنسنِ پشتیبان، سلولهای ستونی کشیدهای هستند و در خارج از ردیف خارجی سلولهای مویی خارجی قرار دارند و سلولهای مکعبی کلودیوس به صورت جانبیتر جایگزین آنها شدهاند.
• The reticular lamina is a tough membrane supported by rods of Corti and presents tiny holes for the passage of stereocilia.
• لایه مشبک، غشایی سفت است که توسط میلههای کورتی پشتیبانی میشود و سوراخهای کوچکی برای عبور استریوسیلیا ارائه میدهد.
The spaces other than the tunnel of Corti in the organ of Corti are filled with the corticolymph.
فضاهای دیگر به جز تونل کورتی در اندام کورتی با کورتیکولنف پر شدهاند.
Steps Involved In Hearing
مراحل دخیل در شنوایی
• The sound waves are collected by the auricle and are conducted through the external auditory meatus to the tympanic membrane to make it vibrate.
• امواج صوتی توسط لاله گوش جمعآوری شده و از طریق مجرای شنوایی خارجی به پرده صماخ هدایت میشوند تا آن را به ارتعاش درآورند.
• The vibrating tympanic membrane causes 3 ear ossicles in the middle ear (malleus, incus and stapes) to vibrate.
• ارتعاش پرده صماخ باعث ارتعاش ۳ استخوانچه گوش در گوش میانی (چکشی، سندانی و رکابی) میشود.
• The vibrations of the footplate of stapes produces vibrations in the perilymph of the scala vestibuli (here sound waves are converted into fluid waves).
• ارتعاشات صفحه پایینی استخوان رکابی، ارتعاشاتی را در پریلنف نردبان دهلیزی ایجاد میکند (در اینجا امواج صوتی به امواج سیال تبدیل میشوند).
• The vibrations of the perilymph cause simultaneous vibrations of the vestibular membrane and the endolymph in the cochlear duct which causes basilar membrane to vibrate.
• ارتعاشات پریلنف باعث ارتعاش همزمان غشای دهلیزی و اندولنف در مجرای حلزونی میشود که باعث ارتعاش غشای پایه میشود.
• As the basilar membrane vibrates the hair cells of the organ of Corti attached to it move. Consequently the stereocilia of hair cells embedded in the tectorial membrane become bent.
• با ارتعاش غشای پایه، سلولهای مویی اندام کورتی متصل به آن حرکت میکنند. در نتیجه، استریوسیلیاهای سلولهای مویی تعبیه شده در غشای تکتوریال خم میشوند.
• Bending of the stereocilia causes depolarization of the hair cells.
• خم شدن استریوسیلیاها باعث دپلاریزاسیون سلولهای مویی میشود.
• The hair cells induce action potentials in the neurons of cochlear nerve.
• سلولهای مویی، پتانسیلهای عمل را در نورونهای عصب حلزونی القا میکنند.
• The action potential generated in the cochlear nerve neurons is conducted to the cerebral cortex via auditory pathway, where it is interpreted and perceived as sound.
• پتانسیل عمل تولید شده در نورونهای عصب حلزونی از طریق مسیر شنوایی به قشر مغز منتقل میشود، جایی که به صورت صدا تفسیر و درک میشود.
Auditory Neural Pathways (fig. 18.12)
The cell bodies of first order sensory neurons of the auditory pathways lie in the spiral (cochlear) ganglion which is located within the cochlear modiolus. These neurons are bipolar. Their peripheral processes reach the organ of Corti which is the end organ (receptor organ) for hearing, while the central processes constitute the cochlear nerve, which enters the brainstem at the junction of medulla and pons. The fibres bifurcate, one branch ending in the dorsal cochlear nucleus and the other in the ventral cochlear nucleus.
مسیرهای عصبی شنوایی (شکل ۱۸.۱۲)
جسم سلولی نورونهای حسی درجه یک مسیرهای شنوایی در گانگلیون مارپیچی (حلزونی) قرار دارند که در مدولوس حلزونی قرار دارد. این نورونها دو قطبی هستند. زوائد محیطی آنها به اندام کورتی میرسد که اندام انتهایی (اندام گیرنده) برای شنوایی است، در حالی که زوائد مرکزی عصب حلزونی را تشکیل میدهند که در محل اتصال بصل النخاع و پل مغزی وارد ساقه مغز میشود. این الیاف دو شاخه میشوند، یک شاخه به هسته حلزونی پشتی و دیگری به هسته حلزونی شکمی ختم میشود.
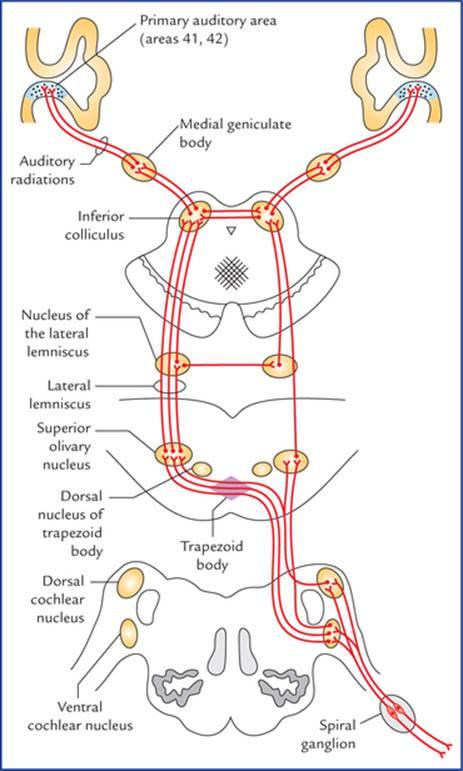
FIG. 18.12 The auditory pathways. The connections are not shown with dorsal nucleus of trapezoid body for the sake of clarity of diagram.
شکل ۱۸.۱۲ مسیرهای شنوایی. برای وضوح نمودار، اتصالات با هسته پشتی جسم ذوزنقهای نشان داده نشدهاند.
The second order sensory neurons arise from the cells of cochlear nuclei, and end in the dorsal nucleus of corpus trapezoideum and superior olivary nucleus of the same side as well as of the opposite side.
نورونهای حسی رده دوم از سلولهای هسته حلزونی منشأ میگیرند و به هسته پشتی جسم ذوزنقهای و هسته زیتونی فوقانی در همان طرف و همچنین در طرف مقابل ختم میشوند.
The third order sensory neurons arise from the dorsal nucleus of the trapezoid body and superior olivary nucleus of both sides and ascend as the lateral lemnisci. The lateral lemniscus is the principal ascending auditory tract. It ascends from the region of superior olivary nucleus, through the lateral part of pontine tegmentum and close to the surface of brainstem. A few of its fibres relay in the nucleus of inferior colliculus (centre for auditory reflexes) for reflex activity, which sends new sets of axons to the medial geniculate body through the brachium of inferior colliculus. However, most of the fibres of lateral lemniscus pass directly to the medial geniculate body without being relayed in the inferior colliculus. The medial geniculate bodies are the final relay stations of the hearing pathway. The fibres arising from medial geniculate bodies (fourth order sensory neurons) form the auditory radiations which run through the sublentiform part of the internal capsule to project into the anterior transverse temporal gyrus (or Heschl’s gyrus) located on the upper surface of superior temporal gyrus partly buried in the lateral sulcus. The Heschl’s gyrus contains primary auditory cortex (areas 41 and 42 of Brodmann).
نورونهای حسی رده سوم از هسته پشتی جسم ذوزنقهای و هسته زیتونی فوقانی در هر دو طرف منشأ میگیرند و به صورت لمنیسکوس جانبی بالا میروند. لمنیسکوس جانبی، مجرای شنوایی صعودی اصلی است. از ناحیه هسته زیتونی فوقانی، از طریق قسمت جانبی تگمنتوم پل مغزی و نزدیک به سطح ساقه مغز بالا میرود. تعدادی از فیبرهای آن برای فعالیت رفلکسی در هسته کولیکولوس تحتانی (مرکز رفلکسهای شنوایی) رله میشوند، که مجموعههای جدیدی از آکسونها را از طریق بازوی کولیکولوس تحتانی به جسم زانویی داخلی میفرستد. با این حال، بیشتر فیبرهای لمنیسکوس جانبی مستقیماً به جسم زانویی داخلی بدون رله شدن در کولیکولوس تحتانی منتقل میشوند. اجسام زانویی داخلی ایستگاههای رله نهایی مسیر شنوایی هستند. فیبرهای ناشی از اجسام زانویی داخلی (نورونهای حسی مرتبه چهارم) تشعشعات شنوایی را تشکیل میدهند که از قسمت زیر عدسی کپسول داخلی عبور میکنند تا به شکنج گیجگاهی عرضی قدامی (یا شکنج هشل) که در سطح بالایی شکنج گیجگاهی فوقانی قرار دارد و تا حدی در شیار جانبی دفن شده است، برسند. شکنج هشل شامل قشر شنوایی اولیه (نواحی ۴۱ و ۴۲ برودمن) است.
N.B. There is a bilateral cortical representation from each organ of Corti provided by distribution of fibres of cochlear nuclei, to the lateral lemnisci of both sides and also interneurons establishing cross-connections between nuclei of lateral lemnisci and between the inferior colliculi.
توجه: یک نمایش قشری دو طرفه از هر اندام کورتی وجود دارد که با توزیع فیبرهای هستههای حلزونی، به لمنیسهای جانبی هر دو طرف و همچنین نورونهای رابط که اتصالات متقاطع بین هستههای لمنیسهای جانبی و بین کولیکولهای تحتانی برقرار میکنند، فراهم میشود.
Points to note
نکات قابل توجه
• The trapezoid body is a conspicuous mass of fibres in the midline of the tegmentum of pons formed, as a result from decussation of efferent fibres of the cochlear nuclei of two sides. It presents a shape of trapezium in cross-section, hence the name trapezoid body. It has some scattered small nuclear masses within its ventral fibres and a large dorsal nucleus which lies lateral to the trapezoid body. The superior olivary nucleus lies lateral to the dorsal trapezoid nucleus.
• جسم ذوزنقهای، تودهای آشکار از فیبرها در خط میانی تگمنتوم پل مغزی است که در نتیجه تقاطع فیبرهای وابران هستههای حلزونی دو طرف تشکیل شده است. این جسم در مقطع عرضی شکلی شبیه ذوزنقه دارد، از این رو جسم ذوزنقهای نامیده میشود. این جسم دارای تودههای هستهای کوچک پراکنده در فیبرهای شکمی خود و یک هسته پشتی بزرگ است که در سمت خارجی جسم ذوزنقهای قرار دارد. هسته زیتونی فوقانی در سمت خارجی هسته ذوزنقه پشتی قرار دارد.
• The nucleus of lateral lemniscus is a small aggregation of nerve cells among the fibres of lateral lemniscus. They receive bilateral afferent connections from both cochlear nuclei and their efferent join the lateral lemniscus.
• هسته لمنیسکوس جانبی، تجمع کوچکی از سلولهای عصبی در میان فیبرهای لمنیسکوس جانبی است. آنها ارتباطات آوران دو طرفه را از هر دو هسته حلزونی دریافت میکنند و وابران آنها به لمنیسکوس جانبی میپیوندد.
Clinical Correlation
Each ear is represented bilaterally in the auditory pathway from the medulla upwards and projects about equally into the two cerebral hemispheres. Consequently the unilateral lesions of auditory cortex produce only a slight effect on the auditory acuity (i.e. sharpness of hearing) which is difficult to detect by audiometry testing.
همبستگی بالینی
هر گوش به صورت دو طرفه در مسیر شنوایی از بصل النخاع به بالا نمایش داده میشود و تقریباً به طور مساوی به دو نیمکره مغزی امتداد مییابد. در نتیجه، ضایعات یک طرفه قشر شنوایی تنها تأثیر کمی بر حدت شنوایی (یعنی تیزی شنوایی) میگذارد که تشخیص آن با آزمایش شنوایی سنجی دشوار است.
Vestibular System
The vestibular pathways transmit impulses from vestibular receptors in the internal ear to the vestibular nuclei in the brainstem, which in turn project to widespread area of the central nervous system. It is responsible for maintenance of body-balance.
سیستم دهلیزی
مسیرهای دهلیزی، تکانهها را از گیرندههای دهلیزی در گوش داخلی به هستههای دهلیزی در ساقه مغز منتقل میکنند که به نوبه خود به ناحیه وسیعی از سیستم عصبی مرکزی امتداد مییابند. این سیستم مسئول حفظ تعادل بدن است.
Vestibular Receptors
The organs of balance can be divided structurally and functionally into two parts: (a) the static labyrinth consisting of utricle and saccule of the vestibule, and (b) the kinetic labyrinth consisting of three semicircular ducts. The static labyrinth detects the position of head with respect to gravity, although it also responds to linear acceleration or deceleration, viz. a car that slows down or speeds. The kinetic labyrinthis involved in evaluating the movements of the head and has a special role in coordination of eye movements with movements of the head.
گیرندههای دهلیزی
ارگانهای تعادل را میتوان از نظر ساختاری و عملکردی به دو بخش تقسیم کرد: (الف) لابیرنت ایستا که از اتریکول و ساکول دهلیز تشکیل شده است، و (ب) لابیرنت جنبشی که از سه مجرای نیمدایرهای تشکیل شده است. لابیرنت ایستا موقعیت سر را نسبت به جاذبه تشخیص میدهد، اگرچه به شتاب یا کاهش سرعت خطی نیز پاسخ میدهد، مثلاً خودرویی که سرعتش کم میشود یا افزایش مییابد. لابیرنت جنبشی در ارزیابی حرکات سر نقش دارد و نقش ویژهای در هماهنگی حرکات چشم با حرکات سر دارد.
The receptors for static balance are maculae in the wall of utricle and saccule. The macula is a specialized patch of epithelium in the wall of utricle and saccule (Fig. 18.13). It consists of columnar supporting cells and hair cells. The hair of these cells which consists of numerous microvilli, called stereocilia, and cilium (kinocilium) that arises from centriole, are embedded into a gelatinous mass containing irregularly shaped concretions (otoliths) composed of protein and calcium carbonate. The gelatinous mass moves in response to gravity, bending the hair cells and initiating the action potentials in the associated neurons (Fig. 18.14).
گیرندههای تعادل ایستا، ماکولاها در دیواره اتریکول و ساکول هستند. ماکولا یک تکه اپیتلیوم تخصصی در دیواره اتریکول و ساکول است (شکل 18.13). این لابیرنت از سلولهای پشتیبان ستونی و سلولهای مویی تشکیل شده است. موهای این سلولها که از میکروویلیهای متعدد به نام استریوسیلیا و مژک (کینوسیلیوم) که از سانتریول منشأ میگیرند، تشکیل شدهاند، در یک توده ژلاتینی حاوی سنگهای نامنظم (اتولیتها) متشکل از پروتئین و کربنات کلسیم قرار گرفتهاند. توده ژلاتینی در پاسخ به جاذبه حرکت میکند، سلولهای مویی را خم میکند و پتانسیلهای عمل را در نورونهای مرتبط آغاز میکند (شکل 18.14).

FIG. 18.13 Anterolateral view of right membranous labyrinth showing location of receptors for static balance (maculae in the wall of utricle and saccule) and kinetic balance (cristae ampul-laris in the ampullae of semicircular canals).
شکل ۱۸.۱۳ نمای قدامی-جانبی لابیرنت غشایی راست که محل گیرندههای تعادل ایستا (ماکولا در دیواره اتریکول و ساکول) و تعادل جنبشی (کریستا آمپولاریس در آمپولهای مجاری نیمدایرهای) را نشان میدهد.
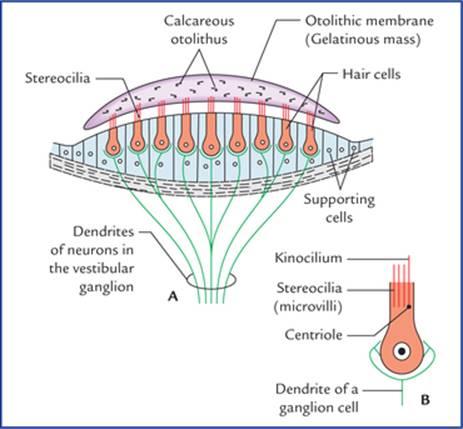
FIG. 18.14 (A) Structure of the macula. (B) An enlarged hair cell showing kinocilium.
شکل ۱۸.۱۴ (الف) ساختار ماکولا. (ب) یک سلول مویی بزرگ شده که کینوسیلیوم را نشان میدهد.
The base of each semicircular canal is expanded into an ampulla. Within each ampulla the epithelium is specialized to form crista ampullaris, the receptor for kinetic balance (Fig. 18.15). It is structurally and functionally very similar to that of macula. Each crista consists of a ridge or crest of epithelium with curved gelatinous mass, the cupula suspended over the crest.
پایه هر کانال نیمدایرهای به یک آمپولا گسترش یافته است. در داخل هر آمپولا، اپیتلیوم برای تشکیل کریستا آمپولاریس، گیرنده تعادل جنبشی، تخصص یافته است (شکل ۱۸.۱۵). از نظر ساختاری و عملکردی بسیار شبیه به ماکولا است. هر کریستا از یک برآمدگی یا تاج اپیتلیوم با توده ژلاتینی خمیده تشکیل شده است، کاپولا روی تاج آویزان است.

FIG. 18.15 Structure of crista ampullaris.
شکل ۱۸.۱۵ ساختار کریستا آمپولاریس.
The hair-like processes of the crista hair cells, similar to those in the maculae are embedded in a gelatinous mass that forms cupula. The cupula does not contain otoliths and therefore cannot respond to the gravitational pull. Instead, the cupula is like a float that is displaced by fluid movements within the semicircular canals. The endolymph movement within each semicircular canal moves the cupula, bends the hairs and initiates the action potentials in the associated neurons of vestibular nerve (Fig. 18.15).
زوائد مو مانند سلولهای مویی کریستا، مشابه زوائد موجود در ماکولا، در یک توده ژلاتینی قرار گرفتهاند که کوپولا را تشکیل میدهند. کوپولا حاوی اتولیت نیست و بنابراین نمیتواند به کشش گرانشی پاسخ دهد. در عوض، کوپولا مانند یک شناور است که توسط حرکات سیال در داخل مجاری نیمدایرهای جابجا میشود. حرکت اندولنف در هر مجرای نیمدایرهای، کوپولا را حرکت میدهد، موها را خم میکند و پتانسیلهای عمل را در نورونهای مرتبط با عصب دهلیزی آغاز میکند (شکل ۱۸.۱۵).
The three semicircular ducts of kinetic labyrinth are placed nearly at right angles to each other, one lying nearly in the transverse plane, one in the coronal plane, and one in the sagittal plane. The arrangement of semicircular canals enables a person to detect movement in any direction.
سه مجرای نیمدایرهای لابیرنت جنبشی تقریباً در زاویه قائمه نسبت به یکدیگر قرار گرفتهاند، یکی تقریباً در صفحه عرضی، یکی در صفحه تاجی و دیگری در صفحه ساژیتال قرار دارد. چیدمان مجاری نیمدایرهای به فرد این امکان را میدهد که حرکت را در هر جهتی تشخیص دهد.
Neural Pathways for Balance (Figs 18.16 and 18.17)
The cell bodies of first order (bipolar) sensory neurons form the vestibular ganglion located at the bottom (fundus) of the internal auditory meatus. The peripheral processes of these cells end on the hair cells of the maculae of the utricle and saccule; and on the hair cells of the cristae of semicircular ducts of the internal ear. The central processes of the bipolar cells of the vestibular ganglion form the vestibular nerve which run in the internal auditory meatus, enters the cranial cavity and traverse the subarachnoid space at cerebellopontine angle to enter the pontomedullary junction superomedial to the cochlear nerve. These fibres relay in the vestibular nuclei (for details see Chapter 8) and flocculonodular lobe of the cerebellum.
مسیرهای عصبی برای تعادل (شکلهای ۱۸.۱۶ و ۱۸.۱۷)
جسم سلولی نورونهای حسی درجه یک (دوقطبی) گانگلیون دهلیزی را تشکیل میدهند که در پایین (فوندوس) مجرای شنوایی داخلی قرار دارد. زوائد محیطی این سلولها به سلولهای مویی ماکولاهای اتریکول و ساکول و به سلولهای مویی کریستاهای مجاری نیمدایرهای گوش داخلی ختم میشوند. زوائد مرکزی سلولهای دوقطبی گانگلیون دهلیزی، عصب دهلیزی را تشکیل میدهند که در مجرای شنوایی داخلی امتداد یافته، وارد حفره جمجمه شده و از فضای زیر عنکبوتیه در زاویه پل مخچهای عبور میکند تا به محل اتصال پل مغزی به نخاع در قسمت فوقانی میانی عصب حلزونی وارد شود. این فیبرها در هستههای دهلیزی (برای جزئیات بیشتر به فصل ۸ مراجعه کنید) و لوب فلوکولوندولار مخچه رله میشوند.

FIG. 18.16 Neural pathways for balance-peripheral connections.
شکل ۱۸.۱۶ مسیرهای عصبی برای ارتباطات تعادل-محیطی.
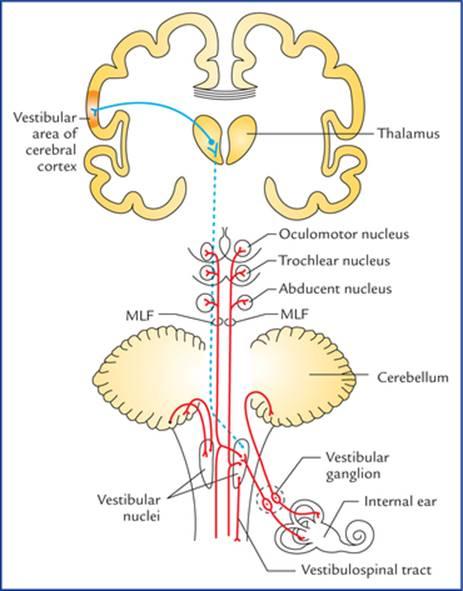
FIG. 18.17 Neural pathways for balance-central connections. (MLF = Medial longitudinal fasciculus).
شکل ۱۸.۱۷ مسیرهای عصبی برای ارتباطات تعادل-مرکزی. (MLF = دسته طولی داخلی).
The second order sensory neurons of vestibular nuclei project: (a) to the flocculonodular lobe of the cerebellum (archicerebellum) through inferior cerebellar peduncle (vestibulocerebellar tract), (b) to the motor nuclei of IIIrd, IVth and VIth cranial nerves through medial longitudinal fasciculus (MLF), and (c) to the anterior horn cells of the spinal cord (vestibulospinal tract). The vestibular nuclei also project to the posteroventral nucleus of the thalamus.
نورونهای حسی رده دوم هستههای دهلیزی: (الف) از طریق ساقه مخچه تحتانی (راه دهلیزی-مخچهای)، (ب) از طریق دسته طولی داخلی (MLF) به هستههای حرکتی اعصاب جمجمهای ردههای سوم، چهارم و ششم، و (ج) به سلولهای شاخ قدامی نخاع (راه دهلیزی-نخاعی) امتداد مییابند. هستههای دهلیزی همچنین به هسته خلفی-شکمی تالاموس امتداد مییابند.
The third order sensory neurons from thalamus project to the vestibular area of the cerebral cortex in the postcentral gyrus.
نورونهای حسی رده سوم از تالاموس به ناحیه دهلیزی قشر مغز در شکنج پس مرکزی امتداد مییابند.
The vestibular connections contribute to the coordination of muscle contraction in maintenance of upright posture.
اتصالات دهلیزی در هماهنگی انقباض عضلات در حفظ وضعیت ایستاده نقش دارند.
Through connections of the medial longitudinal fasciculus of the same and opposite side, the vestibular system controls some vestibuloocular and postural reflexes, e.g. conjugate eye movements and movements of the neck and trunk in response to vestibular stimulation.
سیستم دهلیزی از طریق اتصالات فاسیکول طولی داخلی در همان طرف و طرف مقابل، برخی از رفلکسهای دهلیزی-چشمی و وضعیتی، مانند حرکات همزمان چشم و حرکات گردن و تنه در پاسخ به تحریک دهلیزی را کنترل میکند.
The vestibulothalamocortical connections give a sense of balance and their dysfunction has been associated with the symptoms of motion sickness. The motion sickness presents as nausea, vomiting, weakness and incapacitation caused by the stimulation of semicircular canals during motion such as in swing or amusement park ride.
اتصالات دهلیزی-تالاموکورتیکال حس تعادل را ایجاد میکنند و اختلال عملکرد آنها با علائم بیماری حرکت مرتبط است. بیماری حرکت به صورت حالت تهوع، استفراغ، ضعف و ناتوانی ناشی از تحریک مجاری نیمدایرهای در حین حرکت مانند تاب یا وسایل بازی شهربازی بروز میکند.
Clinical Correlation
The balance of the body is a complex process, dependent on inputs to the vestibular nuclei from vestibular receptors in the internal ear, proprioceptive receptors of the whole body and visual receptors in the eye. The derangement of any of the above components may give rise to clinical manifestations of trunkal ataxia and staggering gait, nystagmus (see Chapter 8), and vertigo.
همبستگی بالینی
تعادل بدن یک فرآیند پیچیده است که به ورودیهای هستههای دهلیزی از گیرندههای دهلیزی در گوش داخلی، گیرندههای حس عمقی کل بدن و گیرندههای بینایی در چشم بستگی دارد. اختلال در هر یک از اجزای فوق ممکن است منجر به تظاهرات بالینی آتاکسی تنه و تلوتلو خوردن در راه رفتن، نیستاگموس (به فصل 8 مراجعه کنید) و سرگیجه شود.
N.B. The vertigo is a feeling of the rotation, either of the patient himself or his environment.
توجه: سرگیجه احساس چرخش، چه در خود بیمار و چه در محیط اطرافش، است.
Taste
The taste pathways transmit sense of taste from taste buds located in the mucous membrane of tongue, epiglottis, soft palate and pharynx to the sensory cortex of the brain.
چشایی
مسیرهای چشایی، حس چشایی را از جوانههای چشایی واقع در غشای مخاطی زبان، اپیگلوت، کام نرم و حلق به قشر حسی مغز منتقل میکنند.
Taste Buds (Fig. 18.18)
The taste buds are the organs of taste. There are about 10,000 taste buds in human. Taste buds are oval (egg-shaped) clusters of cells in the epithelial lining of the tongue, palate and pharynx. The cells within the taste buds are of two types: (a) gustatory cells or receptor cells, and (b) supporting or sustentacular cells.
جوانههای چشایی (شکل 18.18)
جوانههای چشایی اندامهای چشایی هستند. حدود 10000 جوانه چشایی در انسان وجود دارد. جوانههای چشایی، خوشههای بیضی شکل (تخممرغی شکل) از سلولها در پوشش اپیتلیال زبان، کام و حلق هستند. سلولهای درون جوانههای چشایی دو نوع هستند: (الف) سلولهای چشایی یا سلولهای گیرنده، و (ب) سلولهای پشتیبان یا نگهدارنده.
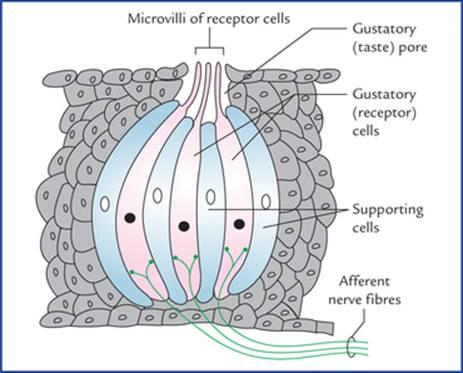
FIG. 18.18 Structure of a taste bud (taste receptor). The gustatory (taste receptor) cells contain synaptic vesicles and are innervated by afferent nerve fibres. The supporting cells are not innervated.
شکل ۱۸.۱۸ ساختار جوانه چشایی (گیرنده چشایی). سلولهای چشایی (گیرنده چشایی) حاوی وزیکولهای سیناپسی هستند و توسط فیبرهای عصبی آوران عصبدهی میشوند. سلولهای پشتیبان عصبدهی ندارند.
The gustatory cells present microvilli, which project on to the surface through the gustatory pores and are exposed to the fluid in the oral cavity.
سلولهای چشایی دارای میکروویلیهایی هستند که از طریق منافذ چشایی به سطح میآیند و در معرض مایع موجود در حفره دهان قرار میگیرند.
The taste producing substances first get dissolved in the oral fluid and then bind with the receptors located on the microvilli of the gustatory cells. The microvilli react by depolarizing the gustatory cells, which lead to the generation of action potentials in the afferent nerve endings.
مواد تولیدکننده طعم ابتدا در مایع دهان حل میشوند و سپس به گیرندههای واقع در میکروویلیهای سلولهای چشایی متصل میشوند. میکروویلیها با دپلاریزه کردن سلولهای چشایی واکنش نشان میدهند که منجر به تولید پتانسیلهای عمل در انتهای اعصاب آوران میشود.
Neural Pathways for Taste (Fig. 18.19)
The cell bodies of first order sensory neurons carrying taste sensations from taste buds are located in the geniculate ganglion, superior petrosal ganglion and inferior ganglion (ganglion nodosum), of facial, glossopharyngeal and vagus nerves respectively.
مسیرهای عصبی چشایی (شکل ۱۸.۱۹)
جسم سلولی نورونهای حسی درجه یک که حس چشایی را از جوانههای چشایی منتقل میکنند، به ترتیب در گانگلیون زانویی، گانگلیون پتروزال فوقانی و گانگلیون تحتانی (گانگلیون ندوزوم) اعصاب صورت، زبانزبانی-حلقی و واگ قرار دارند.
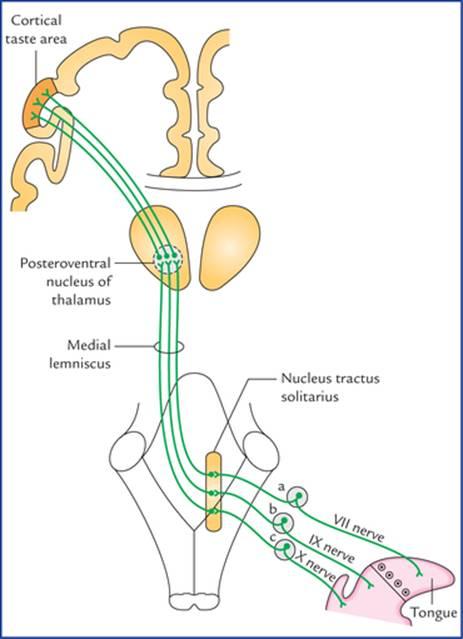
FIG. 18.19 Taste pathways: a = geniculate ganglion of facial nerve, b = superior ganglion of glossopharyngeal nerve, c = inferior ganglion (ganglion nodosum) of vagus nerve. The taste area is located in the inferior part of the postcentral gyrus (primary sensory cortex). According to Barr and Kiemann the taste (gustatory) area is located in the inferior part of the parietal lobe, posterior to the general sensory area for the mouth in the post-central gyrus.
شکل ۱۸.۱۹ مسیرهای چشایی: الف = گانگلیون زانویی عصب صورتی، ب = گانگلیون بالایی عصب زبانی-حلقی، ج = گانگلیون پایینی (گانگلیون گرهای) عصب واگ. ناحیه چشایی در قسمت تحتانی شکنج پس مرکزی (قشر حسی اولیه) قرار دارد. طبق گفته بار و کیمن، ناحیه چشایی (چشایی) در قسمت تحتانی لوب آهیانه، خلفی به ناحیه حسی عمومی دهان در شکنج پس مرکزی واقع شده است.
• Taste fibres from anterior two-third of the tongue excluding vallate papillae run in the facial nerve (from tongue successively through lingual, chorda tympani and facial nerves).
• فیبرهای چشایی از دو سوم قدامی زبان به استثنای پاپیلاهای والت، در عصب صورتی (از زبان به طور متوالی از طریق اعصاب زبانی، طناب صماخ و صورت) عبور میکنند.
• The taste fibres from posterior one-third of the tongue including vallate papillae travel along the glossopharyngeal nerve.
• فیبرهای چشایی از یک سوم خلفی زبان، شامل پاپیلاهای والت، در امتداد عصب زبانی-حلقی حرکت میکنند.
• Taste fibres from posterior most part of tongue, epiglottis and pharynx travel along the vagus nerve (successively through its internal laryngeal, and superior laryngeal branches).
• رشتههای چشایی از قسمت خلفی زبان، اپیگلوت و حلق در امتداد عصب واگ (به ترتیب از طریق شاخههای حنجرهای داخلی و حنجرهای فوقانی آن) حرکت میکنند.
The peripheral processes of these cells connect with the taste buds, while central processes end in the nucleus tractus solitarius (NTS).
زوائد محیطی این سلولها به جوانههای چشایی متصل میشوند، در حالی که زوائد مرکزی به هسته منفرد (NTS) ختم میشوند.
The second order sensory neurons arise from nucleus tractus solitarius, cross the midline, turn upwards to join the medial lemniscus and terminate in the posteroventral nucleus of the thalamus.
نورونهای حسی رده دوم از هسته تک رشتهای (nucleus tractus solitarius) منشأ میگیرند، از خط میانی عبور میکنند، به سمت بالا میچرخند تا به نوار میانی بپیوندند و در هسته خلفی-شکمی تالاموس خاتمه مییابند.
The third order sensory neurons arise from posteroventral nucleus of thalamus and project in the inferior part of the postcentral gyrus of the parietal lobe of the brain, the taste area of the cerebral cortex.
نورونهای حسی رده سوم از هسته خلفی-شکمی تالاموس منشأ میگیرند و در قسمت تحتانی شکنج پس مرکزی لوب آهیانه مغز، ناحیه چشایی قشر مغز، امتداد مییابند.
N.B. All the afferents from tongue, carrying both taste and general sensibility (pain, touch and temperature), travel along the same pathway to the thalamus and sensory cortex.
توجه: تمام آورانهای زبان، که هم حس چشایی و هم حس عمومی (درد، لامسه و دما) را حمل میکنند، در امتداد همان مسیر به تالاموس و قشر حسی میروند.
Clinical Correlation
همبستگی بالینی
• There are four primary taste sensations, viz. bitter, sour, sweet, and salt. The primary tastes are sensed better in certain areas. The bitter is sensed at the back of the tongue, sour at the sides, the sweet at the tip and the salt on the dorsum.
• چهار حس چشایی اولیه وجود دارد، یعنی تلخ، ترش، شیرین و شور. مزههای اولیه در نواحی خاصی بهتر حس میشوند. تلخی در پشت زبان، ترشی در کنارهها، شیرینی در نوک و شوری در پشت زبان حس میشوند.
• The thresholds vary for the four primary tastes. Sensitivity for bitter substances is the highest; sensitivities for sweet and salty tastes are the lowest. Sugars, some carbohydrates, and some proteins produce sweet tastes; acids produce sour tastes; metal ions tend to produce salty tastes; and alkaloids (bases) produce bitter tastes. Many alkaloids are poisonous, thus the high sensitivity for bitter tastes may be protective.
• آستانههای چهار مزه اصلی متفاوت است. حساسیت به مواد تلخ بالاترین و حساسیت به مزههای شیرین و شور کمترین است. قندها، برخی کربوهیدراتها و برخی پروتئینها مزه شیرین ایجاد میکنند؛ اسیدها مزه ترش ایجاد میکنند؛ یونهای فلزی تمایل به ایجاد مزه شور دارند؛ و آلکالوئیدها (بازها) مزه تلخ ایجاد میکنند. بسیاری از آلکالوئیدها سمی هستند، بنابراین حساسیت بالا به مزههای تلخ ممکن است محافظتی باشد.
• Electrical stimulation of postcentral gyrus produces hallucinations of taste and its destruction abolishes the taste and tactile sensibility of the tongue.
• تحریک الکتریکی شکنج پس مرکزی باعث توهم چشایی میشود و تخریب آن، مزه و حساسیت لمسی زبان را از بین میبرد.
Clinical Problems
مشکلات بالینی
1. Explain the cause of blood-stained watery discharge from the nostril and loss of smell in fracture of anterior cranial fossa.
1. علت ترشحات آبکی خونآلود از سوراخ بینی و از دست دادن حس بویایی در شکستگی حفره جمجمهای قدامی را توضیح دهید.
2. The complete injury of optic nerve leads to permanent blindness. Mention the anatomical basis.
2. آسیب کامل عصب بینایی منجر به نابینایی دائمی میشود. اساس آناتومیک را ذکر کنید.
3. The increased intracranial pressure often produces bilateral papilloedema (Swelling of optic discs). Explain.
3. افزایش فشار داخل جمجمه اغلب باعث ادم پاپی دو طرفه (تورم دیسکهای بینایی) میشود. توضیح دهید.
4. Explain how a pituitary tumour produces a bitemporal loss of vision (‘tunnel vision’).
4. توضیح دهید که چگونه تومور هیپوفیز باعث از دست دادن بینایی دو گیجگاهی (“دید تونلی”) میشود.
5. When light is shown to one eye, the pupils of both the eyes constrict. Why?
5. هنگامی که نور به یک چشم تابیده میشود، مردمکهای هر دو چشم تنگ میشوند. چرا؟
6. Mention the anatomical basis of Argyll Robertson pupil in neurosyphilis.
6. اساس آناتومیک مردمک آرگیل رابرتسون را در نوروسیفلیس ذکر کنید.
7. Explain why a sudden brief and extremely loud sound such as due to bomb explosion can result in deafness?
7. توضیح دهید که چرا یک صدای ناگهانی کوتاه و بسیار بلند مانند صدای انفجار بمب میتواند منجر به ناشنوایی شود؟
Clinical Problem Solving
حل مسئله بالینی
1. The blood-stained watery discharge through the nostril occurs due to haemorrhage and leakage of cerebrospinal fluid following fracture of cribriform plate of the ethmoid bone. The loss of smell (anosmia) occurs due to lesion of the olfactory nerves, bulbs and tracts.
1. ترشحات آبکی آغشته به خون از طریق سوراخ بینی به دلیل خونریزی و نشت مایع مغزی نخاعی پس از شکستگی صفحه مشبک استخوان اتموئید رخ میدهد. از دست دادن حس بویایی (آنوسمی) به دلیل آسیب به اعصاب، پیازها و مجاری بویایی رخ میدهد.
2. This is because the optic nerve cannot regenerate as it is not a true peripheral nerve. Remember the optic nerve and retina are extensions of the brain, and the nerve itself represents the white matter. Its fibres do not possess neurilemmal sheath and the oligodendrocytes existing as its glial cells form the myelin sheath around the nerve fibres. Due to absence of neurilemmal sheath the optic nerve is incapable of regeneration after the injury.
2. این به این دلیل است که عصب بینایی نمیتواند بازسازی شود زیرا یک عصب محیطی واقعی نیست. به یاد داشته باشید که عصب بینایی و شبکیه امتداد مغز هستند و خود عصب نمایانگر ماده سفید است. الیاف آن دارای غلاف نوریلمی نیستند و الیگودندروسیتهای موجود به عنوان سلولهای گلیال آن، غلاف میلین را در اطراف الیاف عصبی تشکیل میدهند. به دلیل عدم وجود غلاف نوریلمی، عصب بینایی پس از آسیب قادر به بازسازی نیست.
3. This is because the increased intracranial pressure is transmitted to the subarachnoid space around the optic nerve on both the sides. As a result the venous return through the central vein of the retina, which traverses this space is impeded.
3. این به این دلیل است که افزایش فشار داخل جمجمه به فضای زیر عنکبوتیه اطراف عصب بینایی در هر دو طرف منتقل میشود. در نتیجه، بازگشت وریدی از طریق ورید مرکزی شبکیه که از این فضا عبور میکند، مختل میشود.
4. The pituitary gland is located in the pituitary fossa (sella turcica) and related superiorly to the optic chiasma. The loss of vision in both temporal fields (i.e. bitemporal hemianopia) occurs due to the suprasellar extension of pituitary tumour affecting nasal fibres from the both retinae.
۴. غده هیپوفیز در حفره هیپوفیز (sella turcica) قرار دارد و در بالا به کیاسمای بینایی مرتبط است. از دست دادن بینایی در هر دو میدان گیجگاهی (یعنی همیآنوپی دو گیجگاهی) به دلیل گسترش تومور هیپوفیز به بالای زلالی که فیبرهای بینی را از هر دو شبکیه تحت تأثیر قرار میدهد، رخ میدهد.
5. This is due to direct and indirect consensual light reflexes (seepage 217).
۵. این به دلیل رفلکسهای نوری مستقیم و غیرمستقیم توافقی است (صفحه ۲۱۷).
6. The neurosyphilis produces lesions in or near the aqueduct of Sylvius involving superior colliculi which interrupts the pathway of light reflex only. In ‘Argyll Robertson pupil’ the pupil does not react to light (loss of light reflex) but does react to accommodation (no loss of accommodation reflex) because pathways for light and accommodation reflexes are different (see pages 217 and 218).
۶. نوروسیفیلیس ضایعاتی را در داخل یا نزدیکی مجرای سیلویوس ایجاد میکند که شامل کولیکولهای فوقانی میشود که فقط مسیر رفلکس نور را قطع میکند. در «مردمک آرگیل رابرتسون»، مردمک به نور واکنش نشان نمیدهد (از دست دادن رفلکس نور) اما به تطابق واکنش نشان میدهد (بدون از دست دادن رفلکس تطابق) زیرا مسیرهای رفلکسهای نوری و تطابق متفاوت هستند (به صفحات ۲۱۷ و ۲۱۸ مراجعه کنید).
7. Due to excessive displacement of hair cells of the organ of Corti (the auditory receptors).
۷. به دلیل جابجایی بیش از حد سلولهای مویی اندام کورتی (گیرندههای شنوایی).
Textbook of Clinical Neuroanatomy, 2 ed
Chapter 1. Development of the Nervous System
Chapter 2. Organization and Functions of the Nervous System
Chapter 3. Peripheral Nerves and Ganglia
Chapter 4. Receptors and Effectors
Chapter 5. Dermatomes and Muscular Activity
Chapter 6. Central Nervous System: an Overview
Chapter 7. Spinal Cord
Chapter 8. Brainstem
Chapter 9. Nuclei, Functional Components and Distribution of Cranial Nerves
Chapter 10. Cerebellum and Fourth Ventricle
Chapter 11. Diencephalon and Third Ventricle
Chapter 12. Cerebrum
Chapter 13. Basal Nuclei (Basal Ganglia)
Chapter 14. White Matter of the Cerebrum and Lateral Ventricles
Chapter 15. Blood Supply of the Brain
Chapter 16. Meninges And Cerebrospinal Fluid
Chapter 17. Somatic Motor and Sensory Pathways
Chapter 18. Special Senses and their Neural Pathways
Chapter 19. Reticular Formation and Limbic System
Chapter 20. Autonomic Nervous System