علوم اعصاب برای پزشکان بالینی؛ کنترل اجرایی؛ حالت بیداری، توجه و حافظه کاری

دعای مطالعه [ نمایش ]
بِسْمِ الله الرَّحْمنِ الرَّحیمِ
اَللّهُمَّ اَخْرِجْنى مِنْ ظُلُماتِ الْوَهْمِ
خدايا مرا بيرون آور از تاريكىهاى وهم،
وَ اَكْرِمْنى بِنُورِ الْفَهْمِ
و به نور فهم گرامى ام بدار،
اَللّهُمَّ افْتَحْ عَلَيْنا اَبْوابَ رَحْمَتِكَ
خدايا درهاى رحمتت را به روى ما بگشا،
وَانْشُرْ عَلَيْنا خَزائِنَ عُلُومِكَ بِرَحْمَتِكَ يا اَرْحَمَ الرّاحِمينَ
و خزانههاى علومت را بر ما باز كن به امید رحمتت اى مهربانترين مهربانان.
» کتاب علوم اعصاب برای پزشکان بالینی
» » فصل ۴۲: فصل کنترل اجرایی؛ قسمت اول
در حال ویرایش
» Neuroscience for Clinicians
»» chapter 42: Executive Control
» Overview
» Attention
» Working Memory
» Decision-Making, Motivation, and Allocation of Effort
» Cognitive Control Networks
» Clinical Correlations
» Key Points
| مرور کلی | 781 |
| توجه | 781 |
| حافظه کاری | 785 |
| تصمیم گیری، ایجاد انگیزه و تخصیص تلاش | 787 |
| شبکههای کنترل شناختی | 789 |
| همبستگیهای بالینی | 793 |
| نکات کلیدی | 795 |
OVERVIEW
The ability to rapidly and accurately learn and process new information in response to a changing environment, engage in goal-directed tasks, and switch between behaviors according to the context requires attention, working memory, decision-making, and cognitive control. All these processes engage large-scale networks involving different portions of the frontal cortex and interactions with posterior cortical areas in the parietal and temporal lobes. The frontal lobe consists of multiple cytoarchitectonically distinct areas with specific connections to other brain regions. (1) The lateral prefrontal cortex (PFC), medial PFC, or bitofrontal cortex (OFC), anterior cingulate cortex (ACC), and anterior midcingulate cortex (MCC) are fundamental components of separate yet interactive networks. Metaanalyses of functional neuroimaging studies have provided maps of the functional correlates of behavioral functions in these frontal regions (2, 3) (Figure 42.1). For example, lateral frontal areas are part of parietal networks involved in attention and executive functions, somatosensory networks involved in motor planning and programming, and the default mode network involved in inner thought processing. Similarly, medial frontal cortical areas have been subdi-vided into anterior, middle, and posterior regions according to their participation in networks involved in emotional/ social, cognitive, and motor functions, respectively. Thus, different areas of the frontal cortex are core regions of largescale cognitive networks involved in attention, (4-6) working memory, (7-9) decision-making (10, 11) and executive control. (12, 13) Attention selects task-relevant information. Working memory maintains the relevant information “on-line” over brief periods of time for processing, manipulation, and transformation to guide behavior. Decision-making selects a particular option among a set of alternatives taking into account their reward value, costs, and personal relevance. Motivation is a process by which incentives invigorate and guide behaviors toward goals. Control signals are required for implementation, maintenance, and updating of behavior based on feedback, particularly when it is necessary to overcome distraction or suppress habitual responses when outcomes deviate from expectations. Appropriate cognitive control requires cognitive effort, which is the ability to allocate cognitive resources to the pursuit of a goal. (14, 15) Control mechanisms allow the flexible shift of motivated behaviors between those triggered by relevant external stimuli captured by the attention networks and those driven by internal thoughts of personal relevance processed in the default and functions, these networks are located along a gradient mode network. (16) Based on microstructure, connectivity, of hierarchical cortical organization and are flexibly engaged in different cognitive tasks. (17, 18) Dysfunction of these large-scale networks is a cardinal feature of neurodegenerative dementias, particularly behavioral variant frontotemporal dementia (bvFTD) and Alzheimer disease (AD); focal lesions affecting different portions of the frontal lobe; (19) including stroke; (20) and psychiatric disorders. Mapping of cognitive and emotional networks using resting-state fMRI may have potential applications for the mapping of cognition and emotion in neurosurgical patients. (21) The aims of this chapter are to review: (1) the areas and mechanisms involved in selective attention and working memory; (2) mechanisms of decision-making, motivation, and allocation of effort; (3) the areas involved in cognitive control; and (4) examples of disorders affecting these large-scale networks.
مرور کلی
توانایی یادگیری سریع و دقیق و پردازش اطلاعات جدید در پاسخ به یک محیط در حال تغییر، درگیر شدن در وظایف هدفمند، و تغییر بین رفتارها با توجه به زمینه، نیازمند توجه، حافظه کاری، تصمیم گیری و کنترل شناختی است. همه این فرآیندها شبکههای مقیاس بزرگی را درگیر میکنند که بخشهای مختلف قشر پیشانی و برهمکنشهایی با نواحی قشر خلفی در لوبهای آهیانهای و تمپورال دارند. لوب فرونتال شامل چندین نواحی متمایز از نظر سیتو آرشیتکتونیکی با اتصالات خاص به سایر مناطق مغز است. (۱) قشر پرهفرونتال جانبی (PFC)، PFC میانی، یا قشر اربیتوفرونتال (OFC)، قشر سینگولیت قدامی (ACC) و قشر میانی قدامی (MCC) اجزای اساسی شبکههای مجزا و در عین حال تعاملی هستند. متاآنالیزهای مطالعات تصویربرداری عصبی عملکردی، نقشههایی از همبستگیهای عملکردی عملکردهای رفتاری در این نواحی پیشانی ارائه کرده است (۳، ۲) (شکل 42.1). برای مثال، نواحی فرونتال جانبی بخشی از شبکههای آهیانهای است که در توجه و عملکردهای اجرایی، شبکههای حسی پیکری درگیر در برنامهریزی و برنامهریزی حرکتی، و شبکه حالت پیشفرض درگیر در پردازش فکر درونی هستند. به طور مشابه، نواحی قشر پیشانی میانی بر اساس مشارکت در شبکههای درگیر در عملکردهای عاطفی/اجتماعی، شناختی و حرکتی به نواحی قدامی، میانی و خلفی تقسیم میشوند. بنابراین، نواحی مختلف قشر پیشانی نواحی اصلی شبکههای شناختی در مقیاس بزرگ هستند که در توجه، (۶-۴) حافظه کاری، (۹-۷) تصمیمگیری (۱۱، ۱٠) و کنترل اجرایی دخیل هستند. (۱۳، ۱۲) توجه اطلاعات مربوط به کار را انتخاب میکند. حافظه کاری اطلاعات مربوطه را به صورت آنلاین در مدت زمان کوتاهی برای پردازش، دستکاری و تغییر جهت هدایت رفتار نگهداری میکند. تصمیم گیری یک گزینه خاص را از میان مجموعه ای از گزینهها با در نظر گرفتن ارزش پاداش، هزینهها و ارتباط شخصی آنها انتخاب میکند. انگیزه فرآیندی است که از طریق آن انگیزهها باعث تقویت و هدایت رفتارها به سمت اهداف میشود. سیگنالهای کنترلی برای اجرا، نگهداری و بهروزرسانی رفتار بر اساس بازخورد، بهویژه زمانی که برای غلبه بر حواسپرتی یا سرکوب واکنشهای معمول در زمانی که نتایج از انتظارات منحرف میشود، ضروری است. کنترل شناختی مناسب مستلزم تلاش شناختی است، که توانایی تخصیص منابع شناختی برای دستیابی به یک هدف است. (۱۵، ۱۴) مکانیسمهای کنترلی امکان جابجایی انعطافپذیر رفتارهای با انگیزه را بین محرکهای بیرونی مرتبط که توسط شبکههای توجه بهدست میآیند و آنهایی که توسط افکار درونی در این شبکههای ارتباطی پیشفرض واقع شدهاند، میدهند. (۱۶) بر اساس ریزساختار، اتصال، سازمان سلسله مراتبی قشر مغز و به طور انعطافپذیری درگیر وظایف شناختی مختلف هستند. (۱۸، ۱۷) اختلال در این شبکههای مقیاس بزرگ یکی از ویژگیهای اصلی زوال عقلهای تخریبکننده عصبی، بهویژه نوع رفتاری دمانس فرونتوتمپورال (bvFT) است. ضایعات کانونی که بخشهای مختلف لوب فرونتال را تحت تأثیر قرار میدهند؛ (۱۹) شامل سکته مغزی؛ (۲۰) و اختلالات روانی. نقشهبرداری از شبکههای شناختی و عاطفی با استفاده از fMRI حالت استراحت ممکن است کاربردهای بالقوهای برای نقشهبرداری از شناخت و احساسات در بیماران جراحی مغز و اعصاب داشته باشد. (۲۱) اهداف این فصل مرور: (1) مناطق و مکانیسم های درگیر در توجه انتخابی و حافظه کاری؛ (2) مکانیسم های تصمیم گیری، انگیزه و تخصیص تلاش. (3) حوزه های درگیر در کنترل شناختی؛ و (4) نمونه هایی از اختلالات موثر بر این شبکه های مقیاس بزرگ.
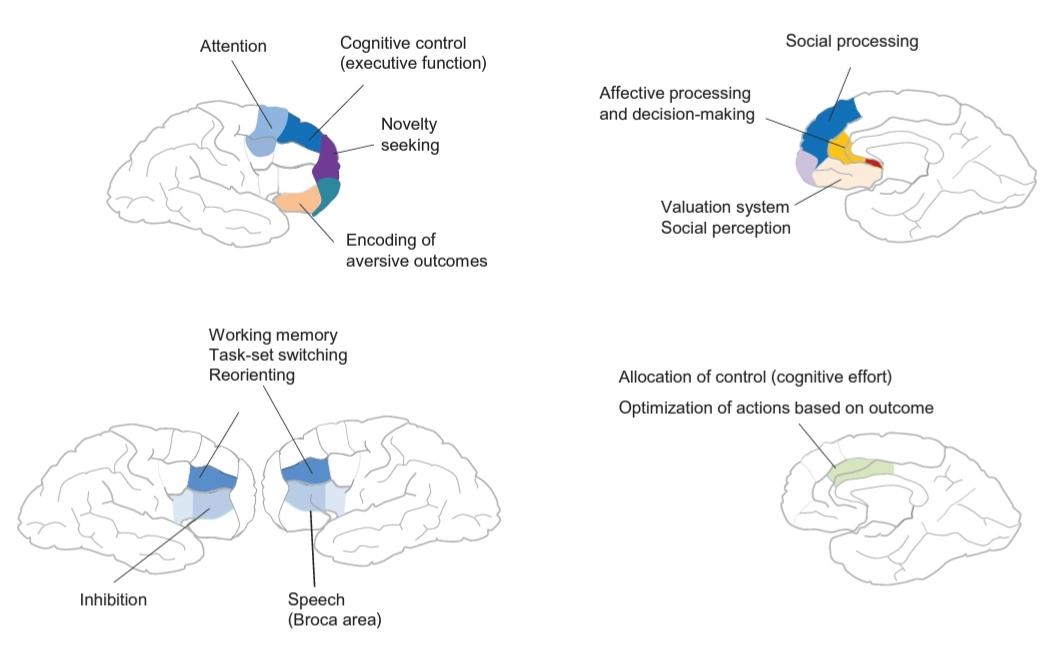
FIGURE 42.1 Overview of functions of the prefrontal cortex
Meta-analyses of functional neuroimaging studies has provided maps of the functional correlates of behavioral functions in the frontal cortex. Areas of the lateral prefrontal cortex (PFC) are subdivided according to their affiliation with other cortical areas and patterns of activation into those that form a part of parietal networks involved in executive functions, the default mode network involved in inner thought processing, and somatosensory networks involved in motor planning and programming). Similarly, medial frontal cortical areas have been subdivided into anterior, middle, and posterior regions according to their participation in networks involved in emotional/social, cognitive, and motor functions (not shown), respectively.
شکل 42.1 مروری بر عملکرد قشر پرهفرونتال
متاآنالیز مطالعات تصویربرداری عصبی عملکردی، نقشههایی از همبستگیهای عملکردی عملکردهای رفتاری در قشر پیشانی ارائه کرده است. نواحی قشر پرهفرونتال جانبی (PFC) بر اساس وابستگی به سایر نواحی قشری و الگوهای فعالسازی به آنهایی تقسیم میشوند که بخشی از شبکههای آهیانهای درگیر در عملکردهای اجرایی، شبکه حالت پیشفرض درگیر در پردازش فکر درونی، و شبکههای حسی پیکری درگیر در برنامهریزی و برنامهریزی حرکتی هستند. به طور مشابه، نواحی قشر پیشانی میانی بر اساس مشارکت آنها در شبکههای درگیر در عملکردهای عاطفی/اجتماعی، شناختی و حرکتی (نشان داده نشده) به نواحی قدامی، میانی و خلفی تقسیم شدهاند.
ATTENTION
Attention is the selection of task-relevant sensory information at the expense of nonrelevant information. (4) The best-studied form of attention is visual attention, using either visual spatial orienting (spatial location-based attention), or visual search (feature-based attention) paradigms. (22, 23) Visual spatial attention may include both the focusing of attention at a selected location and an automatic monitoring of other locations for flexibly gating the reallocation of attentional resources. Attentional processing may be biased by either top-down or bottom-up influences.
توجه
توجه، انتخاب اطلاعات حسی مرتبط با تکلیف به قیمت نادیده گرفتن اطلاعات غیرمرتبط است. (۴) بهترین شکل مورد مطالعه توجه، توجه بصری است، با استفاده از پارادایمهای جهت گیری فضایی بصری (توجه مبتنی بر مکان فضایی)، یا جستجوی بصری (توجه مبتنی بر ویژگی). (۲۳، ۲۲) توجه فضایی بصری ممکن است شامل تمرکز توجه در یک مکان انتخاب شده و نظارت خودکار سایر مکانها برای دسترسی انعطاف پذیر به منابع توجه به مکان واقعی باشد. پردازش توجه ممکن است با تأثیرات از بالا به پایین یا از پایین به بالا سوگیری داشته باشد.
Top-down processing is the goal-oriented voluntary guidance of attention to location, objects, and features of the environment that are relevant to the current behaviors and goals. Bottom-up attention is driven by salient external stimuli. Attention may be reoriented toward objects that are behaviorally relevant because they match current goals, require a completely different course of action, or are signaled as important on the basis of previous experience and longterm memory associations. (4) Reorienting to new objects may also occur reflexively based on their high sensory salience, for example, if they are unexpected based on their features or spatial location. Functional imaging studies show the presence of two separate frontoparietal attention networks, dorsal and ventral, which are defined by their functional connectivity and different roles in attentional processing (Figure 42.2).
پردازش از بالا به پایین، هدایت داوطلبانه هدفمحور توجه به مکان، اشیاء و ویژگیهای محیطی است که با رفتارها و اهداف فعلی مرتبط هستند. توجه از پایین به بالا توسط محرکهای خارجی برجسته هدایت میشود. توجه ممکن است به سمت اشیایی معطوف شود که از نظر رفتاری مرتبط هستند، زیرا با اهداف فعلی مطابقت دارند، به مسیر عمل کاملاً متفاوتی نیاز دارند، یا بر اساس تجربیات قبلی و تداعیهای حافظه بلندمدت به عنوان مهم نشان داده میشوند. (۴) مطالعات تصویربرداری عملکردی وجود دو شبکه توجه مجزای پیشانی، پشتی و شکمی را نشان میدهد که با اتصال عملکردی و نقشهای مختلف در پردازش توجه تعریف میشوند (شکل 42.2).
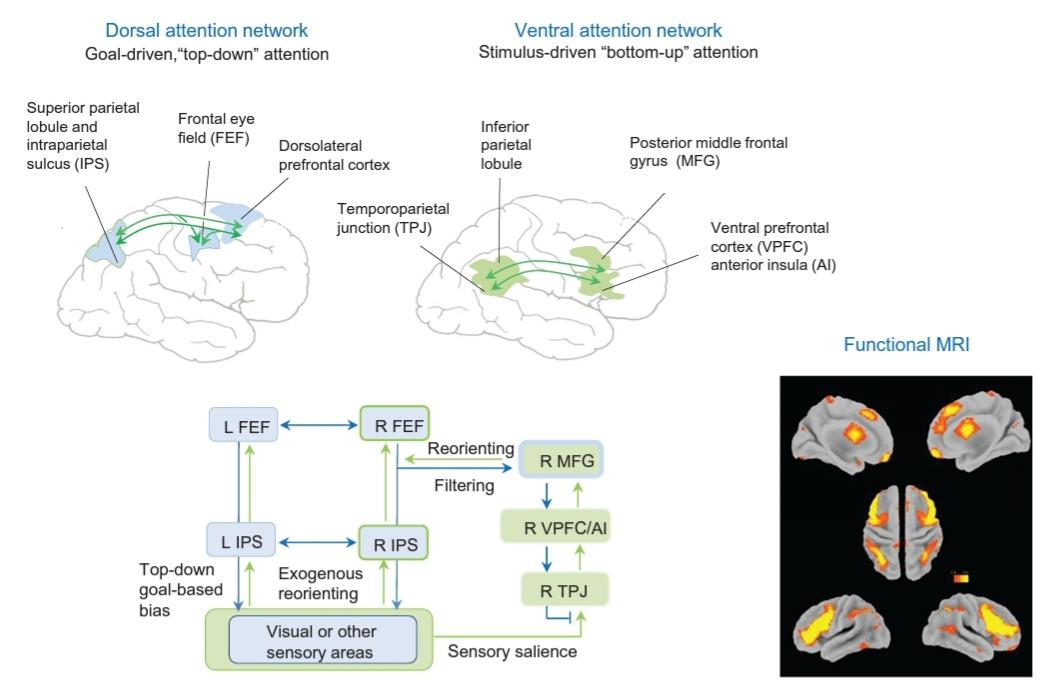
FIGURE 42.2 Attention networks
Functional imaging studies show the presence of two separate frontoparietal attention networks, dorsal and ventral. The dorsal frontoparietal attention network includes the dorsal parietal cortex, particularly intraparietal sulcus (lateral intraparietal area) and superior parietal lobule; the lateral PFC along the precentral sulcus; and the frontal eye field (FEF) and is involved in goal-oriented attention. The ventral frontoparietal network includes the temporoparietal junction and the ventral lateral frontal cortex (parts of middle frontal gyrus, inferior frontal gyrus, frontal operculum, and anterior insula). It is lateralized to the right hemisphere and registers salient events in the environment. The reorienting response is triggered by novel and unexpected stimuli and involves the action of the right hemisphere ventral frontoparietal network that interrupts and resets ongoing activity of the dorsal frontoparietal network. When attention is focused via the dorsal network based on an internal goal (goal-driven), the ventral network is suppressed to prevent reorienting to distracting events. (Functional MRI images courtesy of Dr. David Jones).
شکل 42.2 شبکههای توجه
مطالعات تصویربرداری عملکردی وجود دو شبکه توجه مجزای پیشانی، پشتی و شکمی را نشان میدهد. شبکه فرونتوپاریتال پشتی توجه شامل قشر آهیانهای پشتی، به خصوص شیار داخل آهیانهای (ناحیه داخل آهیانهای جانبی) و لوبول آهیانهای فوقانی. PFC جانبی در امتداد شیار پیش مرکزی. و میدان چشم جلویی (FEF) و در توجه هدف گرا نقش دارد. شبکه فرونتوپاریتال شکمی شامل محل اتصال گیجگاهی و قشر پیشانی جانبی شکمی (بخشهایی از شکنج فرونتال میانی، شکنج فرونتال تحتانی، اپرکولوم پیشانی و اینسولای قدامی) است. به سمت نیمکره راست جانبی میشود و رویدادهای برجسته در محیط را ثبت میکند. پاسخ جهتگیری مجدد توسط محرکهای جدید و غیرمنتظره ایجاد میشود و شامل عمل شبکه فرونتوپاریتال شکمی نیمکره راست میشود که فعالیتهای جاری شبکه فرونتوپاریتال پشتی را قطع و بازنشانی میکند. هنگامیکه توجه از طریق شبکه پشتی بر اساس یک هدف داخلی (هدف محور) متمرکز میشود، شبکه شکمی سرکوب میشود تا از جهت گیری مجدد به رویدادهای حواس پرتی جلوگیری شود. (تصاویر عملکردی MRI توسط دکتر دیوید جونز).
Dorsal Attention Network
The dorsal attention network includes the dorsal parietal cortex, particularly intraparietal sulcus and superior parietal lobule; the lateral PFC along the precentral sulcus;and the frontal eye field (FEF). (5, 6) This dorsal network is involved in goal-oriented attention. It enables the selection of sensory stimuli based on current internal goals or expectations, sends top-down signals to the sensory cortex to bias processing of appropriate stimulus features and locations, and then links the selected stimuli to appropriate motor responses. The dorsal attention network is activated by the expectation of seeing an object at a particular location or with certain features, by the preparation of a specific response, or by the short-term memory of a visual scene. Via the FEF, this network directs attention and gaze toward sensory stimuli appearing at unexpected locations in the contralateral hemispace. There is both segregation and strong correlation between the activity of the dorsal attention network and that of the visual network (including the primary visual and association visual areas) at rest. During a visual attention task that required dynamic and selective rerouting of visual information, there was a reduced of correlation of the ongoing “idling” activity within the visual network and an increase in the top-down connectivity from the dorsal attention to the visual network. (23)
شبکه پشتی توجه
شبکه پشتی توجه شامل قشر آهیانهای پشتی، به ویژه شیار داخل آهیانهای و لوبول آهیانهای فوقانی است. PFC جانبی در امتداد شیار پیش مرکزی و میدان چشم جلویی (FEF). (۵، ۶) این شبکه پشتی درگیر توجه هدف گرا است. انتخاب محرکهای حسی بر اساس اهداف یا انتظارات داخلی فعلی را امکانپذیر میسازد، سیگنالهایی را از بالا به پایین به قشر حسی برای پردازش سوگیری ویژگیها و مکانهای محرک مناسب میفرستد و سپس محرکهای انتخابشده را به پاسخهای حرکتی مناسب پیوند میدهد. شبکه پشتی توجه با انتظار دیدن یک شی در یک مکان خاص یا با ویژگیهای خاص، با تهیه یک پاسخ خاص، یا با حافظه کوتاه مدت یک صحنه بصری فعال میشود. از طریق FEF، این شبکه توجه و نگاه را به سمت محرکهای حسی هدایت میکند که در مکانهای غیرمنتظره در نیمفضای مقابل ظاهر میشوند. بین فعالیت شبکه پشتی توجه و شبکه بینایی (شامل نواحی بصری اولیه و ارتباطی) در حالت استراحت، هم تفکیک و هم ارتباط قوی وجود دارد. در طول یک کار توجه بصری که نیاز به مسیریابی مجدد پویا و انتخابی اطلاعات بصری داشت، کاهش همبستگی فعالیت “بیکار” جاری در شبکه بصری و افزایش اتصال از بالا به پایین از توجه پشتی به شبکه بصری وجود داشت. (۲۳)
Ventral Attention Network
The ventral attention network includes the cortex at the temporoparietal junction, including the posterior superior temporal sulcus, superior temporal gyrus, and ventral part of the supramarginal gyrus, and the ventral lateral frontal cortex, including parts of middle frontal gyrus, inferior frontal gyrus, frontal operculum, and anterior insula. This ventral network is lateralized to the right hemisphere and registers salient events in the environment not only in the visual but also in other sensory modalities and in social situations. A component of this network, the temporoparietal junction-posterior superior temporal sulcus, is highly expanded in humans and contains areas that are engaged not only in attention but also in processing of emotional expression of faces, theory of mind, and memory. (24) The expansion of the temporoparietal junction-posterior superior temporal sulcus has allowed for the elaboration of the cortical areas contained within this region. This has led to the proposal that the temporoparietal junctionposterior superior temporal sulcus is the basis of a third frontoparietal processing stream separate from the classical dorsal (“where”) and the ventral (“what” stream); this third stream would underlie the increased social abilities in humans by functioning as a hub that coordinates the activities of multiple brain networks in the exploration of the social scenes typical of the human social experience. (24) The ventral anterior network includes components of both the salience (25) and cingulo-opercular task control (26) networks, including the anterior insula and anterior MCC. (18) There is a rightward asymmetry in white matter connectivity strength between the temporoparietal junction and the insula, which supports the right-sided lateralization in stimulus-driven attention, sensory awareness, interocepthe duration of the perceptual episodes and thus attention, and pain. (27)
شبکه شکمی توجه
شبکه شکمی توجه شامل قشر در محل اتصال گیجگاهی، شامل شیار گیجگاهی فوقانی خلفی، شکنج گیجگاهی فوقانی، و قسمت شکمی شکنج سوپرامارجینال، و قشر پیشانی جانبی شکمی، شامل بخشهایی از شکنج فرونتال میانی، شکنج فرونتال تحتانی، شکنج فرونتال و قدامی است. این شبکه شکمی به سمت نیمکره راست جانبی میشود و رویدادهای برجسته در محیط را نه تنها در بصری، بلکه در سایر روشهای حسی و در موقعیتهای اجتماعی ثبت میکند. یکی از اجزای این شبکه، محل اتصال موقت آهیانهای-خلفی شیار زمانی فوقانی، در انسان بسیار گسترش یافته است و شامل مناطقی است که نه تنها درگیر توجه، بلکه در پردازش بیان احساسی چهرهها، نظریه ذهن، و حافظه هستند. (۲۴) این منجر به این پیشنهاد شده است که محل اتصال تمپوروپاریتال شیار تمپورال فوقانی خلفی، پایه سومین جریان پردازش پیشانی جدا از پشتی کلاسیک (“کجا”) و شکمی (جریان “چه”) است. این جریان سوم با عملکرد به عنوان مرکزی که فعالیتهای شبکههای مغزی متعدد را در کاوش صحنههای اجتماعی نمونهای از تجربه اجتماعی انسان هماهنگ میکند، زمینه ساز افزایش تواناییهای اجتماعی در انسانها میشود. (۲۴) شبکه قدامی شکمی شامل اجزای شبکههای کنترل تکلیف برجسته (۲۵) و سینگولو-اوپرکولار (۲۶) میشود، از جمله MCC قدامی و اینسولای قدامی (۱۸). یک عدم تقارن به سمت راست در قدرت اتصال ماده سفید بین اتصال تمپوروپاریتال و اینسولا وجود دارد، که از جانبی شدن سمت راست در توجه stimulus-driven، آگاهی حسی، مدت زمان بین اپیزودهای ادراکی و در نتیجه توجه و درد پشتیبانی میکند. (27)
Reorienting Response
The reorienting response is triggered by novel and unexpected stimuli. It involves the action of the right hemisphere dominant ventral frontoparietal network that interrupts and resets ongoing activity of the dorsal frontoparietal networks. (5) When attention is focused via the dorsal network based on an internal goal (goal-driven), the ventral network is suppressed to prevent reorienting to distracting events. Upon detection of behaviorally relevant objects or targets, attention is reoriented to the new source of information (stimulus-driven reorienting) as the ventral network interrupts ongoing selection in the dorsal network, which in turn shifts attention toward the novel object of interest. In contrast, the reduced response of the ventral attention network to unimportant objects prevents shifts of attention that could interfere with the performance of a goal-oriented task. This “filtering” response reflects a top-down suppression signal from the dorsal attention network as well as the input from executive control networks acting at the level of the right temporoparietal junction. The right posterior middle frontal gyrus may be a major link between the ventral and dorsal networks.
جهت گیری مجدد پاسخ
پاسخ جهت گیری مجدد توسط محرکهای بدیع و غیر منتظره ایجاد میشود. این شامل عملکرد شبکه فرونتوپاریتال شکمی غالب نیمکره راست است که فعالیت مداوم شبکههای فرونتوپاریتال پشتی را قطع و تنظیم مجدد میکند. (۵) هنگامیکه توجه از طریق شبکه پشتی بر اساس یک هدف داخلی (هدف محور) متمرکز میشود، شبکه شکمی سرکوب میشود تا از جهت گیری مجدد به رویدادهای حواس پرتی جلوگیری شود. با شناسایی اشیا یا اهداف مرتبط رفتاری، توجه به منبع جدید اطلاعات معطوف میشود (جهتیابی مجدد بر مبنای محرک) زیرا شبکه شکمی انتخاب مداوم در شبکه پشتی را قطع میکند، که به نوبه خود توجه را به سمت شی جدید مورد علاقه سوق میدهد. در مقابل، کاهش پاسخ شبکه شکمی توجه به اشیاء بیاهمیت، از جابهجایی توجهی که میتواند در عملکرد یک وظیفه هدفمدار اختلال ایجاد کند، جلوگیری میکند. این پاسخ “فیلتر کردن” یک سیگنال سرکوب از بالا به پایین از شبکه پشتی توجه و همچنین ورودی از شبکههای کنترل اجرایی که در سطح اتصال تمپوپارتیتال سمت راست عمل میکنند را منعکس میکند. شکنج میانی پیشانی خلفی راست ممکن است پیوند اصلی بین شبکههای شکمی و پشتی باشد.
The different patterns of recruitment of the dorsal and ventral attention networks during goal-oriented behavior and reorienting responses may reflect inputs from the locus coeruleus (LC) noradrenergic system. During focused attention, LC neurons may have a phasic response to target stimuli that shares many similarities with the P300 targetrelated cortical evoked potential. This response may function as an “interrupt” or “network reset” signal that may allow the flexible configuration of the attention networks once a target is detected. The decrease in tonic LC activity during the transition from an exploratory state to a taskfocused state may parallel the deactivation of temporoparietal junction, which would promote engagement on the current task and filtering of distractors. (5, 28)
الگوهای مختلف بهکارگیری شبکههای توجه پشتی و شکمی در طول رفتار هدفگرا و پاسخهای جهتگیری مجدد ممکن است ورودیهای سیستم نورآدرنرژیک لوکوس سرولئوس (LC) را منعکس کند. در طول توجه متمرکز، نورونهای LC ممکن است یک پاسخ فازی به محرکهای هدف داشته باشند که شباهتهای زیادی با پتانسیل برانگیخته قشر مرتبط با هدف P300 دارد. این پاسخ ممکن است به عنوان یک سیگنال “وقفه” یا “بازنشانی شبکه” عمل کند که ممکن است امکان پیکربندی انعطاف پذیر شبکههای توجه را پس از شناسایی هدف فراهم کند. کاهش فعالیت LC تونیک در طول انتقال از حالت اکتشافی به حالت متمرکز بر وظیفه ممکن است به موازات غیرفعال کردن محل اتصال گیجگاهی باشد که درگیری با کار فعلی و فیلتر کردن عوامل حواسپرتی را تقویت میکند.(۲۸، ۵)
Perceptual Episodes and Attentional Blink
Attention is deployed over an interval that frequently encompasses several stimuli. When numerous stimuli are densely packed in time, attentional selection occurs in brief perceptual episodes. The ability to identify a target is usually hindered if it appears shortly after another target. Two episodes of attentional selection cannot occur very close in time. When two items are presented briefly and in close succession, observers can usually report the first but often fail to report the second. This phenomenon is known as the attentional blink. (29-31) A second attentional episode may occur only when the second target follows the first by more than 100 to 250 ms. There are several hypotheses regarding the basis of attentional blink. Evidence indicates that both cognitive control and emotion strongly influence the duration of perceptual episodes and thus attentional blink. (29-31)
اپیزودهای ادراکی و چشمک زدن توجه
توجه در بازهای مستقر میشود که غالباً چندین محرک را در بر میگیرد. هنگامیکه محرکهای متعدد به طور فشرده در زمان بسته بندی میشوند، انتخاب توجه در قسمتهای ادراکی کوتاه اتفاق میافتد. توانایی شناسایی یک هدف معمولاً اگر مدت کوتاهی پس از یک هدف دیگر ظاهر شود، مانع میشود. دو قسمت از انتخاب توجه نمیتواند در زمان بسیار نزدیک رخ دهد. هنگامیکه دو مورد به طور خلاصه و پشت سر هم ارائه میشوند، ناظران معمولا میتوانند اولی را گزارش کنند اما اغلب در گزارش دومیشکست میخورند. این پدیده به چشمک زدن توجه معروف است. (۳۱-۲۹) قسمت دوم توجه ممکن است تنها زمانی رخ دهد که هدف دوم با فاصله بیش از 100 تا 250 میلی ثانیه از هدف اول پیروی کند. فرضیههای متعددی در مورد اساس چشمک زدن توجه وجود دارد. شواهد نشان میدهد که هم کنترل شناختی و هم هیجان به شدت بر طول دورههای ادراکی و در نتیجه چشمک زدن توجه تأثیر میگذارد.(۳۱-۲۹)
Value-Driven Guidance of Visual Attention
One fundamental component of attention is the attraction of gaze direction toward objects with high reward value. (32) This decision-making process is regulated by the caudate nucleus, which is a component of the oculomotor circuit of the basal ganglia. This circuit is integrated to the dorsal attention network and includes the FEF, substantia nigra pars reticulata (SNr), and superior colliculus. The SNr tonically inhibits saccadic burst neurons of the superior colliculus; in response to a reward signaled by a peak of dopamine, inhibitory inputs from the caudate nucleus to the SNr transiently interrupt the tonic inhibition of the superior colliculus, thus promoting the initiation of a saccadic eye movement toward the high-value objects of attention. (32) A caudate head circuit relays on short-term value memories to trigger eye deviation toward recently valued objects; a caudate tail circuit relays on long-term reward memories to allow the subject’s gaze to move automatically to a previously valued object. (32)
راهنمای ارزش محور توجه بصری
یکی از مؤلفههای اساسی توجه، جذب جهت نگاه به سمت اجسام با ارزش پاداش بالا است. (۳۲) این فرآیند تصمیمگیری توسط هسته دمیتنظیم میشود، که جزء مدار چشمیحرکتی عقدههای پایه است. این مدار به شبکه پشتی توجه ادغام شده است و شامل FEF، ماده سیاه پارس مشبک (SNr) و colliculus برتر است. SNr به طور تونیکی نورونهای ساکادیک ترکیدگی کولیکولوس فوقانی را مهار میکند. در پاسخ به پاداشی که توسط یک پیک دوپامین سیگنال میشود، ورودیهای بازدارنده از هسته دمی به SNr به طور گذرا مهار تونیک کولیکولوس برتر را قطع میکند، بنابراین شروع یک حرکت ساکادیک چشم را به سمت اشیاء با ارزش بالای سر در مدارهای کوتاهمدت هد بهمنظور تقویت میکند. (۳۲) یک مدار caudate head روی حافظههای ارزشی کوتاهمدت رله میکند تا انحراف چشم را به سمت اشیایی که اخیراً ارزش گذاری شدهاند، ایجاد کند. یک مدار caudate tail روی حافظههای پاداش بلندمدت رله میکند تا به سوژه اجازه دهد به طور خودکار به سمت یک شی که قبلاً ارزش گذاری شده است حرکت کند. (32)
Effects of Attention on Sensory Processing and Cortical Oscillations
The dorsolateral PFC and components of the dorsal frontoparietal attention network, including the FEF and lateral intraparietal area, provide modulatory attention-related signals that are fed back to sensory cortex. (33) Neurons in the parietal cortex encode a salience map of the visual scene, encoding which locations in space are of potentially high significance, whereas neurons in frontal cortex carry information about task-relevant stimuli. Visual attention elicits an increase in the high-frequency (gamma, 40-80 Hz) oscillations and decreased the low-frequency (<10 Hz) oscillations in populations of neurons in the visual cortex representing the attended location. There is also synchrony between the frontal and parietal cortex during attentional tasks. There is gamma band synchrony when attention is captured by external stimuli and information from the parietal to the frontal cortex (bottom-up) and beta band synchrony when attention is internally controlled and information flows from the prefrontal to parietal cortex.
اثرات توجه بر پردازش حسی و نوسانات قشر مغز
PFC پشتی جانبی و اجزای شبکه توجه جلویی-پاریتال پشتی، از جمله FEF و ناحیه داخل آهیانهای جانبی، سیگنالهای تعدیلی مرتبط با توجه را ارائه میدهند که به قشر حسی بازخورد میشوند. (۳۳) نورونها در قشر آهیانهای، نقشه برجستهای از صحنه بینایی را رمزگذاری میکنند، که در آن مکانهای بالقوه نورونها در فضای قشر با علامت بالا کدگذاری میشوند. اطلاعات در مورد محرکهای مربوط به کار. توجه بصری باعث افزایش نوسانات با فرکانس بالا (گاما، 40-80 هرتز) و کاهش نوسانات فرکانس پایین (<10 هرتز) در جمعیتهای نورونها در قشر بینایی میشود که محل حضور را نشان میدهند. همچنین همزمانی بین قشر پیشانی و آهیانهای در حین انجام وظایف توجه وجود دارد. زمانی که توجه توسط محرکهای بیرونی و اطلاعات از قشر آهیانهای به قشر پیشانی (از پایین به بالا) جذب میشود، همزمانی باند گاما وجود دارد و زمانی که توجه به صورت درونی کنترل میشود و اطلاعات از قشر پرهفرونتال به قشر آهیانهای جریان مییابد.
Spatial attention to a visual stimulus increases neural responses to the selected stimulus across many levels of processing, including primary and secondary visual areas as well as subcortical regions such as the lateral geniculate nucleus, pulvinar, reticular nucleus of the thalamus, and superior colliculus. Interactions between excitatory pyramidal neurons and inhibitory interneurons are central to the mechanism supporting attention. Interneuronmediated lateral inhibition suppresses competing stimuli; attention has a much larger impact on the responses of inhibitory interneurons than in pyramidal cells. Activity of inhibitory interneurons also leads to the generation of high-frequency synchronous oscillations within a cortical region. Inter-areal synchronization occurs as these local oscillations synchronize along with the propagation of a bottom-up sensory drive.
توجه فضایی به یک محرک بینایی پاسخهای عصبی به محرک انتخابی را در سطوح مختلف پردازش افزایش میدهد، از جمله نواحی بینایی اولیه و ثانویه و همچنین نواحی زیر قشری مانند هسته ژنیکولیت جانبی، پولوینار، هسته مشبک تالاموس، و کولیکلوس فوقانی. فعل و انفعالات بین نورونهای هرمی تحریکی و نورونهای بازدارنده در مکانیسم حمایت کننده از توجه مرکزی است. مهار جانبی با واسطه بین عصبی، محرکهای رقابتی را سرکوب میکند. توجه تأثیر بسیار بیشتری بر پاسخهای نورونهای بازدارنده نسبت به سلولهای هرمی دارد. فعالیت نورونهای بازدارنده نیز منجر به تولید نوسانات همزمان با فرکانس بالا در یک ناحیه قشر مغز میشود. همزمانی بین ناحیه ای زمانی رخ میدهد که این نوسانات محلی همراه با انتشار یک درایو حسی از پایین به بالا همگام میشوند.
A “reverse hierarchy” model of visual attention suggests that selection begins in the prefrontal and parietal cortices, where neurons represent abstract categories, and then filters backward along the cortical hierarchy to “simpler” visual areas. Spatial and feature-based attention interact with one another; attending to a spatial location would select an object at that location. Spatial attention can drive featurebased attention (e.g., attending to a single object in space leads to increases in attention to its properties across the entire cortex); conversely, feature-based attention can drive spatial attention (e.g., the detection of a car in a visual scene drives spatial attention to that location).
یک مدل «سلسله مراتب معکوس» توجه بصری نشان میدهد که انتخاب در قشرهای جلوی پیشانی و آهیانهای آغاز میشود، جایی که نورونها مقولههای انتزاعی را نشان میدهند، و سپس به سمت عقب در امتداد سلسله مراتب قشری به سمت مناطق بصری «سادهتر» فیلتر میشوند. توجه فضایی و مبتنی بر ویژگی با یکدیگر تعامل دارند. توجه به یک مکان فضایی یک شی را در آن مکان انتخاب میکند. توجه فضایی میتواند توجه مبتنی بر ویژگی را برانگیزد (به عنوان مثال، توجه به یک شی واحد در فضا منجر به افزایش توجه به ویژگیهای آن در کل قشر میشود). برعکس، توجه مبتنی بر ویژگی میتواند توجه فضایی را جلب کند (به عنوان مثال، تشخیص یک ماشین در یک صحنه بصری توجه فضایی را به آن مکان سوق میدهد).
Thalamic Mechanisms of Attention
Communication in cortical large-scale networks during attention is mediated not only through cortico-cortical interactions but also through cortico-thalamo-cortical interactions involving the pulvinar and the mediodorsal thalamic nucleus. During spatial selection, interactions via the pulvinar increase synchrony of slow alpha oscillations between two interconnected visual cortical areas. During sustained visuospatial attention, alpha activity in the pulvinar reflects the precise temporal coordination of cortical activity via phasic inhibition of projection neurons in the posterior parietal cortex. Alpha band synchronization driven by posterior parietal cortex to the pulvinar may act to gate or suppress the processing of incoming sensory information at early stages of visual cortex. The major thalamic input to the PFC originates in the mediodorsal nucleus. Projections from this nucleus promote relevant recurrent synaptic interactions in the PFC, which maintains representations of task-rules, for example those required for sensory selection. Via this mechanism, the different subdivisions of the mediodorsal nucleus participate in multiple cognitive processes, including attention, working memory, familiarity-based recognition and retrieval, and emotional processing. The medial (magnocellular) portion of the mediodorsal nucleus is a component of a parahippocampal prefrontal system involved in familiarity-based recognition and retrieval processing, and also has a central role in emotional processing by interconnecting the amygdala and the posterior orbitofrontal cortex. (34) The lateral (parvocellular) division of the mediodorsal nucleus via its reciprocal connections with the dorsolateral PFC has a major role in working memory; both areas contain neurons that sustain tonic activity during the delay period between the presentation of the cue and the motor output.
مکانیسمهای توجه تالاموس
ارتباط در شبکههای مقیاس بزرگ قشر مغز در حین توجه نه تنها از طریق فعل و انفعالات قشر مغز بلکه از طریق فعل و انفعالات کورتیکو-تالامو-قشری که شامل هسته تالاموس پولوینار و مدیودورسال میشود، انجام میشود. در طول انتخاب فضایی، فعل و انفعالات از طریق پولوینار باعث افزایش همزمانی نوسانات آلفای آهسته بین دو ناحیه قشری بصری به هم پیوسته میشود. در طول توجه دیداری-فضایی پایدار، فعالیت آلفا در پالوینار منعکس کننده هماهنگی زمانی دقیق فعالیت قشر مغز از طریق مهار فازی نورونهای برآمده در قشر آهیانهای خلفی است. همگامسازی باند آلفا که توسط قشر آهیانهای خلفی به سمت پالوینار هدایت میشود، ممکن است پردازش اطلاعات حسی ورودی را در مراحل اولیه قشر بینایی مهار کند. ورودی اصلی تالاموس به PFC از هسته میانی نشات میگیرد. پیشبینیهای این هسته، برهمکنشهای سیناپسی مربوطه را در PFC ترویج میکنند، که بازنماییهایی از قوانین وظیفه را حفظ میکند، بهعنوان مثال قوانین مورد نیاز برای انتخاب حسی. از طریق این مکانیسم، زیرشاخههای مختلف هسته میانی در فرآیندهای شناختی متعددی از جمله توجه، حافظه کاری، شناخت و بازیابی مبتنی بر آشنایی و پردازش هیجانی شرکت میکنند. بخش داخلی (مگنوسلولار) هسته میانی بخشی از یک سیستم پرهفرونتال پاراهیپوکامپ است که در شناخت و پردازش بازیابی مبتنی بر آشنایی نقش دارد و همچنین با به هم پیوستن آمیگدال و قشر اوربیتوفرونتال خلفی، نقش مرکزی در پردازش هیجانی دارد. (34) تقسیم جانبی (پاروسلولار) هسته میانی پشتی از طریق اتصالات متقابل آن با PFC پشتی جانبی نقش مهمی در حافظه کاری دارد. هر دو ناحیه حاوی نورونهایی هستند که فعالیت تونیک را در طول دوره تاخیر بین ارائه نشانه و خروجی حرکتی حفظ میکنند.
The claustrum may also have an important role in at tention. (35) This thin, sheet-like structure located between the insular cortex and the putamen is characterized by its reciprocal and topographically organized connections with all neocortical areas including visual, somatosensory, auditory, and motor cortical regions. The claustrum communicates most densely with medial frontal cortical regions and receives prominent inputs from the amygdala, hippocampus, and mediodorsal nucleus of the thalamus. Glutamatergic projections from the claustrum target pyramidal neurons in supra and infragranular layers but primarily terminate on local inhibitory neurons in layer 4, thereby promoting feedforward inhibition of incoming information in multiple cortical areas.35 The claustrum may also orchestrate and integrate cortical activity by boosting synchronized cortical oscillations.
کلاستروم نیز ممکن است نقش مهمیدر توجه داشته باشد. (۳۵) این ساختار ورقه مانند و نازک که بین قشر اینسولا و پوتامن قرار دارد با اتصالات متقابل و سازمان یافته توپوگرافی آن با تمام نواحی نئوکورتیکال از جمله نواحی بینایی، حسی تنی، شنوایی و حرکتی مشخص میشود. کلاستروم به شدت با نواحی قشر پیشانی میانی ارتباط برقرار میکند و ورودیهای برجسته ای را از آمیگدال، هیپوکامپ و هسته میانی تالاموس دریافت میکند. پروجکشنهای گلوتاماترژیک از کلاستروم، نورونهای هرمی را در لایههای سوپراگرانولار و اینفراگرانولار مورد هدف قرار میدهند، اما در درجه اول به نورونهای بازدارنده موضعی در لایه 4 ختم میشوند، در نتیجه مانع از اطلاعات دریافتی در مناطق متعدد قشری میشوند. (35) کلاستروم همچنین ممکن است با تقویت نوسانات همگام قشری، فعالیت قشر مغز را هماهنگ و یکپارچه کند.
WORKING MEMORY
Working memory is the “mental sketchpad” for shortterm maintenance of information over brief periods of time (several seconds to minutes) in the absence of sensory input; this process is commonly referred to as “short-term memory.” However, in addition working memory allows manipulation and processing of the transiently stored information for guidance of behavior. Both selective attention and cognitive control operations are integral parts of working-memory processing. (7, 8, 36)
حافظه کاری
حافظه کاری “صفحه طراحی ذهنی” برای نگهداری کوتاه مدت اطلاعات در دورههای زمانی کوتاه (چند ثانیه تا دقیقه) در غیاب ورودی حسی است. این فرآیند معمولاً به عنوان “حافظه کوتاه مدت” شناخته میشود. با این حال، علاوه بر این، حافظه کاری امکان دستکاری و پردازش اطلاعات ذخیره شده موقتی را برای هدایت رفتار فراهم میکند. هر دو عملیات توجه انتخابی و کنترل شناختی بخش جدایی ناپذیر پردازش حافظه کاری هستند. (۳۶، ۸، ۷)
Features of Working Memory
There are several theoretical models of working memory. (37, 38) A well-known model is that working memory involves four components, a “phonological loop” for language processing; a “visuospatial sketchpad” for visual processing; an “episodic buffer that allows features to be bound in “chunks,” such as shape and color or words in a sentence; and finally a “central executive” involved in focusing of attention, dividing attention along parallel verbal or visual tracks, switching between tracks, and interacting with longterm memory processing. (37) Working memory requires both the storage of sensory information in modality-specific posterior cortical areas during the delay period when the stimulus is no longer present, and recurrent activity in the lateral PFC to focus attention on the relevant stimulus, select information from distractors, and process the held information according to the task. Neurophysiological findings support a separation of prefrontal frontal executive and storage mechanisms in the posterior parietal and temporal cortex. An active maintenance process through reverberating activity provides attentional top-down signals to regions involved in perceptual and long-term memory representations related to the current content of working memory. Both sustained attention and the rehearsal process are crucial for maintaining the information in working memory. This process is potentially vulnerable to distraction and interference by task-irrelevant information. Workingmemory capacity is limited; most estimates of the average capacity among healthy young adults suggest that it has a capacity limit of approximately three or four simple items. Capacity increases if the items can be grouped into meaningful units, or “chunks,” which can exploit preexisting information about concepts already stored in long-term memory. Working-memory changes across the lifespan and can be modified by training. Recent models assume that short-term retention of information in working memory requires the allocation of attention to internal representations in long-term memory. When a subject is presented with symbolic information to be remembered (e.g., a list of names or a telephone number), the long-term memory representations of this information are accessed and subsequently maintained via attentional mechanism in an elevated state of activation until this information is no longer needed to achieve a goal.
ویژگیهای حافظه کاری
چندین مدل نظری از حافظه کاری وجود دارد. (۳۸، ۳۷) یک مدل معروف این است که حافظه کاری شامل چهار جزء است، یک “حلقه واجی” برای پردازش زبان. یک “صفحه تصویری فضایی” برای پردازش بصری. یک «حافظ اپیزودیک» که به ویژگیها اجازه میدهد در «تکهها» مانند شکل و رنگ یا کلمات در یک جمله محدود شوند؛ و در نهایت یک «مجری مرکزی» درگیر در تمرکز توجه، تقسیم توجه در مسیرهای کلامییا بصری موازی، جابهجایی بین آهنگها و تعامل با پردازش حافظه بلندمدت. (۳۷) حافظه کاری هم به ذخیره اطلاعات حسی در نواحی قشر خلفی خاص مدالیته در طول دوره تاخیر زمانی که محرک دیگر وجود ندارد و هم به فعالیت مکرر در PFC جانبی نیاز دارد تا توجه را بر محرک مربوطه متمرکز کند، اطلاعات را از حواس پرت کنندهها انتخاب کند، و اطلاعات نگهداری شده را بر اساس وظیفه پردازش کند. یافتههای نوروفیزیولوژیک از جداسازی مکانیسمهای اجرایی و ذخیرهسازی پیشانی پیشانی در قشر آهیانهای خلفی و گیجگاهی حمایت میکنند. یک فرآیند نگهداری فعال از طریق فعالیت طنین انداز، سیگنالهای توجهی از بالا به پایین را به مناطق درگیر در بازنمایی حافظه ادراکی و بلندمدت مرتبط با محتوای فعلی حافظه کاری ارائه میدهد. هم توجه پایدار و هم فرآیند تمرین برای حفظ اطلاعات در حافظه کاری بسیار مهم هستند. این فرآیند به طور بالقوه در برابر حواس پرتی و تداخل اطلاعات غیر مرتبط با کار آسیب پذیر است. ظرفیت حافظه کاری محدود است. اکثر برآوردها از میانگین ظرفیت در میان بزرگسالان جوان سالم نشان میدهد که ظرفیت آن تقریباً سه یا چهار مورد ساده است. ظرفیت افزایش مییابد اگر آیتمها را بتوان به واحدهای معنادار یا «تکههایی» گروهبندی کرد که میتوانند از اطلاعات موجود درباره مفاهیمیکه قبلاً در حافظه بلندمدت ذخیره شدهاند بهرهبرداری کنند. حافظه کاری در طول عمر تغییر میکند و میتوان آن را با آموزش تغییر داد. مدلهای اخیر فرض میکنند که نگهداری کوتاه مدت اطلاعات در حافظه کاری مستلزم تخصیص توجه به بازنماییهای داخلی در حافظه بلند مدت است. هنگامیکه اطلاعات نمادینی برای به خاطر سپردن به موضوعی ارائه میشود (مثلاً فهرستی از نامها یا شماره تلفن)، بازنمایی حافظه بلند مدت این اطلاعات قابل دسترسی است و متعاقباً از طریق مکانیسم توجه در حالت فعال سازی بالا نگهداری میشود تا زمانی که این اطلاعات دیگر برای رسیدن به هدف مورد نیاز نباشد.
Brain Areas and Connections Involved in Working Memory
Working memory results from the dynamic interaction of a large number of brain regions (Figure 42.3). The specific regions involved depend on a number of factors, including the type of material to be remembered, the task, and the stage of the dynamic interplay of processing components. (7, 8)
نواحی مغز و اتصالات درگیر در حافظه کاری
حافظه کاری ناشی از تعامل پویا تعداد زیادی از مناطق مغز است (شکل 42.3). مناطق خاص درگیر به عوامل متعددی از جمله نوع ماده ای که باید به خاطر بسپارید، وظیفه و مرحله تعامل پویا اجزای پردازش بستگی دارد. (۸، ۷)
The current view of the mechanisms underlying working memory is based on persistent activity in a recurrent network with fixed connectivity via reverberant excitatory circuits in areas 9/46 of the lateral PFC. The strong lateral inhibition between memory representations causes a winner-take-all dynamics that typically leaves only the winning representation active. The strength of dorsolateral PFC activity varies with the state of arousal: workingmemory abilities are greatly impaired during fatigue or stress. (39) Maintenance of working memory results from the interactions between selective attentional processing of sensory information and long-term memory representations. These interactions involve reentrant loops between frontal and posterior cortical areas encoding perceptual information, as well as subcortical structures such as the basal ganglia and the cerebellum. The same brain regions dedicated to sensory processing store sensory information during delay periods during performance of working-memory tasks. Therefore, there is not a single but rather multiple working-memory network interactions. (8) For example, spatial working-memory tasks commonly activate parietal cortex bilaterally predominantly on the right hemisphere. Different stimulus categories, such as faces and houses, activate category-specific regions of the ventral visual cortex. The left inferior parietal superior temporal and ventral prefrontal regions are activated during verbal working memory. The lateral PFC is critical for information maintenance during all working-memory tasks. (9) Meta-analyses have demonstrated that the left, particularly ventrolateral, PFC is more involved in verbal working-memory tasks, whereas the right, particularly dorsolateral, PFC is more involved in spatial working-memory tasks. There is substantial overlap between the neural mechanisms supporting working memory and those involved in attention and cognitive control. Together with the lateral PFC, the superior parietal cortex has also been associated with executive aspects of working memory. Both the cerebellum and the striatum are also commonly activated in working-memory tasks. Neuroimaging studies have shown that activity of the medial temporal lobe may be needed for working-memory tasks that require binding and relational processing associated with stored memories. (8)
دیدگاه فعلی مکانیسمهای زیربنایی حافظه کاری مبتنی بر فعالیت مداوم در یک شبکه بازگشتی با اتصال ثابت از طریق مدارهای تحریکی پژواک در مناطق 9/46 PFC جانبی است. مهار جانبی قوی بین بازنماییهای حافظه باعث ایجاد یک پویایی برنده میشود که معمولاً فقط نمایش برنده را فعال میکند. قدرت فعالیت PFC پشتی-جانبی با حالت برانگیختگی متفاوت است: تواناییهای حافظه کاری در طول خستگی یا استرس بسیار مختل میشوند. (39) حفظ حافظه کاری ناشی از تعامل بین پردازش توجه انتخابی اطلاعات حسی و بازنمایی حافظه بلند مدت است. این فعل و انفعالات شامل حلقههای ورودی مجدد بین نواحی قشر پیشانی و خلفی است که اطلاعات ادراکی را رمزگذاری میکند، و همچنین ساختارهای زیر قشری مانند عقدههای قاعدهای و مخچه. همان مناطق مغز که به پردازش حسی اختصاص داده شده اند، اطلاعات حسی را در طول دورههای تاخیر در انجام وظایف حافظه کاری ذخیره میکنند. بنابراین، یک تعامل شبکه با حافظه کاری واحد، بلکه چندگانه وجود ندارد. (۸) به عنوان مثال، وظایف حافظه کاری فضایی معمولاً قشر آهیانهای را به صورت دو طرفه و عمدتاً در نیمکره راست فعال میکنند. دستههای محرک مختلف، مانند چهرهها و خانهها، نواحی خاص دستهبندی قشر بینایی شکمی را فعال میکنند. نواحی پرهفرونتال فوقانی آهیانه تحتانی چپ در طول حافظه کاری کلامیفعال میشوند. PFC جانبی برای حفظ اطلاعات در طول تمام وظایف حافظه کاری حیاتی است. (۹) متاآنالیزها نشان داده اند که PFC سمت چپ، به ویژه بطنی جانبی، بیشتر درگیر وظایف حافظه کاری کلامی است، در حالی که سمت راست، به ویژه پشتی جانبی، PFC بیشتر در وظایف حافظه کاری فضایی درگیر است. همپوشانی قابل توجهی بین مکانیسم های عصبی حامی حافظه فعال و آنهایی که در توجه و کنترل شناختی نقش دارند وجود دارد. همراه با PFC جانبی، قشر جداری فوقانی نیز با جنبه های اجرایی حافظه کاری مرتبط است. مخچه و مخطط نیز معمولاً در وظایف حافظه کاری فعال می شوند. مطالعات تصویربرداری عصبی نشان دادهاند که فعالیت لوب گیجگاهی داخلی ممکن است برای کارهای حافظه کاری که نیاز به پردازش پیوندی و رابطهای مرتبط با خاطرات ذخیرهشده دارند، مورد نیاز باشد. (8)

FIGURE 42.3 Areas involved in working memory
Working memory results from the dynamic interaction of a large number of brain regions. Maintenance of working memory involves reentrant loops between frontal and posterior cortical areas encoding perceptual information. Spatial working-memory tasks commonly activate parietal cortex bilaterally predominantly on the right hemisphere; stimulus categories, such as faces and houses, activate category-specific regions of the ventral visual cortex; and the left inferior parietal superior temporal and ventral prefrontal regions are activated during verbal working memory. The lateral prefrontal cortex is critical for information maintenance and manipulation during all working-memory tasks (Functional MRI images courtesy of Dr. David Jones).
شکل 42.3 مناطق درگیر در حافظه کاری
حافظه کاری ناشی از تعامل پویا تعداد زیادی از مناطق مغز است. حفظ حافظه کاری شامل حلقههای ورودی مجدد بین نواحی قشر پیشانی و خلفی است که اطلاعات ادراکی را کد میکند. وظایف حافظه کاری فضایی معمولاً قشر آهیانهای را به طور دوطرفه و عمدتاً در نیمکره راست فعال میکنند. دستههای محرک، مانند چهرهها و خانهها، مناطق خاص دستهبندی قشر بینایی شکمی را فعال میکنند. و نواحی پرهفرونتال گیجگاهی و شکمی فوقانی آهیانه تحتانی چپ در طول حافظه کاری کلامیفعال میشوند. قشر پرهفرونتال جانبی برای نگهداری و دستکاری اطلاعات در طول تمام وظایف حافظه کاری حیاتی است (تصاویر MRI عملکردی توسط دکتر دیوید جونز).
Processing Hierarchy in the Lateral Frontal Cortex
As discussed previously, a key feature of human cortical organization is the spatial arrangement of areas along a global gradient between sensory motor and transmodal regions. (17) Functional MRI studies show that there is a functional gradient along the anterior-to-posterior axis of the lateral frontal cortex, representing a processing hierarchy according to the task involved. Simple response tasks activate the lateral premotor cortex (area 6), feature-based tasks activate area 8; dimension-based tasks activate the inferior frontal sulcus (areas 45 and 9/46); and task contexts activate the frontopolar cortex (area 10). Thus, the frontopolar regions are preferentially activated in relationship to the abstraction level of goals and task rules, whereas activity toward posterior portions of the lateral frontal cortex becomes progressively more specific to certain situational contexts. This hierarchical organization indicates that the frontopolar cortex influences information processing in posterior regions via widespread back projections down to area 6 at the lowest level. The frontopolar cortex is at the highest of cognitive processing and may have a major role functions such as novelty seeking, multitasking, and prospective thinking. Multitasking requires the ability to hold goals in mind while performing or processing secondary subgoals (“cognitive branching”), ability to remember to carry out an intended act in the future while engaging in another task, and effectively managing time while switching tasks. Prospection, imagining the future, or “metacognition,” requires reality monitoring, introspection, self-judgment, and analogical reasoning and involves interactions with components of the default network, which constitutes the final transmodal region along the gradient of cortical organization. (17)
پردازش سلسله مراتب در قشر پیشانی جانبی
همانطور که قبلاً بحث شد، یک ویژگی کلیدی سازمان کورتیکال انسان، آرایش فضایی مناطق در امتداد یک گرادیان جهانی بین نواحی حسی حرکتی و ترانسمودال است. (۱۷) مطالعات MRI عملکردی نشان میدهد که یک گرادیان عملکردی در امتداد محور قدامی به خلفی قشر پیشانی جانبی وجود دارد، که نشاندهنده یک سلسلهمراتب پردازش با توجه به تکلیف درگیر است. تکالیف پاسخ ساده قشر پرهموتور جانبی (ناحیه 6) را فعال میکند، تکالیف مبتنی بر ویژگی ناحیه 8 را فعال میکند. تکالیف مبتنی بر بُعد، شیار فرونتال تحتانی را فعال میکند (مناطق 45 و 9/46) و بافتهای تکلیف قشر فرونتوپلار (ناحیه 10) را فعال می کنند. بنابراین، مناطق فرونتوپلار ترجیحاً در رابطه با سطح انتزاع اهداف و قوانین تکلیف فعال میشوند، در حالی که فعالیت به سمت بخشهای خلفی قشر پیشانی جانبی به تدریج به بافتهای موقعیتی ویژه، خاص تر میشود. این سازماندهی سلسله مراتبی نشان میدهد که قشر فرونتوپلار بر پردازش اطلاعات در نواحی خلفی از طریق پیش بینیهای گسترده پشتی تا ناحیه 6 در پایین ترین سطح تأثیر میگذارد. قشر فرونتوپلار در بالاترین سطح پردازش شناختی قرار دارد و ممکن است عملکردهای مهمی مانند تازگی طلبی، چندوظیفه ای و تفکر آینده نگر داشته باشد. چندوظیفه ای مستلزم توانایی در نظر گرفتن اهداف در حین انجام یا پردازش اهداف فرعی ثانویه (“شاخههای شناختی”)، توانایی یادآوری برای انجام یک عمل مورد نظر در آینده در حین درگیر شدن در کار دیگری، و مدیریت موثر زمان در حین جابجایی وظایف است. آینده نگری، تصور آینده یا «فراشناخت» نیازمند نظارت بر واقعیت، درون نگری، خود قضاوتی، و استدلال قیاسی است و متضمن تعامل با اجزای شبکه پیشفرض است، که منطقه فرامودی نهایی را در امتداد گرادیان سازمان قشری تشکیل میدهد. (۱۷)
DECISION-MAKING, MOTIVATION, AND ALLOCATION OF EFFORT
Decision-making is the selection of a particular option among a set of alternatives that are expected to produce different outcomes. This requires assignation of value to the available options and comparison of values and outcomes, and depends on both past experience and current motivational state. Motivation invigorates and guides behaviors toward goals. (40) The successful pursuing of a goal requires cognitive control, especially when it is necessary to overcome distraction or strong habitual responses or when outcomes deviate from expectations. Appropriate cognitive control requires cognitive effort, which allocates cognitive resources, such as working memory, to the pursuit of a goal. These functions reflect the coordinated interaction among multiple cortical areas including the OFC, ventromedial PFC, ACC, and MCC and their interactions with the amygdala, ventral striatum (particularly the nucleus accumbens, NAc), and midbrain dopaminergic system (Figure 42.4) (10,14,15 ,40-44)
تصمیم گیری، انگیزه و تخصیص تلاش
تصمیم گیری عبارت است از انتخاب یک گزینه خاص از میان مجموعه ای از گزینهها که انتظار میرود نتایج متفاوتی به همراه داشته باشد. این امر مستلزم تخصیص ارزش به گزینههای موجود و مقایسه ارزشها و نتایج است و به تجربه گذشته و وضعیت انگیزشی فعلی بستگی دارد. انگیزه باعث تقویت و هدایت رفتارها به سمت اهداف میشود. (۴٠) پیگیری موفقیت آمیز یک هدف مستلزم کنترل شناختی است، به ویژه زمانی که برای غلبه بر حواس پرتی یا پاسخهای قوی عادتی ضروری است یا زمانی که نتایج از انتظارات منحرف میشود. کنترل شناختی مناسب نیاز به تلاش شناختی دارد که منابع شناختی مانند حافظه کاری را برای دستیابی به یک هدف اختصاص میدهد. این عملکردها منعکس کننده تعامل هماهنگ بین چندین نواحی قشر مغز از جمله OFC، PFC شکمی، ACC، و MCC و برهمکنش آنها با آمیگدال، جسم مخطط شکمی (به ویژه هسته اکومبنس، NAc) و سیستم دوپامینرژیک مغز میانی است (شکل 42.4). (۱٠، ۱۴، ۱۵، ۴۴-۴٠)
Orbitofrontal Cortex: Decisions Based on Outcomes
As discussed in chapter 39, the OFC has a major role in assignation of value to available options and decision-making based on comparison of values. (10, 42) The OFC processes sensory stimuli from all modalities to compute their subjective value and reward expectations, in order to estimate the expected outcome that can affect behavioral choices. Some neurons of the medial OFC encode the attributes of potential choices or rewards to be expected, whereas others encode the value of potential outcomes of these choices. Neuroimaging and neuropsychological studies show that activity of the human medial OFC is related with the learning, memory, and monitoring of the reward value of stimuli, including their hedonic quality. (41) Pleasant stimuli or positive rewards activate the medial OFC, which is connected to the pregenual ACC. Activation of these areas correlates linearly with the subjective emotional experience of affective stimuli. Unpleasant stimuli or punishers activate the lateral PFC, which is connected to the anterior MCC. The most anterior portion of the medial OFC, corresponding to the medial frontopolar area 10, is involved in the final binary decision about which among available stimuli is chosen to guide behavior. The classical tests of OFC function are reward devaluation paradigms and stimulus-outcome reversal learning. In reward devaluation tasks, subjects learn two stimuli that predict two different rewards. Then one of these stimuli is devalued due to an aversive outcome. In normal conditions, this change in value in one stimulus immediately leads to the selection of the alternative stimulus. In reversal learning studies, one stimulus is paired with a reward and other is not. When learning has been acquired, the rewarding stimulus switches, so that the reward is now delivered after the previous unrewarded stimulus. In normal conditions, this leads to a reversal of choice. (42)
قشر اوربیتوفرونتال: تصمیم گیری بر اساس نتایج
همانطور که در فصل 39 بحث شد، OFC نقش عمده ای در تعیین ارزش به گزینههای موجود و تصمیم گیری بر اساس مقایسه ارزشها دارد. (۱٠، ۴۲) OFC محرکهای حسی را از همه روشها پردازش میکند تا ارزش ذهنی و انتظارات پاداش را محاسبه کند، تا نتیجه مورد انتظاری را که میتواند بر انتخابهای رفتاری تأثیر بگذارد، برآورد کند. برخی از نورونهای OFC داخلی ویژگیهای انتخابهای بالقوه یا پاداشهای مورد انتظار را رمزگذاری میکنند، در حالی که برخی دیگر ارزش نتایج بالقوه این انتخابها را رمزگذاری میکنند. مطالعات تصویربرداری عصبی و عصب روانشناختی نشان میدهد که فعالیت OFC داخلی انسان با یادگیری، حافظه و نظارت بر ارزش پاداش محرکها، از جمله کیفیت لذتگرایانه آنها مرتبط است. (۴۱) محرکهای خوشایند یا پاداشهای مثبت، OFC میانی را فعال میکنند، که به ACC پیش از تولد مرتبط است. فعالسازی این نواحی بهطور خطی با محرکهای هیجانی تأثیرگذار یا تجربهای تنبیهکننده غیرفعال مرتبط است. PFC جانبی، که به MCC قدامی متصل است، قسمت قدامی OFC میانی، مربوط به ناحیه پیشانی میانی 10، در تصمیم گیری نهایی در مورد اینکه کدام یک از محرکهای موجود برای هدایت رفتار انتخاب میشوند، عبارتند از: پاداش کاهش ارزش در موضوع یادگیری محرکهایی که دو پاداش متفاوت را پیشبینی میکنند، سپس یکی از این محرکها به دلیل یک نتیجه بد، بیارزش میشود. در مطالعات یادگیری معکوس، یک محرک با یک پاداش همراه است و دیگری نه. هنگامیکه یادگیری به دست آمد، محرک پاداش دهنده تغییر میکند، به طوری که پاداش اکنون پس از محرک بدون پاداش قبلی ارائه میشود. در شرایط عادی، این منجر به معکوس شدن انتخاب میشود. (۴۲)

FIGURE 42.4 Areas involved in decision-making and motivation
Assignation of value to stimuli and outcomes, decision-making, cognitive control when it is necessary to overcome distraction, cognitive effort, and motivation reflect the coordinated interaction among multiple cortical and subcortical areas, including the orbitofrontal cortex, ventromedial prefrontal cortex, anterior cingulate cortex, ventral striatum (particularly the nucleus accumbens), and midbrain dopamine neurons.
شکل 42.4 حوزههای درگیر در تصمیم گیری و انگیزه
تخصیص ارزش به محرکها و پیامدها، تصمیم گیری، کنترل شناختی زمانی که برای غلبه بر حواس پرتی ضروری است، تلاش شناختی و انگیزه منعکس کننده تعامل هماهنگ بین چندین نواحی قشری و زیر قشری، از جمله قشر اوربیتوفرونتال، قشر پرهفرونتال شکمی، قشر سینگولیت قدامی، جسم مخطط شکمی (به ویژه هسته اکومبنس) و نورون های دوپامین مغز میانی است.
Cingulate Cortex and Subsystems for Control of Reward-Based and Outcome-Based Behavior
As also discussed in chapter 39, the cingulate cortex harbors at least three subsystems controlling behavior. (43, 44) The ACC has an emotional function and includes a subgenual (subcallosal) region primarily activated by negatively valenced stimuli and memories and a pregenual region activated by positively affective (pleasant) reward values with activation that correlates with the subjective state of pleasure. The activation of the pregenual ACC area reflects emotional awareness. The continuous scaling of value in the medial OFC and pregenual ACC is critical for selecting action outcomes in order to select a specific goal during decision-making. The pregenual ACC is also involved in assessment of the cost of actions. The ventral PCC is engaged in assessment of personal relevance of sensory information. (45) Once the personal relevance is coded in the ventral PCC it becomes available to the subgenual ACC. Personal relevance is also encoded in pregenual ACC area 24, where there is attribution of personal relevance to rewards. The MCC enables reward outcome information from the OFC and ACC to be associated with personally relevant space and action information from the PCC to guide motor behavior. The anterior MCC is involved in cognitive motor control and intentional movement generation, which are linked to its major function in feedback-based action responsiveness to error and reward. (43, 44)
قشر سینگولیت و زیرسیستمهایی برای کنترل رفتار مبتنی بر پاداش و نتیجه
همانطور که در فصل 39 نیز بحث شد، قشر سینگولیت حداقل سه زیرسیستم کنترل کننده رفتار را در خود جای داده است. (۴۴، ۴۳) ACC یک عملکرد عاطفی دارد و شامل یک ناحیه فرعی (سابکالوزال) است که عمدتاً توسط محرکها و خاطرات با ظرفیت منفی فعال میشود و یک ناحیه پیشجنسی فعال میشود که با ارزشهای مثبت فعال با حالتهای عاطفی مثبت مرتبط است. لذت فعال شدن ناحیه ACC قبل از تولد نشان دهنده آگاهی عاطفی است. مقیاس گذاری مداوم ارزش در OFC داخلی و ACC پیش از تولد برای انتخاب نتایج اقدام به منظور انتخاب یک هدف خاص در طول تصمیم گیری بسیار مهم است. ACC پیش از تولد نیز در ارزیابی هزینه اقدامات دخالت دارد. PCC شکمی درگیر ارزیابی ارتباط شخصی اطلاعات حسی است. (۴۵) هنگامی که ارتباط شخصی در PCC شکمی کدگذاری شد، در دسترس ACC فرعی قرار میگیرد. ارتباط شخصی نیز در ناحیه 24 ACC پیش از تولد کدگذاری شده است، جایی که ارتباط شخصی به پاداشها نسبت داده میشود. MCC اطلاعات نتیجه پاداش از OFC و ACC را قادر میسازد تا با فضای شخصی مرتبط و اطلاعات عمل از PCC برای هدایت رفتار موتور مرتبط شود. MCC قدامی در کنترل حرکت شناختی و تولید حرکت عمدی نقش دارد که با عملکرد اصلی آن در واکنش عمل مبتنی بر بازخورد به خطا و پاداش مرتبط است. (۴۴، ۴۳)
Subcortical Components of Motivation: Ventral Striatum and Midbrain Dopaminergic System
The key subcortical components of the motivation system are the ventral striatum, particularly the NAc, and the midbrain dopaminergic neurons. (15) The ventral striatum receives partially overlapping inputs from the OFC, medial PFC, and MCC. The caudal ventromedial PFC projects primarily to the NAC; the OFC and insula to the medial caudate and rostral ventral putamen; and the MCC to the head of the caudate. (46) As mentioned in previous chapters, the NAC consists of a core region and a shell region; the NAC core shares features with other components of the striatum whereas the NAC shell constitutes an interface between the basal ganglia and the emotional limbic subcircuits based on its interaction with the amygdala, hypothalamus, periaqueductal gray, and brainstem autonomic regions. Diffusion tractography shows that the lateral-rostral region of the NAC (putative core) is preferentially connected to other components of the basal ganglia, the frontal pole, OFC, and thalamus; the medial-caudal region of the NAC (putative shell) to the amygdala; and both divisions are connected to the ACC, insula, and hippocampus. (47)
اجزای زیر قشری انگیزه: مخطط شکمی و سیستم دوپامینرژیک مغز میانی
اجزای اصلی زیر قشری سیستم انگیزشی عبارتند از جسم مخطط شکمی، به ویژه NAc، و نورونهای دوپامینرژیک مغز میانی. (۱۵) جسم مخطط شکمی ورودیهای تا حدی همپوشانی را از OFC، PFC داخلی و MCC دریافت میکند. پروژه PFC شکمی میانی دمی عمدتاً به NAC انجام میشود. OFC و اینسولا به دم داخلی و پوتامن شکمی منقاری. و MCC به سر دم. (۴۶) همانطور که در فصلهای قبلی ذکر شد، NAC از یک منطقه هسته و یک منطقه پوسته تشکیل شده است. هسته NAC با سایر اجزای جسم مخطط دارای ویژگیهای مشترک است در حالی که پوسته NAC بر اساس تعامل آن با آمیگدال، هیپوتالاموس، خاکستری اطراف مجرا و مناطق خودمختار ساقه مغز، رابطی بین عقدههای پایه و زیرمدارهای لیمبیک عاطفی ایجاد میکند. tractography انتشار نشان میدهد که منطقه جانبی منقاری NAC (هسته فرضی) ترجیحا به اجزای دیگر از گانگلیون پایه، قطب فرونتال، OFC، و تالاموس متصل است. ناحیه داخلی-دمی NAC (پوسته فرضی) تا آمیگدال؛ و هر دو بخش به ACC، اینسولا و هیپوکامپ متصل هستند. (47)
Projections from the NAC target the ventral pallidum and substantia nigra pars reticulata, which in turn project to the ventral anterior and dorsomedial nuclei of the thalamus; these thalamic nuclei project back to the OFC, ACC, and MCC thus closing the so-called limbic circuit of the basal ganglia. The ventral pallidum and substantia nigra pars reticulata also project to the STN, superior colliculus, and pedunculopontine tegmental nucleus. (48, 49) Core-based circuits may function in action selection, whereas shellbased circuits participate in encoding the reward value and reinforcing effects of rewards. (48, 50) Both the ventral striatum and the MCC receive strong dopaminergic inputs from the ventral tegmental area and substantia nigra pars compacta. (51) The NAC shell primarily receives inputs from midbrain dopaminergic neurons that encode the reward value of stimuli. The dopamine peaks in the striatum promote the selection of the rewarding action.52 Dopaminergic inputs to the NAC thus promote behavioral activation, approach behavior, exertion of effort, and sustained engagement in tasks. (52-54) In contrast, areas of the PFC, including the anterior MCC, receive dopaminergic inputs from midbrain neurons that encode the motivational salience of stimuli and increase their activity both in response to a positive or a negative outcome. These signals are important to allocate cognitive effort. Whereas the dopamine peaks are important signals for rewards, the reduced firing of dopaminergic neurons may not be the major signal encoding rewardprediction errors or punishers. (10) The lateral OFC (areas 47/ 12) contains error neurons that detect nonreward and respond when an expected reward is not obtained or when an action leads to an aversive outcome. These OFC negative reward prediction error neurons may be more suitable to encode negative reward-prediction errors than midbrain dopaminergic neurons, as OFC neurons encode both the expected reward and learned reinforcer (stimulus-reward association). (10) The lateral OFC projects to the anterior MCC and ventrolateral PFC, which are involved in outcomebased action control. (10) The OFC also sends inputs to the ventral striatum, which via the globus pallidus projects to the lateral habenula. The lateral habenula responds to negative outcomes and controls the activity of midbrain dopamine neurons. Thus, the error-encoding neurons of the OFC may have a major role in the mechanism of stimulusoutcome reversal learning, by which a previously rewarding stimulus is no longer chosen when it no longer produces an expected favorable outcome or results in punishment. (42)
پروجکشنهای NAC، پالیدوم شکمی و ماده سیاه پارس رتیکولاتا را هدف قرار میدهند که به نوبه خود به هستههای قدامی شکمی و پشتی میانی تالاموس میرسند. این هستههای تالاموس به OFC، ACC و MCC برمیگردند و بنابراین مدار لیمبیک عقدههای پایه را میبندند. رنگ پری شکمی و ماده سیاه پارس رتیکولاتا نیز به STN، کولیکولوس فوقانی و هسته تگمنتال پدانکولوپونتین پیش میروند. (۴۹، ۴۸) مدارهای مبتنی بر هسته ممکن است در انتخاب کنش عمل کنند، در حالی که مدارهای مبتنی بر پوسته در رمزگذاری ارزش پاداش و تقویت اثرات پاداشها شرکت میکنند. (۵۰، ۴۸) هر دو جسم مخطط شکمی و MCC ورودیهای دوپامینرژیک قوی از ناحیه تگمنتال شکمی و ماده سیاه پارس فشرده دریافت میکنند. (51) پوسته NAC در درجه اول ورودیهایی را از نورونهای دوپامینرژیک مغز میانی دریافت میکند که ارزش پاداش محرکها را رمزگذاری میکند. پیکهای دوپامین در جسم مخطط انتخاب عمل پاداشدهنده را ارتقا میدهند. ورودیهای دوپامینرژیک به NAC بنابراین فعالسازی رفتاری، رفتار رویکرد، تلاش و مشارکت پایدار در تکالیف را ترویج میکنند. (۵۴-۵۲) در مقابل، نواحی PFC، از جمله MCC قدامی، ورودیهای دوپامینرژیک را از نورونهای مغز میانی دریافت میکنند که برجستگی انگیزشی محرکها را رمزگذاری میکنند و فعالیت آنها را هم در پاسخ به یک نتیجه مثبت یا منفی افزایش میدهند. این سیگنالها برای تخصیص تلاش شناختی مهم هستند. در حالی که پیکهای دوپامین سیگنالهای مهمی برای پاداش هستند، کاهش شلیک نورونهای دوپامینرژیک ممکن است سیگنال اصلی کدکننده خطاها یا مجازاتکنندههای پیشبینی پاداش نباشد. (10) OFC جانبی (مناطق 47/12) حاوی نورونهای خطا است که بدون پاداش را تشخیص میدهند و زمانی که پاداش مورد انتظار به دست نمیآید یا زمانی که عملی منجر به یک نتیجه بد میشود، پاسخ میدهند. این نورونهای خطای پیشبینی پاداش منفی OFC ممکن است برای رمزگذاری خطاهای پیشبینی پاداش منفی نسبت به نورونهای دوپامینرژیک مغز میانی مناسبتر باشند، زیرا نورونهای OFC هم پاداش مورد انتظار و هم تقویتکننده آموختهشده (ارتباط محرک-پاداش) را رمزگذاری میکنند. (10) OFC جانبی به MCC قدامی و PFC بطنی جانبی، که در کنترل عمل مبتنی بر نتیجه نقش دارند، می پردازد. (10) OFC همچنین ورودی ها را به مخطط شکمی میفرستد که از طریق گلوبوس پالیدوس به هابنولای جانبی میرسد. هابنولای جانبی به پیامدهای منفی پاسخ میدهد و فعالیت نورونهای دوپامین مغز میانی را کنترل میکند. بنابراین، نورونهای کدگذاری خطای OFC ممکن است نقش عمدهای در مکانیسم یادگیری معکوس نتیجه محرک داشته باشند، بهوسیله آن یک محرک پاداشدهنده قبلی، زمانی که دیگر نتیجه مطلوب مورد انتظار را ایجاد نمیکند یا منجر به تنبیه نمیشود، دیگر انتخاب نمیشود. (42)
Anterior Midcingulate Cortex Use of Outcome Information for Cognitive Control of Effort
The anterior MCC has a major role in feedback-mediated decision-making for reward/approach and fear/avoidance decisions, by providing an interface among reward, decision-making, action selection, motivation, and effort allocation. (55) The anterior MCC is primarily engaged in tasks that demand cognitive control to pursue a goal, especially when it is necessary to overcome distraction or strong habitual automatic responses. (56, 57) The behavioral relevance of incentives, determined both by the expected size of the reward and the effort cost (demands) to obtaining it, drives the allocation of working-memory resources to accomplish a goal. The anterior MCC monitors action-outcome associations to determine benefits and costs and allocate cognitive effort, then recruits the lateral PFC to implement control. (58) The MCC may monitor both the increased costs and incremental progress and then convey the balance between these variables to the midbrain dopaminergic neurons. Whereas tonic dopamine levels in the lateral PFC promote stability of mental representations, phasic dopamine signals render these circuits into a labile state that allows flexible adaptation of working-memory representations. This phasic dopamine release may also encode a reward-minus-effort signal that promotes the allocation of effort by the anterior MCC. In the ventral striatum, dopamine would first select the action sequences for goal-directed behavior and then invigorate action as the goal is reached. (54) The anterior MCC monitors the outcome of a choice, which is critical for behavioral adjustments. (59) Human studies show activation of the anterior MCC when subjects receive negative feedback following inappropriate behaviors. In experimental animals, neurons in the anterior MCC are activated together with those in the lateral habenula in response to adverse outcomes such as reward omission. (59) Inputs from the lateral habenula indirectly inhibit the activity of dopaminergic neurons encoding stimulus value and projecting to the NAC shell, while directly promoting activity of dopaminergic neurons encoding stimulus valence and projecting to the anterior MCC. As the anterior MCC reciprocally projects to the lateral habenula, these two structures can communicate with each other and cooperate to monitor a negative outcome. Whereas the lateral habenula quickly detects the negative outcome of an ongoing action, the anterior MCC stores information about negative past experiences and signals the appropriate behavioral adjustment during the next choice trial. (59)
قشر سینگولیت میانی قدامی استفاده از اطلاعات نتیجه برای کنترل شناختی تلاش
MCC قدامی نقش عمده ای در تصمیم گیری با واسطه بازخورد برای تصمیمات پاداش/رویکرد و ترس/اجتناب، با ایجاد رابطی بین پاداش، تصمیم گیری، انتخاب اقدام، انگیزه و تخصیص تلاش دارد. (55) MCC قدامی اساساً درگیر وظایفی است که نیازمند کنترل شناختی برای تعقیب یک هدف هستند، به ویژه زمانی که برای غلبه بر حواس پرتی یا پاسخ های خودکار عادتی قوی لازم باشد. (56، 57) ارتباط رفتاری انگیزهها، که هم توسط اندازه مورد انتظار پاداش و هم هزینه تلاش (تقاضاها) برای به دست آوردن آن تعیین میشود، تخصیص منابع حافظه کاری را برای دستیابی به یک هدف هدایت میکند. MCC قدامی، ارتباطهای عمل-نتیجه را برای تعیین مزایا و هزینهها و تخصیص تلاش شناختی نظارت میکند، سپس PFC جانبی را برای اجرای کنترل به کار می گیرد. (58) MCC ممکن است هم هزینههای افزایشیافته و هم پیشرفت تدریجی را کنترل کند و سپس تعادل بین این متغیرها را به نورونهای دوپامینرژیک مغز میانی منتقل کند. در حالی که سطوح دوپامین تونیک در PFC جانبی ثبات بازنمایی ذهنی را افزایش میدهد، سیگنالهای دوپامین فازی این مدارها را به یک حالت ناپایدار تبدیل میکنند که امکان سازگاری انعطافپذیر بازنماییهای حافظه کاری را فراهم میکند. این آزادسازی فازی دوپامین همچنین ممکن است یک سیگنال پاداش منهای تلاش را رمزگذاری کند که تخصیص تلاش توسط MCC قدامی را ترویج میکند. در جسم مخطط شکمی، دوپامین ابتدا توالی عمل را برای رفتار هدفمند انتخاب می کند و سپس با رسیدن به هدف، عمل را تقویت می کند. (54) MCC قدامی نتیجه یک انتخاب را که برای تنظیمات رفتاری حیاتی است، نظارت میکند. (59) مطالعات انسانی فعال شدن MCC قدامی را هنگامی که آزمودنیها به دنبال رفتارهای نامناسب بازخورد منفی دریافت می کنند، نشان میدهد. در حیوانات آزمایشی، نورونها در MCC قدامی همراه با نورونهای موجود در هابنولا جانبی در پاسخ به پیامدهای نامطلوب مانند حذف پاداش فعال میشوند. (59) ورودی هابنولا جانبی به طور غیرمستقیم از فعالیت نورونهای دوپامینرژیک که ارزش محرک را کد میکنند و به پوسته NAC پروجکت میکنند، مهار میکند، در حالی که مستقیماً فعالیت نورونهای دوپامینرژیک را ترویج میکند که ظرفیت محرک را کد میکنند و به MCC قدامی پروجکت میکنند. همانطور که MCC قدامی به طور متقابل به هابنولا جانبی می پردازد، این دو ساختار میتوانند با یکدیگر ارتباط برقرار کرده و برای نظارت بر یک نتیجه منفی همکاری کنند. در حالی که هابنولا جانبی به سرعت نتیجه منفی یک عمل در حال انجام را تشخیص می دهد، MCC قدامی اطلاعات مربوط به تجربیات منفی گذشته را ذخیره میکند و تنظیم رفتاری مناسب را در طول آزمایش انتخابی بعدی سیگنال میدهد. (59)
»» ادامه فصل کنترل اجرایی
»» تمامی کتاب

 ورود/ ثبت نام با جیمیل
ورود/ ثبت نام با جیمیل
