علوم اعصاب پروس؛ سیستم دهلیزی و تعادل: نیستاگموس و آزمونهای کالریک با مثالهای بالینی

دعای مطالعه [ نمایش ]
بِسْمِ الله الرَّحْمنِ الرَّحیمِ
اَللّهُمَّ اَخْرِجْنى مِنْ ظُلُماتِ الْوَهْمِ
خدايا مرا بيرون آور از تاريكىهاى وهم،
وَ اَكْرِمْنى بِنُورِ الْفَهْمِ
و به نور فهم گرامى ام بدار،
اَللّهُمَّ افْتَحْ عَلَيْنا اَبْوابَ رَحْمَتِكَ
خدايا درهاى رحمتت را به روى ما بگشا،
وَانْشُرْ عَلَيْنا خَزائِنَ عُلُومِكَ بِرَحْمَتِكَ يا اَرْحَمَ الرّاحِمينَ
و خزانههاى علومت را بر ما باز كن به امید رحمتت اى مهربانترين مهربانان.
کتاب «علوم اعصاب» اثر پروس و همکاران بهعنوان یکی از جامعترین و معتبرترین منابع در حوزه علوم اعصاب (Neuroscience)، همچنان مرجع کلیدی برای درک پیچیدگیهای مغز و سیستم عصبی است. این اثر با بهرهگیری از تازهترین پژوهشها و توضیحات دقیق درباره سازوکارهای عصبی، پلی میان دانش پایه علوم اعصاب و کاربردهای بالینی ایجاد میکند و نقشی بیبدیل در آموزش، پژوهش و ارتقای دانش مغز و اعصاب ایفا مینماید.
ترجمه دقیق و علمی این شاهکار توسط برند علمی «آیندهنگاران مغز» به مدیریت داریوش طاهری، دسترسی فارسیزبانان به مرزهای نوین دانش علوم اعصاب را ممکن ساخته و رسالتی علمی برای ارتقای آموزش، فهم عمیقتر عملکرد مغز و سیستم عصبی و توسعه روشهای نوین در حوزه سلامت عصبی فراهم آورده است.
» کتاب علوم اعصاب پروس
» » فصل ۱۴: سیستم دهلیزی
در حال ویرایش
» Neuroscience; Dale Purves, et al.
»» CHAPTER 14: The Vestibular System

Overview
THE VESTIBULAR SYSTEM PROCESSES sensory information underlying motor responses to (and perceptions of) self-motion, head position, and spatial orientation relative to gravity, helping to stabilize gaze, head, and posture. The peripheral portion of the vestibular system includes inner ear structures that function as an inertial guidance device made up of small linear accelerometers and angular velocity sensors, continuously reporting information about the motions and position of the head to integrative centers in the brainstem, cerebellum, and cerebral cortices. The central portion of the system includes the vestibular nuclei, which make extensive connections with brainstem and cerebellar structures. The vestibular nuclei also innervate motor neurons controlling extraocular, cervical, and postural muscles, thus mediating stabilization of gaze, head orientation, and posture during movement. Interestingly, the vestibular nuclei also receive visual input, resulting in multisensory integration at the earliest point in central vestibular processing. Although we are normally unaware of its functioning, the vestibular system is a key component in postural reflexes and eye movements and, in concert with the visual and proprioceptive systems, plays a central role in distinguishing self-generated “active” movements of the head and body from passive movements resulting from externally applied forces. This multimodal integration is critical to our perception of self-motion, spatial orientation, and body representation. In summary, the vestibular system gives rise to a “sixth sense” that is critical both to automatic behaviors and to perception, with the consequence that balance, gaze stabilization during head movement, and sense of orientation in space are all adversely affected if the system is damaged.
مرور کلی
سیستم دهلیزی اطلاعات حسی زیربنایی پاسخهای حرکتی به (و درک) حرکت خود، موقعیت سر و جهتگیری فضایی نسبت به جاذبه را پردازش میکند و به تثبیت نگاه، سر و وضعیت بدن کمک میکند. بخش محیطی سیستم دهلیزی شامل ساختارهای گوش داخلی است که به عنوان یک دستگاه هدایت اینرسی متشکل از شتابسنجهای خطی کوچک و حسگرهای سرعت زاویهای عمل میکنند و به طور مداوم اطلاعات مربوط به حرکات و موقعیت سر را به مراکز یکپارچهسازی در ساقه مغز، مخچه و قشر مغز گزارش میدهند. بخش مرکزی سیستم شامل هستههای دهلیزی است که ارتباطات گستردهای با ساختارهای ساقه مغز و مخچه برقرار میکنند. هستههای دهلیزی همچنین نورونهای حرکتی کنترلکننده عضلات خارج چشمی، گردنی و وضعیتی را عصبدهی میکنند و در نتیجه واسطه تثبیت نگاه، جهتگیری سر و وضعیت بدن در حین حرکت هستند. جالب توجه است که هستههای دهلیزی نیز ورودی بصری دریافت میکنند که منجر به ادغام چندحسی در اولین نقطه در پردازش دهلیزی مرکزی میشود. اگرچه ما معمولاً از عملکرد آن بیاطلاع هستیم، سیستم دهلیزی یک جزء کلیدی در رفلکسهای وضعیتی و حرکات چشم است و در هماهنگی با سیستمهای بینایی و حس عمقی، نقش محوری در تمایز حرکات “فعال” خودجوش سر و بدن از حرکات غیرفعال ناشی از نیروهای خارجی اعمال شده ایفا میکند. این ادغام چندوجهی برای درک ما از حرکت خود، جهتگیری فضایی و نمایش بدن بسیار مهم است. به طور خلاصه، سیستم دهلیزی باعث ایجاد “حس ششم” میشود که هم برای رفتارهای خودکار و هم برای ادراک حیاتی است، با این نتیجه که تعادل، تثبیت نگاه در حین حرکت سر و حس جهتگیری در فضا در صورت آسیب دیدن این سیستم، همگی تحت تأثیر منفی قرار میگیرند.
The Vestibular Labyrinth
The main peripheral component of the vestibular system is an elaborate set of interconnected chambers the labyrinth that has much in common, and is in fact continuous, with the cochlea (see Chapter 13). Like the cochlea, the labyrinth is derived from the oticplacode of the embryo and uses the same specialized set of sensory cells hair cellsto transduce physical motion into neural impulses. In the cochlea, the motion arises from airborne sound stimuli; in the labyrinth, the pertinent motions arise from the effects of gravity and from translational and rotational movements of the head.
لابیرنت دهلیزی
جزء محیطی اصلی سیستم دهلیزی، مجموعهای پیچیده از محفظههای به هم پیوسته به نام لابیرنت است که اشتراکات زیادی با حلزون گوش دارد و در واقع پیوسته است (به فصل ۱۳ مراجعه کنید). لابیرنت، مانند حلزون گوش، از پلاکود شنوایی جنینی مشتق شده و از همان مجموعه تخصصی سلولهای حسی، سلولهای مویی، برای تبدیل حرکت فیزیکی به تکانههای عصبی استفاده میکند. در حلزون گوش، حرکت از محرکهای صوتی هوابرد ناشی میشود؛ در لابیرنت، حرکات مربوطه از اثرات گرانش و حرکات انتقالی و چرخشی سر ناشی میشوند.
The labyrinth is buried deep in the temporal bone and consists of the two otolith organs the utricle and saccule and three semicircular canals (Figure 14.1). The elaborate architecture of these components explains why this part of the vestibular system is called the labyrinth. The utricle and saccule are specialized primarily to respond to translational movements of the head and static head position relative to the gravitational axis (i.e., head tilts), whereas the semicircular canals, as their shapes suggest, are specialized for responding to rotations of the head.
لابیرنت در اعماق استخوان گیجگاهی قرار دارد و از دو اندام اتولیتی، اوتریکول و ساکول و سه کانال نیمدایرهای، تشکیل شده است (شکل 14.1). معماری پیچیده این اجزا توضیح میدهد که چرا این بخش از سیستم دهلیزی، لابیرنت نامیده میشود. اوتریکول و ساکول در درجه اول برای پاسخ به حرکات انتقالی سر و موقعیت ثابت سر نسبت به محور گرانش (یعنی کج شدن سر) تخصص یافتهاند، در حالی که کانالهای نیمدایرهای، همانطور که شکل آنها نشان میدهد، برای پاسخ به چرخشهای سر تخصص یافتهاند.

FIGURE 14.1 The labyrinth and its innervation. The vestibular and auditory portions of cranial nerve VIII are shown; the small connection from the vestibular nerve to the cochlea contains auditory efferent fibers. General orientation within the head is shown in Figure 13.4; see also Figure 14.8.
شکل ۱۴.۱ لابیرنت و عصبدهی آن. بخشهای دهلیزی و شنوایی عصب جمجمهای VIII نشان داده شده است؛ اتصال کوچک عصب دهلیزی به حلزون گوش حاوی الیاف وابران شنوایی است. جهتگیری کلی در سر در شکل ۱۳.۴ نشان داده شده است؛ همچنین به شکل ۱۴.۸ مراجعه کنید.
The intimate relationship between the cochlea and the labyrinth goes beyond their common embryonic origin. Indeed, the cochlear and vestibular spaces are actually joined (see Figure 14.1), and the specialized ionic environments of the vestibular end organ are much like those of the cochlea. The membranous sacs within the bone are filled with fluid (endolymph) and are collectively called the membranous labyrinth. Between the bony walls (the osseous labyrinth) and the membranous labyrinth is another fluid, the perilymph, which is similar in composition to cerebrospinal fluid (see Chapter 13). The vestibular hair cells are located in the utricle and saccule and in three jug- like swellings called ampullae, located at the base of the semicircular canals next to the utricle. As in the cochlea, tight junctions seal the apical surfaces of the vestibular hair cells, ensuring that endolymph selectively bathes the hair cell bundle while remaining separate from the perilymph surrounding the basal portion of the hair cell.
رابطهی نزدیک بین حلزون گوش و لابیرنت فراتر از منشأ جنینی مشترک آنهاست. در واقع، فضاهای حلزونی و دهلیزی در واقع به هم متصل هستند (شکل 14.1 را ببینید) و محیطهای یونی تخصصی اندام انتهایی دهلیزی بسیار شبیه به محیطهای یونی حلزون گوش هستند. کیسههای غشایی درون استخوان پر از مایع (اندولنف) هستند و در مجموع لابیرنت غشایی نامیده میشوند. بین دیوارههای استخوانی (لابیرنت استخوانی) و لابیرنت غشایی مایع دیگری به نام پریلنف وجود دارد که از نظر ترکیب مشابه مایع مغزی نخاعی است (به فصل 13 مراجعه کنید). سلولهای مویی دهلیزی در اتریکول و ساکول و در سه برآمدگی کوزهمانند به نام آمپول قرار دارند که در پایه مجاری نیمدایرهای کنار اتریکول واقع شدهاند. همانند حلزون گوش، اتصالات محکم، سطوح رأسی سلولهای مویی دهلیزی را میبندند و تضمین میکنند که اندولنف به طور انتخابی دسته سلولهای مویی را در بر میگیرد و در عین حال از پریلنف اطراف قسمت پایه سلول مویی جدا میماند.
Vestibular Hair Cells
The vestibular hair cells, which, like cochlear hair cells, transduce minute displacements into behaviorally relevant receptor potentials, provide the basis for vestibular function. Vestibular and auditory hair cells are quite similar; Chapter 13 gave a detailed description of hair cell structure and function. As in the case of auditory hair cells, movement of the stereocilia toward the kinocilium in the vestibular end organs opens mechanically gated transduction channels located at the tips of the stereocilia, depolarizing the hair cell and causing neurotransmitter release onto (and excitation of) the vestibular nerve fibers (Figure 14.2A,B). Movement of the stereocilia in the direction away from the kinocilium closes the channels, hyperpolarizing the hair cell and thus reducing vestibular nerve activity. The biphasic nature of the receptor potential means that some transduction channels are open in the absence of stimulation, with the result that hair cells tonically release transmitter, thereby generating considerable spontaneous activity in vestibular nerve fibers (see Figure 14.6). One consequence of these spontaneous action potentials is that the firing rates of vestibular fibers can increase or decrease in a manner that faithfully mimics the receptor potentials produced by the hair cells. Adaptation in vestibular hair cells, mediated by calcium entering through mechanoelectrical transduction (MET) and voltage gated calcium channels, is especially important to vestibular function, as it allows hair cells to continue to signal small changes in head position despite much larger tonic forces of gravity.
سلولهای مویی دهلیزی
سلولهای مویی دهلیزی، که مانند سلولهای مویی حلزون، جابجاییهای جزئی را به پتانسیلهای گیرنده مرتبط با رفتار تبدیل میکنند، اساس عملکرد دهلیزی را فراهم میکنند. سلولهای مویی دهلیزی و شنوایی کاملاً مشابه هستند؛ فصل ۱۳ شرح مفصلی از ساختار و عملکرد سلول مویی ارائه داد. همانند سلولهای مویی شنوایی، حرکت استریوسیلیا به سمت کینوسیلیوم در اندامهای انتهایی دهلیزی، کانالهای انتقال مکانیکی دریچهدار واقع در نوک استریوسیلیا را باز میکند، سلول مویی را دپلاریزه میکند و باعث آزادسازی انتقالدهنده عصبی به (و تحریک) فیبرهای عصبی دهلیزی میشود (شکل ۱۴.۲A، B). حرکت استریوسیلیا در جهت دور از کینوسیلیوم، کانالها را میبندد، سلول مویی را هایپرپلاریزه میکند و در نتیجه فعالیت عصب دهلیزی را کاهش میدهد. ماهیت دوفازی پتانسیل گیرنده به این معنی است که برخی از کانالهای انتقال در غیاب تحریک باز هستند، در نتیجه سلولهای مویی به صورت تونیک فرستنده را آزاد میکنند و در نتیجه فعالیت خودبخودی قابل توجهی در فیبرهای عصبی دهلیزی ایجاد میکنند (شکل 14.6 را ببینید). یکی از پیامدهای این پتانسیلهای عمل خودبخودی این است که سرعت شلیک فیبرهای دهلیزی میتواند به گونهای افزایش یا کاهش یابد که پتانسیلهای گیرنده تولید شده توسط سلولهای مویی را به طور دقیق تقلید کند. سازگاری در سلولهای مویی دهلیزی، که با ورود کلسیم از طریق انتقال مکانیکی-الکتریکی (MET) و کانالهای کلسیم وابسته به ولتاژ ایجاد میشود، به ویژه برای عملکرد دهلیزی مهم است، زیرا به سلولهای مویی اجازه میدهد تا علیرغم نیروهای تونیک بسیار بزرگتر گرانش، به ارسال سیگنال تغییرات کوچک در موقعیت سر ادامه دهند.
Importantly, the hair cell bundles in each vestibular organ have specific orientations (Figure 14.2C). As a result, the organ as a whole is responsive to displacements in all directions. In a given semicircular canal, the hair cells in the ampulla are all polarized in the same direction, and the three semicircular canals are oriented to detect rotational motion around the three cardinal (i.e., x, y, and z) axes. In the utricle and saccule, a specialized area called the striola divides the hair cells into two populations having opposing polarities (see Figures 14.2C and 14.4C). The directional polarization of the receptor surfaces is a principle of organization in the vestibular system, as will become apparent in the following descriptions of the individual vestibular organs.
نکته مهم این است که دستههای سلولهای مویی در هر اندام دهلیزی جهتگیریهای خاصی دارند (شکل 14.2C). در نتیجه، کل اندام به جابجاییها در همه جهات واکنش نشان میدهد. در یک مجرای نیمدایرهای مشخص، سلولهای مویی در آمپولا همگی در یک جهت قطبی شدهاند و سه مجرای نیمدایرهای طوری جهتگیری شدهاند که حرکت چرخشی را حول سه محور اصلی (یعنی x، y و z) تشخیص دهند. در اتریکول و ساکول، یک ناحیه تخصصی به نام استریولا، سلولهای مویی را به دو جمعیت با قطبهای متضاد تقسیم میکند (به شکلهای 14.2C و 14.4C مراجعه کنید). قطبش جهتدار سطوح گیرنده، یک اصل سازماندهی در سیستم دهلیزی است، همانطور که در توضیحات بعدی در مورد اندامهای دهلیزی جداگانه آشکار خواهد شد.
Otolith Organs: The Utricle and Saccule
The two otolith organs, the utricle and the saccule, detect tilting and translational (i.e, linear, as opposed to rotational) movements of the head. Both of these organs contain a sensory epithelium, the macula, which consists of hair cells and associated supporting cells. Overlying the hair cells and their hair bundles is a gelatinous layer; above this layer is a fibrous structure, the otolithic membrane, in which are embedded crystals of calcium carbonate called otoconia (Figure 14.3). The crystals give the otolith organs their name (otolith is Greek for “ear stones”). The otoconia make the otolithic membrane heavier than the structures and fluids surrounding it; thus, when the head tilts, gravity causes the membrane to shift relative to the macula (Figure 14.4). The resulting shearing motion between the otolithic membrane and the macula displaces the hair bundles, which are embedded in the lower, gelatinous surface of the membrane. This displacement of the hair bundles generates a receptor potential in the hair cells. A shearing motion between the macula and the otolithic membrane also occurs when the head undergoes translational movements (Figure 14.5); the greater relative mass of the otolithic membrane causes it to lag behind the macula temporarily, leading to transient displacement of the hair bundle.
اندامهای اتولیتی: اوتریکول و ساکول
دو اندام اتولیتی، اوتریکول و ساکول، حرکات کج شدن و انتقالی (یعنی خطی، در مقابل چرخشی) سر را تشخیص میدهند. هر دوی این اندامها حاوی یک اپیتلیوم حسی به نام ماکولا هستند که از سلولهای مویی و سلولهای پشتیبان مرتبط تشکیل شده است. روی سلولهای مویی و دستههای مویی آنها یک لایه ژلاتینی قرار دارد. بالای این لایه یک ساختار فیبری به نام غشای اتولیتی وجود دارد که در آن کریستالهای کربنات کلسیم به نام اتوکونیا تعبیه شدهاند (شکل 14.3). این کریستالها نام اندامهای اتولیتی را به آنها میدهند (اوتولیت در زبان یونانی به معنای “سنگ گوش” است). اتوکونیا غشای اتولیتی را سنگینتر از ساختارها و مایعات اطراف آن میکند. بنابراین، هنگامی که سر کج میشود، گرانش باعث میشود که غشا نسبت به ماکولا جابجا شود (شکل 14.4). حرکت برشی حاصل بین غشای اتولیتیک و ماکولا، دستههای مو را که در سطح ژلاتینی پایینی غشاء قرار دارند، جابجا میکند. این جابجایی دستههای مو، پتانسیل گیرندهای را در سلولهای مو ایجاد میکند. حرکت برشی بین ماکولا و غشای اتولیتیک همچنین زمانی رخ میدهد که سر تحت حرکات انتقالی قرار میگیرد (شکل 14.5)؛ جرم نسبی بیشتر غشای اتولیتیک باعث میشود که به طور موقت از ماکولا عقب بماند و منجر به جابجایی گذرای دسته مو شود.
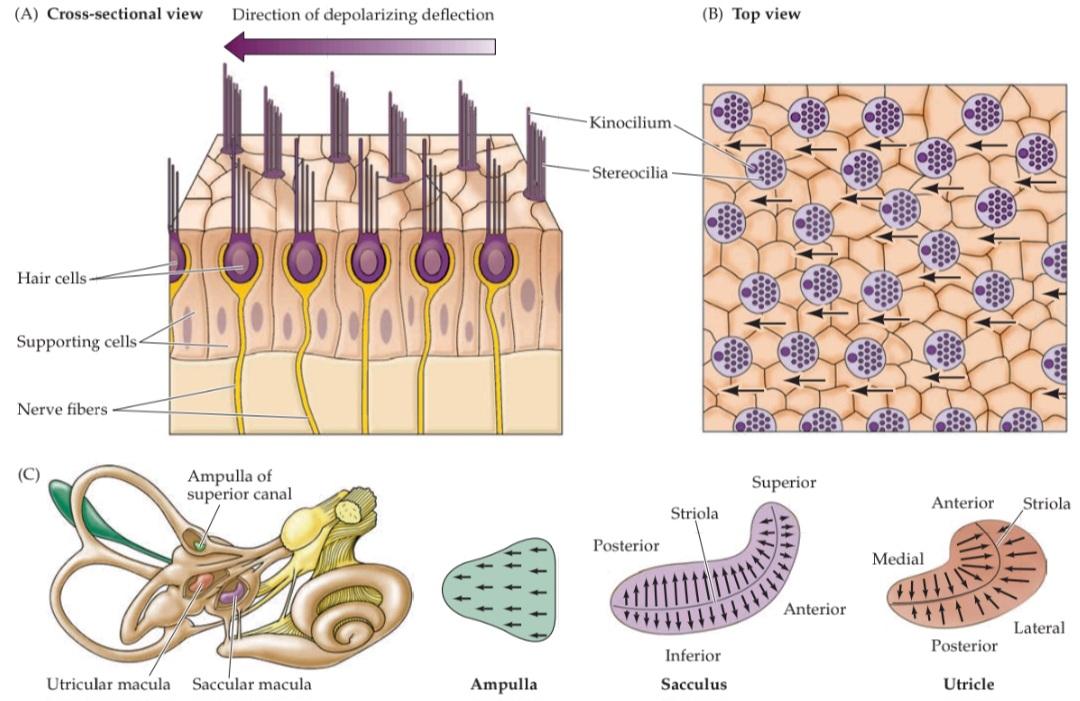
FIGURE 14.2 The morphological polarization of vestibular hair cells and polarization maps of the vestibular organs. (A) A cross section of hair cells shows that the kinocilia of a group of hair cells are all located on the same side of the hair cell. The arrow indicates the direction of deflection that depolarizes the hair cell. (B) View looking down on the hair bundles. (C) In the ampulla located at the base of each semicircular canal, the hair bundles are oriented in the same direction. In the sacculus and utricle, the striola divides the hair cells into populations with opposing hair bundle polarities.
شکل ۱۴.۲ قطبش مورفولوژیکی سلولهای مویی دهلیزی و نقشههای قطبش اندامهای دهلیزی. (الف) برش عرضی سلولهای مویی نشان میدهد که کینوسیلیای یک گروه از سلولهای مویی، همگی در یک طرف سلول مویی قرار دارند. فلش جهت انحرافی را نشان میدهد که سلول مویی را دپلاریزه میکند. (ب) نمای رو به پایین از دستههای مو. (ج) در آمپولا که در پایه هر کانال نیمدایرهای قرار دارد، دستههای مو در یک جهت قرار گرفتهاند. در ساکول و اتریکول، استریولا سلولهای مو را به جمعیتهایی با قطبیتهای دسته مویی مخالف تقسیم میکند.

FIGURE 14.3 Calcium carbonate crystals (otoconia) in the utricular macula of a quail. Each otoconium in this scanning electron micrograph is about 50 μm long. (From Dickman et al., 2004.)
شکل ۱۴.۳ کریستالهای کربنات کلسیم (اتوکونیا) در ماکولای اتریکولار یک بلدرچین. هر اتوکونیوم در این میکروگراف الکترونی روبشی حدود ۵۰ میکرومتر طول دارد. (از دیکمن و همکاران، ۲۰۰۴.)
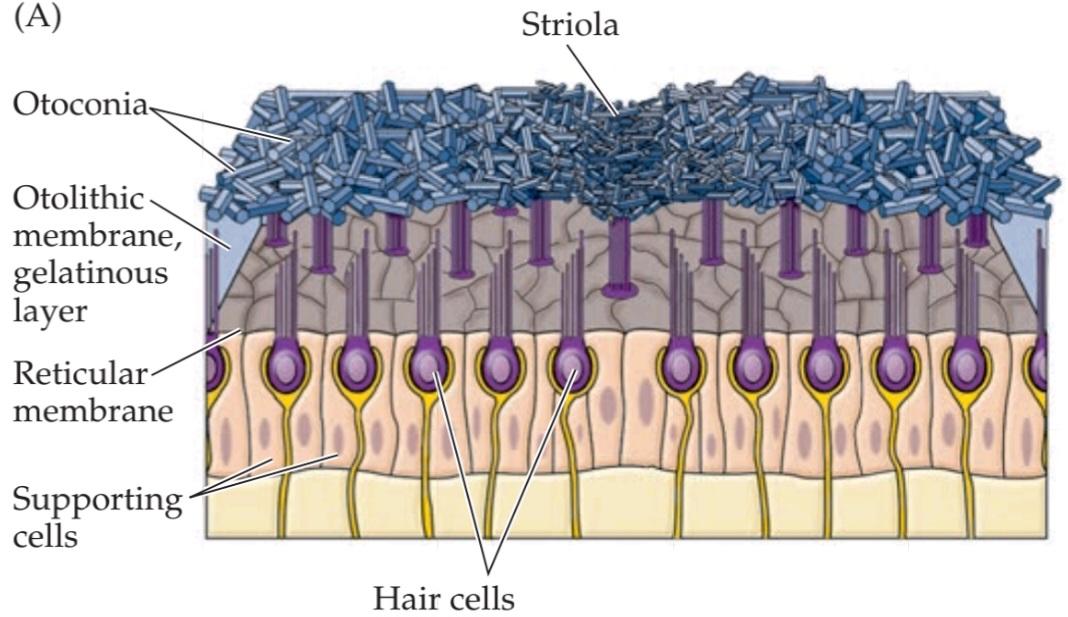

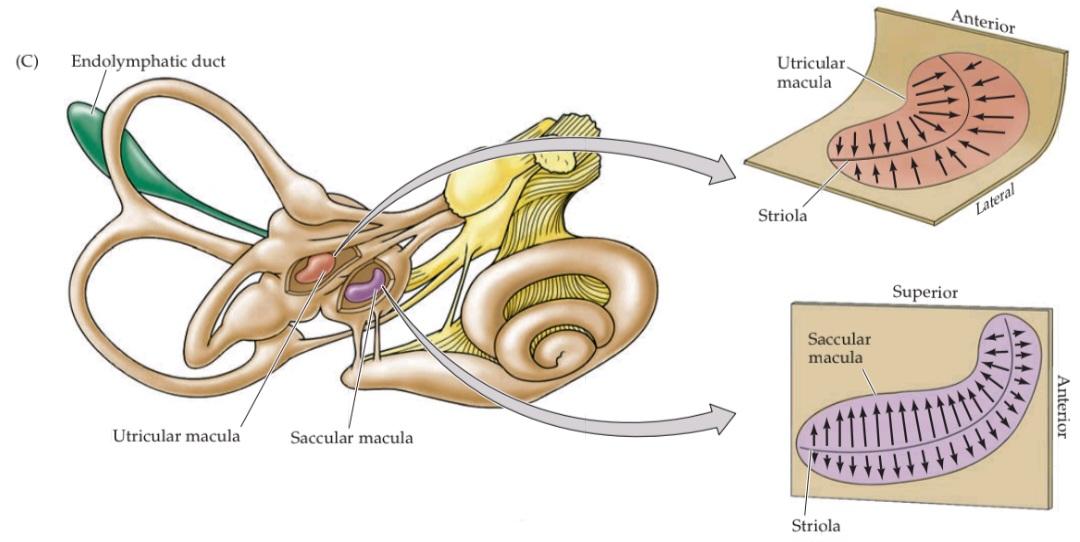
FIGURE 14.4 Morphological polarization of hair cells in the utricular and saccular maculae. (A) Cross section of the utricular macula showing hair bundles projecting into the gelatinous layer when the head is level. (B) Cross section of the utricular macula when the head is tilted. The hair bundles are deflected by the otoconia in the direction of the gravitational force along the macular plane. An equivalent linear acceleration opposite to this force would induce the same deflection of the otoconia and is referred to as the equivalent acceleration. (C) Orientation of the utricular and saccular maculae in the head; arrows show orientation of the kinocilia, as in Figure 14.2. The saccules on either side are oriented more or less vertically, and the utricles more or less horizontally. The striola is a structural landmark consisting of small otoconia arranged in a narrow trench that divides each otolith organ. In the utricular macula, the kinocilia are directed toward the striola. In the saccular macula, the kinocilia point away from the striola. Note that, given the utricle and sacculus on both sides of the body, there is a continuous representation of all directions of head movement.
شکل ۱۴.۴ قطبش مورفولوژیکی سلولهای مویی در ماکولاهای اوتیکولار و ساکولار. (الف) برش عرضی ماکولا اوتیکولار که دستههای مو را نشان میدهد که در حالت سر صاف به داخل لایه ژلاتینی بیرون زدهاند. (ب) برش عرضی ماکولا اوتیکولار هنگام کج شدن سر. دستههای مو توسط اتوکونیا در جهت نیروی گرانش در امتداد صفحه ماکولار منحرف میشوند. یک شتاب خطی معادل در خلاف جهت این نیرو، همان انحراف اتوکونیا را ایجاد میکند و به عنوان شتاب معادل شناخته میشود. (ج) جهتگیری ماکولاهای اوتیکولار و ساکولار در سر؛ فلشها جهتگیری کینوسیلیها را نشان میدهند، همانطور که در شکل 14.2 نشان داده شده است. ساکولها در دو طرف کم و بیش عمودی و اوتیکولها کم و بیش افقی هستند. استرولا یک نقطه عطف ساختاری است که از اتوکونیاهای کوچک تشکیل شده است که در یک شیار باریک قرار گرفتهاند و هر اندام اتولیتی را از هم جدا میکنند. در ماکولای اوتریکولار، کینوسیلیها به سمت استریولا هدایت میشوند. در ماکولای ساکولار، کینوسیلیها از استریولا دور میشوند. توجه داشته باشید که با توجه به اوتریکول و ساکول در دو طرف بدن، نمایش پیوستهای از تمام جهات حرکت سر وجود دارد.
One consequence of the similar effects exerted on otolithic hair cells by certain head tilts and translational movements is that otolith afferents cannot convey information that distinguishes between these two types of stimuli. Consequently, one might expect that these different stimuli would be rendered perceptually equivalent when visual feedback is absent, as occurs in the dark or when the eyes are closed. Nevertheless, blindfolded subjects can discriminate between these two stimulus categories, a feat that depends on the integration of information from the otolith organs and the semicircular canals in the central vestibular system, as described in more detail in the section “Central Pathways for Stabilizing Gaze, Head, and Posture.”
یکی از پیامدهای اثرات مشابهی که بر سلولهای مویی اتولیتی توسط کج کردنهای خاص سر و حرکات انتقالی اعمال میشود، این است که آورانهای اتولیتی نمیتوانند اطلاعاتی را که بین این دو نوع محرک تمایز قائل میشود، منتقل کنند. در نتیجه، میتوان انتظار داشت که این محرکهای مختلف در صورت عدم وجود بازخورد بصری، مانند آنچه در تاریکی یا هنگام بسته بودن چشمها رخ میدهد، از نظر ادراکی معادل باشند. با این وجود، افراد چشمبسته میتوانند بین این دو دسته محرک تمایز قائل شوند، شاهکاری که به ادغام اطلاعات از اندامهای اتولیتی و مجاری نیمدایرهای در سیستم دهلیزی مرکزی بستگی دارد، همانطور که با جزئیات بیشتر در بخش “مسیرهای مرکزی برای تثبیت نگاه، سر و وضعیت بدن” توضیح داده شده است.
As already mentioned, the orientation of the hair cell bundles is organized relative to the striola, which demar- cates the overlying layer of otoconia (see Figure 14.4A). The striola forms an axis of mirror symmetry such that hair cells on opposite sides of the striola have opposing morphological polarizations. Thus, a head tilt along the axis of the striola will excite the hair cells on one side while inhibiting the hair cells on the other side. The saccular macula is oriented vertically and the utricular macula horizontally, with continuous variation in the morphological polarization of the hair cells located in each macula (as shown in Figure 14.4C, where the black arrows indicate the direction of movement that produces excitation). Inspection of the excitatory orientations in the maculae indicates that the utricle responds to translational movements of the head in the horizontal plane and to sideways head tilts, whereas the saccule responds to vertical translational movements of the head and to upward or downward head tilts.
همانطور که قبلاً ذکر شد، جهتگیری دستههای سلولهای مویی نسبت به استرئولا سازماندهی شده است، که لایه پوشاننده اتوکونیا را مشخص میکند (شکل 14.4A را ببینید). استرئولا یک محور تقارن آینهای تشکیل میدهد به طوری که سلولهای مویی در دو طرف مخالف استرئولا، قطبشهای مورفولوژیکی متضادی دارند. بنابراین، کج کردن سر در امتداد محور استرئولا، سلولهای مویی یک طرف را تحریک میکند در حالی که سلولهای مویی طرف دیگر را مهار میکند. ماکولای ساکولار به صورت عمودی و ماکولای اوتریکولار به صورت افقی قرار دارند، با تغییرات مداوم در قطبش مورفولوژیکی سلولهای مویی واقع در هر ماکولا (همانطور که در شکل 14.4C نشان داده شده است، جایی که فلشهای سیاه جهت حرکتی را که باعث تحریک میشود نشان میدهند). بررسی جهتگیریهای تحریکی در ماکولا نشان میدهد که اتریکول به حرکات انتقالی سر در صفحه افقی و کج شدن سر به طرفین پاسخ میدهد، در حالی که ساکول به حرکات انتقالی عمودی سر و کج شدن سر به بالا یا پایین پاسخ میدهد.


FIGURE 14.5 Forces acting on the head displace the otolithic membrane of the utricular macula. For each of the positions and accelerations due to translational movements, some set of hair cells will be maximally excited, while another set will be maximally inhibited. Note that head tilts produce displacements similar to certain accelerations.
شکل ۱۴.۵ نیروهایی که بر سر وارد میشوند، غشای اتولیتیک ماکولای اتریکولار را جابجا میکنند. برای هر یک از موقعیتها و شتابهای ناشی از حرکات انتقالی، مجموعهای از سلولهای مویی به حداکثر تحریک میشوند، در حالی که مجموعهای دیگر به حداکثر مهار میشوند. توجه داشته باشید که کج کردن سر، جابجاییهایی مشابه شتابهای خاص ایجاد میکند.
Note that the saccular and utricular maculae on one side of the head are mirror images of those on the other side. Thus, a tilt of the head to one side has opposite effects on corresponding hair cells of the two utricular maculae. This concept is important in understanding how the central connections of the vestibular periphery mediate the interaction of inputs from the two sides of the head.
توجه داشته باشید که ماکولاهای ساکولار و اتریکولار در یک طرف سر، تصاویر آینهای ماکولاهای طرف دیگر هستند. بنابراین، کج کردن سر به یک طرف، اثرات متضادی بر سلولهای مویی متناظر در دو ماکولا اتریکولار دارد. این مفهوم در درک چگونگی واسطهگری اتصالات مرکزی حاشیه دهلیزی در تعامل ورودیها از دو طرف سر، مهم است.
How Otolith Neurons Sense Head Tilts and Translational Head Movements
The structure of the otolith organs enables them to sense both static displacements, as would be caused by head tilts, and translational components of head movements. Figure 14.5 illustrates some of the forces produced by head tilts and translational movements on the utricular macula.
چگونه نورونهای اتولیت کج شدن سر و حرکات انتقالی سر را حس میکنند
ساختار اندامهای اتولیتی آنها را قادر میسازد تا هم جابجاییهای استاتیک، که در اثر کج شدن سر ایجاد میشوند، و هم اجزای انتقالی حرکات سر را حس کنند. شکل ۱۴.۵ برخی از نیروهای تولید شده توسط کج شدن سر و حرکات انتقالی بر روی ماکولای اتریکولار را نشان میدهد.
The mass of the otolithic membrane relative to the surrounding endolymph, as well as the otolithic membrane’s physical uncoupling from the underlying macula, means that hair bundle displacement will occur transiently in response to translational head movements, and tonically in response to tilting of the head. The resulting hair bundle displacements are reflected in the responses of the vestibular nerve fibers that innervate the otolith organs. As mentioned, these nerve fibers have a steady and relatively high firing rate when the head is upright. Figure 14.6 shows these responses recorded from an otolith afferent fiber, or axon, in a monkey seated in a chair that could be tilted for several seconds to produce a steady force on the head. Prior to the tilt, the axon has a high firing rate, which increases or decreases depending on the direction of the tilt. Notice also that the response remains at a high level as long as the tilting force remains constant; thus, such neurons faithfully encode the static force being applied to the head (see Figure 14.6A). When the head is returned to the original position, the firing level of the neurons returns to baseline value. Conversely, when the tilt is in the opposite direction, the neurons respond by decreasing their firing rate below the resting level (see Figure 14.6B) and remain depressed as long as the static force continues. In a similar fashion, transient increases or decreases in firing rate from spontaneous levels signal the direction of translational movements of the head.
جرم غشای اتولیتی نسبت به اندولنف اطراف، و همچنین جدا شدن فیزیکی غشای اتولیتی از ماکولای زیرین، به این معنی است که جابجایی دسته مو به صورت گذرا در پاسخ به حرکات انتقالی سر و به صورت تونیک در پاسخ به کج کردن سر رخ میدهد. جابجاییهای دسته موی حاصل در پاسخهای فیبرهای عصبی دهلیزی که اندامهای اتولیتی را عصبدهی میکنند، منعکس میشود. همانطور که گفته شد، این فیبرهای عصبی وقتی سر صاف است، سرعت شلیک ثابت و نسبتاً بالایی دارند. شکل 14.6 این پاسخهای ثبت شده از یک فیبر آوران اتولیتی یا آکسون را در یک میمون نشان میدهد که روی صندلی نشسته است که میتواند برای چند ثانیه کج شود تا نیروی ثابتی روی سر ایجاد کند. قبل از کج کردن، آکسون سرعت شلیک بالایی دارد که بسته به جهت کج کردن افزایش یا کاهش مییابد. همچنین توجه داشته باشید که تا زمانی که نیروی کج کردن ثابت بماند، پاسخ در سطح بالایی باقی میماند. بنابراین، چنین نورونهایی به طور دقیق نیروی استاتیک اعمال شده به سر را رمزگذاری میکنند (شکل 14.6A را ببینید). وقتی سر به موقعیت اولیه خود برمیگردد، سطح شلیک نورونها به مقدار پایه برمیگردد. برعکس، وقتی شیب در جهت مخالف باشد، نورونها با کاهش نرخ شلیک خود به زیر سطح استراحت پاسخ میدهند (شکل 14.6B را ببینید) و تا زمانی که نیروی استاتیک ادامه دارد، در حالت انقباض باقی میمانند. به همین ترتیب، افزایش یا کاهش گذرا در نرخ شلیک از سطوح خودبهخودی، جهت حرکات انتقالی سر را نشان میدهد.


FIGURE 14.6 Response of a vestibular nerve axon from an otolith organ. The utricle is the example shown here. (A) The stimulus (top) is a change in head tilt. The spike histogram shows the neuron’s response to tilting in a particular direction. (B) A response of the same fiber to tilting in the opposite direction. (After Fernández and Goldberg, 1976.)
شکل ۱۴.۶ پاسخ آکسون عصب دهلیزی از یک اندام اتولیتی. اتریکول مثالی است که در اینجا نشان داده شده است. (الف) محرک (بالا) تغییر در کج شدن سر است. هیستوگرام اسپایک پاسخ نورون به کج شدن در یک جهت خاص را نشان میدهد. (ب) پاسخ همان فیبر به کج شدن در جهت مخالف. (برگرفته از فرناندز و گلدبرگ، ۱۹۷۶.)
The range of orientations of hair bundles within the otolith organs enables them to transmit information about linear forces in every direction the head might move (see Figure 14.4C). The utricle, which is primarily concerned with motion in the horizontal plane, and the saccule, which is concerned with vertical motion, combine to effectively gauge the linear forces acting on the head at any instant, in three dimensions. Tilts of the head off the horizontal plane and translational movements of the head in any direction stimulate a distinct subset of hair cells in the saccular and utricular maculae, while simultaneously suppressing the responses of other hair cells in these organs. Ultimately, variations in hair cell polarity in the otolith organs produce patterns of vestibular nerve fiber activity that, at a population level, encode head position and the forces that influence it.
طیف جهتگیری دستههای مو در اندامهای اتولیتی، آنها را قادر میسازد تا اطلاعات مربوط به نیروهای خطی را در هر جهتی که سر ممکن است حرکت کند، منتقل کنند (شکل 14.4C را ببینید). اتریکول، که در درجه اول مربوط به حرکت در صفحه افقی است، و ساکول، که مربوط به حرکت عمودی است، با هم ترکیب میشوند تا نیروهای خطی وارد بر سر را در هر لحظه و در سه بعد به طور مؤثر اندازهگیری کنند. کج شدن سر از صفحه افقی و حرکات انتقالی سر در هر جهتی، زیرمجموعه مشخصی از سلولهای مویی را در ماکولهای ساکولی و اتریکولی تحریک میکند، در حالی که همزمان پاسخهای سایر سلولهای مویی در این اندامها را سرکوب میکند. در نهایت، تغییرات در قطبیت سلولهای مویی در اندامهای اتولیتی، الگوهایی از فعالیت فیبر عصبی دهلیزی ایجاد میکند که در سطح جمعیت، موقعیت سر و نیروهایی را که بر آن تأثیر میگذارند، رمزگذاری میکنند.
Semicircular Canals
Whereas the otolith organs are concerned primarily with sensing translational components of head movements and static head tilts, the semicircular canals sense head rotational components of head movements arising either from self induced head rotations or from angular accelerations of the head imparted by external forces, such as a merrygo round. Each of the three semicircular canals has at its base a bulbous expansion the ampulla that houses the sensory epithelium, or crista, that contains the hair cells (Figure 14.7). The structure of the canals suggests how they detect the angular accelerations that arise through rotation of the head. The hair bundles extend out of the crista into a gelatinous mass, the cupula, that bridges the width of the ampulla, forming a viscous barrier through which endolymph cannot circulate. As a result, movements of the endolymphatic fluid distort the relatively compliant cupula. When the head turns in the plane of one of the semicircular canals, the inertia of the endolymph produces a force across the cupula, distending it away from the direction of head movement and causing a displacement of the hair bundles within the crista (Figure 14.8A,B). Note that semircircular canals can be excited by rotations that occur during the initiation of a head tilt as well as rotations resulting from other active or passive movements of the head. In contrast, translational movements of the head produce equal forces on the two sides of the cupula, so the hair bundles within the ampulla are not displaced.
مجاری نیمدایرهای
در حالی که اندامهای اتولیتی در درجه اول با حس کردن اجزای انتقالی حرکات سر و کج شدنهای استاتیک سر سروکار دارند، مجاری نیمدایرهای اجزای چرخشی حرکات سر را که یا از چرخشهای خودبهخودی سر یا از شتابهای زاویهای سر که توسط نیروهای خارجی، مانند چرخش چرخ و فلک، ایجاد میشود، ناشی میشوند، حس میکنند. هر یک از سه مجرای نیمدایرهای در پایه خود دارای یک برآمدگی حبابی شکل به نام آمپولا هستند که اپیتلیوم حسی یا کریستا را در خود جای داده است که حاوی سلولهای مویی است (شکل 14.7). ساختار این مجاری نشان میدهد که چگونه آنها شتابهای زاویهای ناشی از چرخش سر را تشخیص میدهند. دستههای مو از کریستا به داخل یک توده ژلاتینی به نام کوپولا امتداد مییابند که عرض آمپولا را میپوشاند و یک سد چسبناک تشکیل میدهد که از طریق آن اندولنف نمیتواند گردش کند. در نتیجه، حرکات مایع اندولنف، کوپولای نسبتاً سازگار را منحرف میکند. وقتی سر در صفحه یکی از مجاری نیمدایرهای میچرخد، اینرسی اندولنف نیرویی در عرض کوپولا ایجاد میکند که آن را از جهت حرکت سر منبسط کرده و باعث جابجایی دستههای مو در داخل کریستا میشود (شکل 14.8A,B). توجه داشته باشید که مجاری نیمدایرهای میتوانند توسط چرخشهایی که در طول شروع کج کردن سر و همچنین چرخشهای ناشی از سایر حرکات فعال یا غیرفعال سر رخ میدهند، تحریک شوند. در مقابل، حرکات انتقالی سر نیروهای مساوی را در دو طرف کوپولا ایجاد میکنند، بنابراین دستههای مو در داخل آمپول جابجا نمیشوند.

FIGURE 14.7 The ampulla of the posterior semicircular canal. The crista, hair bundles, and cupula are diagrammed. When the head rotates, fluid in the membranous canal distorts the cupula.
شکل ۱۴.۷ آمپول مجرای نیمدایرهای خلفی. کریستا، دستههای مو و کوپولا در نمودار نشان داده شدهاند. هنگامی که سر میچرخد، مایع موجود در مجرای غشایی، کوپولا را کج میکند.
Unlike the saccular and utricular maculae, all of the hair cells in the crista within each semicircular canal are organized with their kinocilia pointing in the same direction (see Figure 14.2B). Thus, when the cupula moves in the appropriate direction, the entire population of hair cells is depolarized and activity in all of the innervating axons increases. When the cupula moves in the opposite direction, the population is hyperpolarized and neuronal activity decreases. Deflections orthogonal to the excitatory-inhibitory direction produce little or no response.
برخلاف ماکولاهای ساکولار و اوتیکولار، تمام سلولهای مویی در کریستا در هر کانال نیمدایرهای با کینوسیلهای خود در یک جهت سازماندهی شدهاند (شکل 14.2B را ببینید). بنابراین، هنگامی که کوپولا در جهت مناسب حرکت میکند، کل جمعیت سلولهای مویی دپلاریزه میشود و فعالیت در تمام آکسونهای عصبدهنده افزایش مییابد. هنگامی که کوپولا در جهت مخالف حرکت میکند، جمعیت هایپرپلاریزه میشود و فعالیت نورونی کاهش مییابد. انحرافات عمود بر جهت تحریکی-مهاری، پاسخ کمی ایجاد میکنند یا اصلاً پاسخی ایجاد نمیکنند.
Each semicircular canal works in concert with the partner located on the other side of the head that has its hair cells aligned oppositely. There are three such pairs: the two (right and left) horizontal canals, and the anterior canal on each side working with the posterior canal on the other side (Figure 14.8C). Head rotation deforms the cupula in opposing directions for the two partners, resulting in opposite changes in their firing rates. Thus, the orientation of the horizontal canals makes them selectively sensitive to rotation in the horizontal plane. More specifically, the hair cells in the canal toward which the head is turning are depolarized, while those on the other side are hyperpolarized.
هر کانال نیمدایرهای با کانال شریکی که در طرف دیگر سر قرار دارد و سلولهای مویی آن در جهت مخالف هم قرار گرفتهاند، هماهنگ عمل میکند. سه جفت از این کانالها وجود دارد: دو کانال افقی (راست و چپ) و کانال قدامی در هر طرف که با کانال خلفی در طرف دیگر کار میکنند (شکل 14.8C). چرخش سر، کوپولا را در جهتهای مخالف برای دو شریک تغییر شکل میدهد و منجر به تغییرات متضاد در نرخ شلیک آنها میشود. بنابراین، جهتگیری کانالهای افقی آنها را به طور انتخابی به چرخش در صفحه افقی حساس میکند. به طور خاصتر، سلولهای مویی در کانالی که سر به سمت آن میچرخد، دپلاریزه میشوند، در حالی که سلولهای مویی در طرف دیگر، هایپرپلاریزه میشوند.



FIGURE 14.8 Functional organization of the semicircular canals. (A) The position of the cupula without angular acceleration. (B) Distortion of the cupula during angular acceleration. When the head is rotated in the plane of the canal (arrow outside canal), the inertia of the endolymph creates a force (arrow inside canal) that displaces the cupula. (C) Arrangement of the canals in pairs. The two horizontal canals form a pair; the right anterior canal (AC) and the left posterior canal (PC) form a pair; and the left AC and the right PC form a pair.
شکل ۱۴.۸ سازماندهی عملکردی مجاری نیمدایرهای. (الف) موقعیت کوپولا بدون شتاب زاویهای. (ب) اعوجاج کوپولا در حین شتاب زاویهای. هنگامی که سر در صفحه کانال میچرخد (فلش خارج کانال)، اینرسی اندولنف نیرویی (فلش داخل کانال) ایجاد میکند که کوپولا را جابجا میکند. (ج) ترتیب کانالها به صورت جفت. دو کانال افقی یک جفت تشکیل میدهند؛ کانال قدامی راست (AC) و کانال خلفی چپ (PC) یک جفت تشکیل میدهند؛ و AC چپ و PC راست یک جفت تشکیل میدهند.
For example, when the head rotates to the left, the cupula is pushed toward the kinocilium in the left horizontal canal, and the firing rate of the relevant axons in the left vestibular nerve increases. In contrast, the cupula in the right horizontal canal is pushed away from the kinocilium, with a concomitant decrease in the firing rate of the related neurons. If the head rotation is to the right, the result is just the opposite. This push pull arrangement operates for all three pairs of canals; the pair whose activity is modulated is in the plane of the rotation, and the member of the pair whose activity is increased is on the side toward which the head is turning. The net result is a system that provides information about the rotation of the head in any direction.
برای مثال، وقتی سر به سمت چپ میچرخد، کوپولا به سمت کینوسیلیوم در کانال افقی چپ رانده میشود و سرعت شلیک آکسونهای مربوطه در عصب دهلیزی چپ افزایش مییابد. در مقابل، کوپولا در کانال افقی راست از کینوسیلیوم دور میشود و همزمان سرعت شلیک نورونهای مربوطه کاهش مییابد. اگر چرخش سر به سمت راست باشد، نتیجه دقیقاً برعکس است. این آرایش فشاری-کششی برای هر سه جفت کانال عمل میکند؛ جفتی که فعالیت آن تعدیل میشود در صفحه چرخش قرار دارد و عضوی از جفت که فعالیت آن افزایش مییابد در سمتی است که سر به سمت آن میچرخد. نتیجه نهایی، سیستمی است که اطلاعاتی در مورد چرخش سر در هر جهتی ارائه میدهد.
How Semicircular Canal Neurons Sense Head Rotations
Like axons that innervate the otolith organs, the vestibular fibers that innervate the semicircular canals exhibit a high level of spontaneous activity. As a result, they can transmit information by either increasing or decreasing their firing rate, thus more effectively encoding rotational head movements (see above). The bidirectional responses of fibers innervating the hair cells of the semicircular canal have been studied by recording the axonal firing rates in a monkey’s vestibular nerve. Seated in a chair, the monkey was rotated continuously in one direction during three phases: an initial period of acceleration, then a period of several seconds at constant velocity, and finally a period of sudden deceleration to a stop (Figure 14.9). The maximum firing rates ob- served correspond to the period of acceleration, when the cupula is deflected; the minimum firing rate corresponds to the period of deceleration, when the cupula is deflected in the opposite direction. During the constant-velocity phase, firing rates return to a baseline level as the cupula returns to its undeflected state over a time course that is related to the cupular elasticity and the viscosity of the endolymph (about 15 seconds). Note that the time it takes the cupula to return to its undistorted state (and for the hair bundles to return to their undeflected position) can occur while the head is still turning, as long as a constant angular velocity is maintained. Such constant forces are rarely found in nature, although they are encountered aboard ships, airplanes, space vehicles, and amusement park rides, where prolonged acceleratory arcs can occur.
چگونه نورونهای کانال نیمدایرهای چرخشهای سر را حس میکنند
مانند آکسونهایی که اندامهای اتولیت را عصبدهی میکنند، فیبرهای دهلیزی که کانالهای نیمدایرهای را عصبدهی میکنند، سطح بالایی از فعالیت خودبهخودی را نشان میدهند. در نتیجه، آنها میتوانند اطلاعات را با افزایش یا کاهش سرعت شلیک خود منتقل کنند و در نتیجه حرکات چرخشی سر را به طور مؤثرتری رمزگذاری کنند (به بالا مراجعه کنید). پاسخهای دو طرفه فیبرهایی که سلولهای مویی کانال نیمدایرهای را عصبدهی میکنند، با ثبت سرعت شلیک آکسونی در عصب دهلیزی یک میمون مورد مطالعه قرار گرفتهاند. میمون که روی صندلی نشسته بود، در طول سه مرحله به طور مداوم در یک جهت چرخانده شد: یک دوره اولیه شتاب، سپس یک دوره چند ثانیهای با سرعت ثابت، و در نهایت یک دوره کاهش سرعت ناگهانی تا توقف (شکل 14.9). حداکثر سرعت شلیک مشاهده شده مربوط به دوره شتاب است، زمانی که کوپولا منحرف میشود؛ حداقل سرعت شلیک مربوط به دوره کاهش سرعت است، زمانی که کوپولا در جهت مخالف منحرف میشود. در طول فاز سرعت ثابت، نرخهای شلیک به سطح پایه بازمیگردند، زیرا کوپولا در طول یک دوره زمانی که به خاصیت ارتجاعی کوپولا و ویسکوزیته اندولنف (حدود ۱۵ ثانیه) مربوط میشود، به حالت بدون انحراف خود بازمیگردد. توجه داشته باشید که مدت زمانی که طول میکشد تا کوپولا به حالت بدون انحراف خود بازگردد (و دستههای مو به موقعیت بدون انحراف خود بازگردند) میتواند در حالی که سر هنوز در حال چرخش است، تا زمانی که سرعت زاویهای ثابت حفظ شود، رخ دهد. چنین نیروهای ثابتی به ندرت در طبیعت یافت میشوند، اگرچه در کشتیها، هواپیماها، وسایل نقلیه فضایی و وسایل بازی پارکهای تفریحی، که در آنها قوسهای شتاب طولانی مدت میتوانند رخ دهند، با آنها مواجه میشویم.

FIGURE 14.9 Response of a vestibular nerve axon from the semicircular canal to angular acceleration. The stimulus (top) is a rotation that first accelerates, then maintains constant velocity, and then decelerates the head. The stimulus evoked change in firing rate of this vestibular unit (bot- tom) reflects the fact that the endolymph has viscosity and inertia and that the cupola has elasticity. Thus, during the initial acceleration, the deflection of the cupula causes the unit activity to rapidly increase. During constant angular velocity, the cupula returns to its undeflected state over a time course related to its elasticity and the viscosity of the fluid, and the unit activity returns to the baseline rate. During deceleration, the cupula is deflected in the opposite direction, causing a transient decrease in the unit firing rate. This behavior can be thought of as the cupula endolymph system dynamic; the inertia of the fluid plays a minor role in this dynamic, coming into play only at very high frequencies of head movement. (After Fernández and Goldberg, 1976.)
شکل ۱۴.۹ پاسخ آکسون عصب دهلیزی از مجرای نیمدایرهای به شتاب زاویهای. محرک (بالا) چرخشی است که ابتدا شتاب میگیرد، سپس سرعت ثابتی را حفظ میکند و سپس سرعت سر را کاهش میدهد. تغییر ناشی از محرک در نرخ شلیک این واحد دهلیزی (پایین) نشان دهنده این واقعیت است که اندولنف دارای ویسکوزیته و اینرسی و کوپولا دارای خاصیت ارتجاعی است. بنابراین، در طول شتاب اولیه، انحراف کوپولا باعث افزایش سریع فعالیت واحد میشود. در طول سرعت زاویهای ثابت، کوپولا در یک دوره زمانی مرتبط با خاصیت ارتجاعی و ویسکوزیته سیال، به حالت بدون انحراف خود بازمیگردد و فعالیت واحد به نرخ پایه بازمیگردد. در طول کاهش سرعت، کوپولا در جهت مخالف منحرف میشود و باعث کاهش گذرا در نرخ شلیک واحد میشود. این رفتار را میتوان به عنوان دینامیک سیستم کوپولا-اندولنف در نظر گرفت. اینرسی سیال نقش جزئی در این دینامیک ایفا میکند و فقط در فرکانسهای بسیار بالای حرکت سر نقش ایفا میکند. (برگرفته از فرناندز و گلدبرگ، ۱۹۷۶)
An interesting aspect of the cupula endolymph system dynamic is that it “smooths” the transduction of head accelerations into neural signals. For example, when the head is angularly accelerated to a constant velocity rather rapidly (corresponding to high frequency rotational movements of the head), vestibular units associated with the affected canal generate a velocity signal; note that the axon firing rate in Figure 14.9 rises linearly during the acceleration phase. However, when the head is moving at a constant angular velocity (i.e., low frequency rotational movements), the rate decays to the spontaneous level (corresponding to an acceleration of zero). This transduction process results in a velocity signal at high frequencies and an acceleration signal at low frequencies, a behavior that can be seen clearly in response to sinusoidal stimuli applied over a wide frequency range.
یکی از جنبههای جالب دینامیک سیستم اندولنف کوپولا این است که تبدیل شتابهای سر به سیگنالهای عصبی را “هموار” میکند. به عنوان مثال، هنگامی که سر به سرعت و با شتاب زاویهای ثابت (مطابق با حرکات چرخشی فرکانس بالای سر) شتاب میگیرد، واحدهای دهلیزی مرتبط با کانال آسیبدیده یک سیگنال سرعت تولید میکنند. توجه داشته باشید که نرخ شلیک آکسون در شکل 14.9 در طول مرحله شتاب به صورت خطی افزایش مییابد. با این حال، هنگامی که سر با سرعت زاویهای ثابت حرکت میکند (یعنی حرکات چرخشی فرکانس پایین)، این نرخ به سطح خودبهخودی (مطابق با شتاب صفر) کاهش مییابد. این فرآیند انتقال منجر به یک سیگنال سرعت در فرکانسهای بالا و یک سیگنال شتاب در فرکانسهای پایین میشود، رفتاری که میتوان آن را به وضوح در پاسخ به محرکهای سینوسی اعمال شده در طیف وسیعی از فرکانس مشاهده کرد.
Central Pathways for Stabilizing Gaze, Head, and Posture
The vestibular system contributes to rapid automatic behaviors, such as reflexive eye movements that stabilize gaze and rapid postural adjustments to maintain balance, and also to higher order processes that are important to our sense of spatial orientation and self motion. The organization of the central vestibular pathways reflects this multifunctional role; these pathways also display two features that distinguish them from the pathways that convey information important to sight, hearing, and touch. First, central vestibular processing is inherently multisensory, because many neurons in the vestibular nuclei the earliest point in central vestibular processingreceive visual input. Second, many neurons in the vestibular nuclei function as premotor neurons in addition to giving rise to ascending sensory projections, providing a very short latency sensorimotor arc that can drive extremely rapid (~10 ms) compensatory eye and head movements in response to vestibular stimulation.
مسیرهای مرکزی برای تثبیت نگاه، سر و وضعیت بدن
سیستم دهلیزی در رفتارهای خودکار سریع، مانند حرکات رفلکسی چشم که نگاه را تثبیت میکنند و تنظیمات سریع وضعیت بدن برای حفظ تعادل، و همچنین در فرآیندهای مرتبه بالاتر که برای حس جهتگیری فضایی و حرکت خود ما مهم هستند، نقش دارد. سازماندهی مسیرهای دهلیزی مرکزی این نقش چندمنظوره را منعکس میکند. این مسیرها همچنین دو ویژگی را نشان میدهند که آنها را از مسیرهایی که اطلاعات مهم برای بینایی، شنوایی و لامسه را منتقل میکنند، متمایز میکند. اول، پردازش دهلیزی مرکزی ذاتاً چندحسی است، زیرا بسیاری از نورونها در هستههای دهلیزی، که اولین نقطه در پردازش دهلیزی مرکزی هستند، ورودی بصری را دریافت میکنند. دوم، بسیاری از نورونها در هستههای دهلیزی علاوه بر ایجاد پیشبینهای حسی صعودی، به عنوان نورونهای پیش حرکتی نیز عمل میکنند و یک قوس حسی-حرکتی با تأخیر بسیار کوتاه فراهم میکنند که میتواند حرکات جبرانی چشم و سر بسیار سریع (حدود 10 میلیثانیه) را در پاسخ به تحریک دهلیزی هدایت کند.
The vestibular end organs communicate, via the vestibular branch of cranial nerve VIII, with targets in the brain stem and the cerebellum that process much of the information necessary to compute head position and motion. As with the cochlear nerve, the vestibular nerves arise from a population of bipolar neurons, the cell bodies of which in this instance reside in the vestibular nerve ganglion (also called Scarpa’s ganglion; see Figure 14.1). The distal processes of these cells innervate the semicircular canals and the otolith organs, while the central processes project via the vestibular portion of the vestibulocochlear nerve (cranial nerve VIII) to the vestibular nuclei (and also directly to the cerebellum; Figure 14.10). Although the canal and otolith afferents are largely segregated in the periphery, a large amount of canalotolith convergence is found in the vestibular nuclei, a feature that ultimately enables the unambiguous encoding of head orientation and motion through the environment. Indeed, although head tilts and translational movements of the head can similarly excite otolith organs, the semicircular canals are excited only by rotations that accompany head tilts and not by purely translational movements. Therefore, integration of information from the otolith organs and semicircular canals in the vestibular nuclei and cerebellum can be used to distinguish head tilts from translational head movements. The vestibular nuclei also integrate a broad range of vestibular and non vestibular information, receiving input from the vestibular nuclei of the opposite side as well as from the cerebellum and the visual and somatosensory systems.
اندامهای انتهایی دهلیزی، از طریق شاخه دهلیزی عصب جمجمهای VIII، با اهدافی در ساقه مغز و مخچه که بخش زیادی از اطلاعات لازم برای محاسبه موقعیت و حرکت سر را پردازش میکنند، ارتباط برقرار میکنند. همانند عصب حلزونی، اعصاب دهلیزی از جمعیتی از نورونهای دوقطبی ناشی میشوند که اجسام سلولی آنها در این مورد در گانگلیون عصب دهلیزی (که گانگلیون اسکارپا نیز نامیده میشود؛ به شکل 14.1 مراجعه کنید) قرار دارند. زوائد دیستال این سلولها، مجاری نیمدایرهای و اندامهای اتولیتی را عصبدهی میکنند، در حالی که زوائد مرکزی از طریق بخش دهلیزی عصب دهلیزی-حلزونی (عصب جمجمهای VIII) به هستههای دهلیزی (و همچنین مستقیماً به مخچه) امتداد مییابند؛ شکل 14.10). اگرچه آورانهای کانال و اتولیت تا حد زیادی در حاشیه از هم جدا هستند، اما مقدار زیادی همگرایی کانال-اتولیتی در هستههای دهلیزی یافت میشود، ویژگیای که در نهایت رمزگذاری بدون ابهام جهتگیری و حرکت سر را در محیط امکانپذیر میکند. در واقع، اگرچه کج شدن سر و حرکات انتقالی سر میتوانند به طور مشابه اندامهای اتولیت را تحریک کنند، اما مجاری نیمدایرهای فقط توسط چرخشهایی که همراه با کج شدن سر هستند تحریک میشوند و نه توسط حرکات صرفاً انتقالی. بنابراین، ادغام اطلاعات از اندامهای اتولیتی و مجاری نیمدایرهای در هستههای دهلیزی و مخچه میتواند برای تشخیص کج شدن سر از حرکات انتقالی سر استفاده شود. هستههای دهلیزی همچنین طیف وسیعی از اطلاعات دهلیزی و غیر دهلیزی را ادغام میکنند و ورودی را از هستههای دهلیزی طرف مقابل و همچنین از مخچه و سیستمهای بینایی و حسی-پیکری دریافت میکنند.
The central projections of the vestibular system participate in two major classes of reflexes: (1) those responsible for maintaining equilibrium and gaze during movement, and (2) those responsible for maintaining posture. The first helps coordinate head and eye movements to keep the gaze fixed on objects of interest during movement. (Other functions include protective or escape reactions; see Box 14A.) The vestibulo ocular reflex (VOR) in particular is a mechanism for producing eye movements that counter head movements, thus permitting the gaze to remain fixed on a particular point (Clinical Applications; see also Chapter 20). For example, activity in the left horizontal canal induced by leftward rotary acceleration of the head excites neurons in the left vestibular nucleus and results in compensatory eye movements to the right.
برآمدگیهای مرکزی سیستم دهلیزی در دو دسته اصلی از رفلکسها شرکت میکنند: (1) آنهایی که مسئول حفظ تعادل و نگاه در حین حرکت هستند، و (2) آنهایی که مسئول حفظ وضعیت بدن هستند. دسته اول به هماهنگی حرکات سر و چشم کمک میکند تا نگاه در حین حرکت روی اشیاء مورد نظر ثابت بماند. (سایر عملکردها شامل واکنشهای محافظتی یا فرار است؛ به کادر 14A مراجعه کنید.) رفلکس دهلیزی-چشمی (VOR) به طور خاص مکانیسمی برای تولید حرکات چشم است که حرکات سر را خنثی میکند، بنابراین اجازه میدهد نگاه روی یک نقطه خاص ثابت بماند (کاربردهای بالینی؛ همچنین به فصل 20 مراجعه کنید). به عنوان مثال، فعالیت در کانال افقی چپ که توسط شتاب چرخشی سر به سمت چپ القا میشود، نورونهای هسته دهلیزی چپ را تحریک میکند و منجر به حرکات جبرانی چشم به سمت راست میشود.
Figure 14.10 illustrates the circuits mediating this reflex. Vestibular nerve fibers originating in the left horizontal semicircular canal project to the medial and superior vestibular nuclei. Excitatory fibers from the medial vestibular nucleus cross to the contralateral abducens nucleus, which has two outputs. One of these is a motor pathway that causes the lateral rectus of the right eye to contract; the other is an excitatory projection that crosses the midline and ascends via the medial longitudinal fasciculus to the left oculomotor nucleus, where it activates neurons that cause the medial rectus of the left eye to contract. Finally, inhibitory neurons project from the medial vestibular nucleus to the left abducens nucleus, directly causing the motor drive on the lateral rectus of the left eye to decrease and also indirectly causing the right medial rectus to relax. The consequence of these several connections is that excitatory input from the horizontal canal on one side produces eye movements toward the opposite side. Therefore, turning the head to the left causes eye movements to the right. In a similar fashion, head turns in other planes activate other semicircular canals, causing other appropriate compensatory eye movements. Thus, the VOR also plays an important role in vertical gaze stabilization in response to the linear vertical head oscillations that accompany locomotion and in response to vertical angular accelerations of the head, as can occur when riding on a swing. Notably, voluntary movements to redirect gaze transiently diminish the VOR, preventing vestibular reflexes from interfering with goal-directed movements. In the clinic, caloric testing provides a useful way to activate the VOR without moving the head, and it is a valuable tool for diagnosing peripheral and central lesions to the vestibular system (see Clinical Applications).
شکل 14.10 مدارهای واسطه این رفلکس را نشان میدهد. فیبرهای عصبی دهلیزی که از کانال نیمدایرهای افقی چپ منشأ میگیرند، به هستههای دهلیزی داخلی و فوقانی امتداد مییابند. فیبرهای تحریکی از هسته دهلیزی داخلی به هسته ابدوسنس مقابل میرسند که دو خروجی دارد. یکی از اینها یک مسیر حرکتی است که باعث انقباض عضله راست جانبی چشم راست میشود؛ دیگری یک برآمدگی تحریکی است که از خط وسط عبور میکند و از طریق فاسیکولوس طولی داخلی به هسته اکولوموتور چپ صعود میکند، جایی که نورونهایی را فعال میکند که باعث انقباض عضله راست داخلی چشم چپ میشوند. در نهایت، نورونهای مهاری از هسته دهلیزی داخلی به هسته ابدوسنس چپ امتداد مییابند و مستقیماً باعث کاهش نیروی محرکه حرکتی عضله راست جانبی چشم چپ میشوند و همچنین به طور غیرمستقیم باعث شل شدن عضله راست داخلی راست میشوند. نتیجه این چندین اتصال این است که ورودی تحریکی از کانال افقی در یک طرف، حرکات چشم را به سمت مقابل ایجاد میکند. بنابراین، چرخاندن سر به سمت چپ باعث حرکات چشم به سمت راست میشود. به همین ترتیب، چرخش سر در صفحات دیگر، کانالهای نیمدایرهای دیگری را فعال میکند و باعث حرکات جبرانی مناسب چشم میشود. بنابراین، VOR همچنین نقش مهمی در تثبیت نگاه عمودی در پاسخ به نوسانات خطی عمودی سر که همراه با حرکت هستند و در پاسخ به شتابهای زاویهای عمودی سر، همانطور که هنگام سوار شدن بر تاب میتواند رخ دهد، ایفا میکند. نکته قابل توجه این است که حرکات ارادی برای تغییر مسیر نگاه، VOR را به طور موقت کاهش میدهند و از تداخل رفلکسهای دهلیزی با حرکات هدفمند جلوگیری میکنند. در کلینیک، آزمایش کالریک روشی مفید برای فعال کردن VOR بدون حرکت سر فراهم میکند و ابزاری ارزشمند برای تشخیص ضایعات محیطی و مرکزی سیستم دهلیزی است (به کاربردهای بالینی مراجعه کنید).

FIGURE 14.10 Connections underlying the vestibulo ocular reflex. Projections of the vestibular nucleus to the nuclei of cranial nerves III (oculomotor) and VI (abducens). The connections to the oculomotor nucleus and to the contralateral abducens nucleus are excitatory (red), whereas the connections to ipsilateral abducens nucleus are inhibitory (black). There are connections from the oculomotor nucleus to the medial rectus of the left eye and from the abducens nucleus to the lateral rectus of the right eye. This circuit moves the eyes to the right-that is, in the direction away from the left horizontal canal-when the head rotates to the left. Turning to the right, which causes increased activity in the right horizontal canal, has the opposite effect on eye movements. The projections from the right vestibular nucleus are omitted for clarity.
شکل ۱۴.۱۰ اتصالات زیربنایی رفلکس دهلیزی-چشمی. برآمدگیهای هسته دهلیزی به هستههای اعصاب جمجمهای III (حرکت چشم) و VI (ربودهکننده). اتصالات به هسته حرکت چشم و هسته حرکت چشم مقابل، تحریکی (قرمز) هستند، در حالی که اتصالات به هسته حرکت چشم همان طرف، مهاری (سیاه) هستند. اتصالاتی از هسته حرکت چشم به عضله راست داخلی چشم چپ و از هسته حرکت چشم به عضله راست خارجی چشم راست وجود دارد. این مدار، چشمها را به سمت راست یعنی در جهت دور از کانال افقی چپ حرکت میدهد، زمانی که سر به سمت چپ میچرخد. چرخش به راست، که باعث افزایش فعالیت در کانال افقی راست میشود، اثر معکوس بر حرکات چشم دارد. برآمدگیهای هسته دهلیزی راست برای وضوح بیشتر حذف شدهاند.
CLINICAL APPLICATIONS
Clinical Evaluation of the Vestibular System
he vestibulo-ocular reflex provides an important means of assessing the and oculomotor nerves and connections between their associated cell bodies in the brainstem. When the head is rotated in the horizontal plane, the vestibular afferents on the side toward the turning motion increase their firing rate, while the afferents on the opposite side decrease their firing rate (Figures A and B). The net difference in firing rates then leads to slow movements of the eyes counter to the turning motion; in a conscious person with normal vestibular function, a fast saccade (see Chapter 20) resets the eye position when the eye reaches its far excursion. This process is referred to as physiological nystagmus, which means “nodding” or oscillatory movements of the eyes (see Figure B1). Physiological nystagmus is an adaptive process that enables the subject to fixate on a visual target despite ongoing rotational movements of the head and body.
کاربردهای بالینی
ارزیابی بالینی سیستم دهلیزی
رفلکس دهلیزی-چشمی وسیله مهمی برای ارزیابی اعصاب و اعصاب حرکتی چشم و ارتباطات بین اجسام سلولی مرتبط با آنها در ساقه مغز فراهم میکند. هنگامی که سر در صفحه افقی چرخانده میشود، آورانهای دهلیزی در سمتی که به سمت حرکت چرخش است، سرعت شلیک خود را افزایش میدهند، در حالی که آورانهای سمت مقابل سرعت شلیک خود را کاهش میدهند (شکلهای A و B). سپس تفاوت خالص در سرعت شلیک منجر به حرکات آهسته چشمها در خلاف جهت حرکت چرخش میشود. در یک فرد هوشیار با عملکرد دهلیزی طبیعی، یک جهش سریع (به فصل 20 مراجعه کنید) موقعیت چشم را هنگامی که چشم به دورترین نقطه خود میرسد، تنظیم مجدد میکند. این فرآیند به عنوان نیستاگموس فیزیولوژیکی شناخته میشود که به معنی “سر تکان دادن” یا حرکات نوسانی چشمها است (به شکل B1 مراجعه کنید). نیستاگموس فیزیولوژیکی یک فرآیند تطبیقی است که فرد را قادر میسازد تا علیرغم حرکات چرخشی مداوم سر و بدن، روی یک هدف بصری متمرکز شود.
Pathologic spontaneous nystagmus can occur if there is unilateral damage to the vestibular system. In this case, the silencing of output from the damaged side results in an abnormal difference in firing rate between the two sides (see Figure B2). This difference causes nystagmus even though no head movements are being made, often resulting in vertigo sufficiently severe to trigger falls and vomiting. Meniere’s disease, named after the nineteenth Century French physician Prosper Meniere, is one cause of acute unilateral vestibular dysfunction and can be particularly disabling for the patient; another is vestibular nerve section, which is often an unavoidable consequence of surgical removal of a vestibular Schwannoma (also known as an acoustic neuroma, a benign tumor of the nerve sheath around the vestibular and acoustic nerves). Interestingly, spontaneous nystagmus slowly diminishes following unilateral vestibular nerve section, presumably because of central compensation and plasticity. In contrast, Meniere’s disease is typically progressive, initially affecting only one ear but slowly encompassing both ears, and is also usually accompanied by tinnitus and a sensorineural hearing deficit in the affected ear(s).
نیستاگموس خودبهخودی پاتولوژیک میتواند در صورت آسیب یکطرفه به سیستم دهلیزی رخ دهد. در این حالت، خاموش شدن خروجی از سمت آسیبدیده منجر به تفاوت غیرطبیعی در میزان شلیک بین دو طرف میشود (شکل B2 را ببینید). این تفاوت باعث نیستاگموس میشود، حتی اگر هیچ حرکتی در سر انجام نشود، که اغلب منجر به سرگیجهای میشود که به اندازه کافی شدید است که باعث افتادن و استفراغ میشود. بیماری منییر، که به نام پزشک فرانسوی قرن نوزدهم، پروسپر منییر، نامگذاری شده است، یکی از علل اختلال عملکرد حاد یکطرفه دهلیزی است و میتواند به ویژه برای بیمار ناتوانکننده باشد. مورد دیگر، قطع عصب دهلیزی است که اغلب نتیجه اجتنابناپذیر برداشتن جراحی شوانوم دهلیزی (همچنین به عنوان نوروم آکوستیک، تومور خوشخیم غلاف عصبی اطراف اعصاب دهلیزی و آکوستیک شناخته میشود) است. جالب توجه است که نیستاگموس خودبهخودی پس از قطع عصب دهلیزی یکطرفه، احتمالاً به دلیل جبران مرکزی و انعطافپذیری، به آرامی کاهش مییابد. در مقابل، بیماری منیر معمولاً پیشرونده است، در ابتدا فقط یک گوش را تحت تأثیر قرار میدهد اما به آرامی هر دو گوش را در بر میگیرد و معمولاً با وزوز گوش و نقص شنوایی حسی-عصبی در گوش (گوشهای) مبتلا همراه است.
(A) View looking down on the top of a person’s head illustrates the fluid motion generated in the left and right horizontal canals and the changes in vestibular nerve firing rates when the head turns to the right.
(الف) نمای رو به پایین از بالای سر یک فرد، حرکت سیال ایجاد شده در کانالهای افقی چپ و راست و تغییرات در نرخ شلیک عصب دهلیزی را هنگام چرخش سر به سمت راست نشان میدهد.

(B) In normal individuals, rotating the head elicits physiological nystagmus (1), which consists of a slow eye movement counter to the direction of head turning. The slow component of the eye movements is due to the net differences in left and right vestibular nerve firing rates acting via the central circuit diagrammed in Figure 14.10. Spontaneous nystagmus (2), where the eyes move rhythmically from side to side in the absence of any head movements, occurs when one of the canals or nerves is damaged. In this situation, net differences in vestibular nerve firing rates exist even when the head is stationary because the vestibular nerve innervating the intact canal fires steadily at rest, in contrast to a lack of activity on the damaged side.
(ب) در افراد طبیعی، چرخاندن سر باعث نیستاگموس فیزیولوژیکی (1) میشود که شامل حرکت آهسته چشم در خلاف جهت چرخش سر است. جزء آهسته حرکات چشم به دلیل تفاوتهای خالص در نرخ شلیک عصب دهلیزی چپ و راست است که از طریق مدار مرکزی نشان داده شده در شکل 14.10 عمل میکنند. نیستاگموس خودبهخودی (2)، که در آن چشمها در غیاب هرگونه حرکت سر، به صورت ریتمیک از یک طرف به طرف دیگر حرکت میکنند، زمانی رخ میدهد که یکی از کانالها یا اعصاب آسیب دیده باشد. در این وضعیت، تفاوتهای خالص در نرخ شلیک عصب دهلیزی حتی زمانی که سر ثابت است نیز وجود دارد زیرا عصب دهلیزی که کانال سالم را عصبدهی میکند، در حالت استراحت به طور پیوسته شلیک میکند، برخلاف عدم فعالیت در سمت آسیبدیده.

Responses to vestibular stimulation are also clinically important because they can be useful in assessing the integrity of the brainstem in unconscious patients. If the individual is placed on his or her back and the head is elevated to about 30° above horizontal, the horizontal canals lie in an almost vertical orientation. Irrigating one ear with cold water will then lead to spontaneous eye movements because convection currents in the canal and direct cooling of the nerve mimic rotational head movements away from the irrigated ear (Figure C). In normal individuals, these eye movements consist of a slow movement toward the irrigated ear and a fast movement away from it. In patients who are comatose due to dysfunction of both cerebral hemispheres but whose brainstem is intact, saccadic movements are no longer made, and the response to cold water consists of only the slow movement component of the eyes toward the side of the irrigated ear (Figure D). In the presence of brainstem lesions involving either the vestibular nuclei themselves, the connections from the vestibular nuclei to oculomotor nuclei (cranial nerves III, IV, or VI), or the peripheral nerves exiting these nuclei, vestibular responses are abolished or altered, depending on the severity of the lesion.
پاسخ به تحریک دهلیزی از نظر بالینی نیز مهم است زیرا میتواند در ارزیابی سلامت ساقه مغز در بیماران بیهوش مفید باشد. اگر فرد به پشت خوابانده شود و سر او حدود 30 درجه بالاتر از سطح افق قرار گیرد، کانالهای افقی تقریباً در جهت عمودی قرار میگیرند. شستشوی یک گوش با آب سرد منجر به حرکات خود به خودی چشم میشود زیرا جریانهای همرفتی در کانال و خنک شدن مستقیم عصب، حرکات چرخشی سر را به سمت خارج از گوش شستشو شده تقلید میکنند (شکل C). در افراد طبیعی، این حرکات چشم شامل یک حرکت آهسته به سمت گوش شستشو شده و یک حرکت سریع به سمت خارج از آن است. در بیمارانی که به دلیل اختلال عملکرد هر دو نیمکره مغزی در کما هستند اما ساقه مغز آنها سالم است، حرکات ساکادیک دیگر انجام نمیشود و پاسخ به آب سرد فقط شامل جزء حرکت آهسته چشمها به سمت کنار گوش شستشو شده است (شکل D). در صورت وجود ضایعات ساقه مغز که خود هستههای دهلیزی، اتصالات هستههای دهلیزی به هستههای حرکتی چشم (اعصاب جمجمهای III، IV یا VI) یا اعصاب محیطی خروجی از این هستهها را درگیر میکند، پاسخهای دهلیزی بسته به شدت ضایعه از بین میروند یا تغییر میکنند.
(C) Caloric testing of vestibular function is possible because irrigating an ear with water slightly warmer than body temperature generates convection currents in the canal that mimic the endolymph movement induced by turning the head to the irrigated side. Irrigation with cold water induces the opposite effect. These currents result in changes in the firing rate of the associated vestibular nerve, with an increased rate on the warmed side and a decreased rate on the chilled side. As in head rotation and spontaneous nystagmus, net differences in firing rates generate eye movements.
(ج) آزمایش کالریک عملکرد دهلیزی امکانپذیر است زیرا شستشوی گوش با آبی که کمی گرمتر از دمای بدن است، جریانهای همرفتی در کانال ایجاد میکند که حرکت اندولنف ناشی از چرخاندن سر به سمت آبیاری شده را تقلید میکنند. آبیاری با آب سرد اثر معکوس دارد. این جریانها منجر به تغییراتی در سرعت شلیک عصب دهلیزی مرتبط میشوند، به طوری که سرعت در سمت گرم افزایش و در سمت سرد کاهش مییابد. همانند چرخش سر و نیستاگموس خودبهخودی، تفاوتهای خالص در سرعت شلیک باعث حرکات چشم میشود.
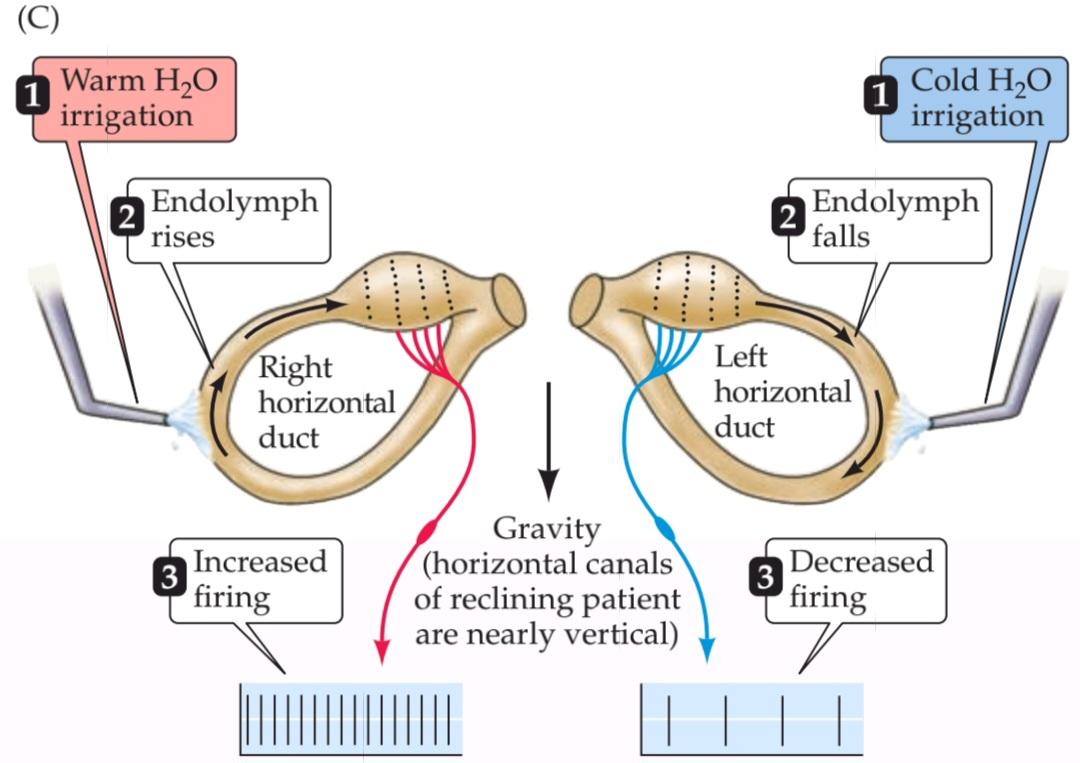
(D) Caloric testing can be used to test the function of the brainstem in an unconscious patient. The figures show eye movements resulting from cold or warm water irrigation in one ear for (1) a normal subject, and in three different conditions in an unconscious patient: (2) with the brainstem intact: (3) with a lesion of the medial longitudinal fasciculus (MLF: note that irrigation in this case results in lateral movement of the eye only on the less active side); and (4) with a low brainstem lesion (see Figure 14.10).
(د) آزمایش کالریک میتواند برای آزمایش عملکرد ساقه مغز در یک بیمار بیهوش استفاده شود. شکلها حرکات چشم ناشی از شستشوی گوش با آب سرد یا گرم را برای (1) یک فرد سالم و در سه حالت مختلف در یک بیمار بیهوش نشان میدهند: (2) با ساقه مغز سالم: (3) با ضایعه فاسیکول طولی داخلی (MLF: توجه داشته باشید که شستشو در این مورد منجر به حرکت جانبی چشم فقط در سمت کمتر فعال میشود)؛ و (4) با ضایعه ساقه مغز در قسمت پایین (شکل 14.10 را ببینید).

BOX 14A Mauthner Cells in Fish
Aprimary function of the vestibular system is to provide information about the direction and speed of ongoing movements, ultimately enabling rapid, coordinated reflexes to compensate for both selfinduced and externally generated forces. One of the most impressive and speediest vestibular mediated reflexes is the tail-flip escape behavior of fish (and larval amphibians), a stereotyped response that allows a potential prey to elude its predators (Figure A; tap on the side of a fish tank if you want to observe the reflex). In response to a perceived risk, fish flick their tail and are thus propelled laterally away from the approaching threat.
جعبه ۱۴A سلولهای ماوتنر در ماهی
عملکرد اصلی سیستم دهلیزی، ارائه اطلاعات در مورد جهت و سرعت حرکات مداوم است که در نهایت رفلکسهای سریع و هماهنگ را برای جبران نیروهای خودالقایی و خارجی فعال میکند. یکی از چشمگیرترین و سریعترین رفلکسهای واسطه دهلیزی، رفتار فرار با چرخاندن دم در ماهی (و لاروهای دوزیست) است، یک پاسخ کلیشهای که به طعمه بالقوه اجازه میدهد از شکارچیان خود فرار کند (شکل A؛ اگر میخواهید رفلکس را مشاهده کنید، به کنار مخزن ماهی ضربه بزنید). در پاسخ به یک خطر درک شده، ماهیها دم خود را تکان میدهند و بنابراین به صورت جانبی از تهدید نزدیک شونده دور میشوند.
The circuitry underlying the tail-flip escape reflex includes a pair of giant medullary neurons called Mauthner cells, their vestibular inputs, and the spinal cord motor neurons to which the Mauthner cells project. (Most fish possess one pair of Mauthner cells in a stereotypic location. Thus, these cells can be consistently visualized and studied from animal to animal.) Movements in the water, such as might be caused by an approaching predator, excite saccular hair cells in the vestibular labyrinth. These receptor potentials are transmitted via the central processes of vestibular ganglion cells in cranial nerve VIII to the two Mauthner cells in the brainstem. As in the vestibulospinal pathway in humans, the Mauthner cells project directly to spinal motor neurons. The small number of synapses intervening between the receptor cells and the motor neurons is one of the ways that this circuit has been optimized for speed by natural selection, an arrangement evident in humans. as well. The large size of the Mauthner axons is another; the axons from these cells in a goldfish are about 50 μm in diameter.
مدار زیربنایی رفلکس فرار با دمپیچ شامل یک جفت نورون غولپیکر مغزی به نام سلولهای ماوتنر، ورودیهای دهلیزی آنها و نورونهای حرکتی نخاع است که سلولهای ماوتنر به آنها متصل میشوند. (بیشتر ماهیها دارای یک جفت سلول ماوتنر در یک مکان کلیشهای هستند. بنابراین، این سلولها را میتوان به طور مداوم از حیوانی به حیوان دیگر تجسم و مطالعه کرد.) حرکات در آب، مانند حرکاتی که ممکن است توسط یک شکارچی در حال نزدیک شدن ایجاد شود، سلولهای مویی ساکولار را در لابیرنت دهلیزی تحریک میکند. این پتانسیلهای گیرنده از طریق فرآیندهای مرکزی سلولهای گانگلیون دهلیزی در عصب جمجمهای VIII به دو سلول ماوتنر در ساقه مغز منتقل میشوند. همانند مسیر دهلیزی-نخاعی در انسان، سلولهای ماوتنر مستقیماً به نورونهای حرکتی نخاعی متصل میشوند. تعداد کم سیناپسهایی که بین سلولهای گیرنده و نورونهای حرکتی قرار دارند، یکی از راههایی است که این مدار توسط انتخاب طبیعی برای سرعت بهینه شده است، ترتیبی که در انسان نیز مشهود است. اندازه بزرگ آکسونهای ماوتنر یکی دیگر از این موارد است؛ آکسونهای این سلولها در ماهی قرمز حدود ۵۰ میکرومتر قطر دارند.
(A) Bird’s-eye view of the sequential body orientations of a fish engaging in a tail-flip escape behavior, with time progressing from left to right. This behavior is mediated largely by vestibular inputs to Mauthner cells. (After Eaton et al., 1977.)
(الف) نمای دید پرنده از جهتگیریهای متوالی بدن یک ماهی که در حال انجام رفتار فرار با چرخاندن دم است، با گذشت زمان از چپ به راست. این رفتار تا حد زیادی توسط ورودیهای دهلیزی به سلولهای ماوتنر انجام میشود. (بعد از Eaton و همکاران، ۱۹۷۷)

(B) Diagram of synaptic events in the Mauthner cells of a fish in response to a disturbance in the water coming from the right. (C) Complementary responses of the right and left Mauthner cells mediating the escape response. Times 1 and 2 correspond to those indicated in Figure B. (After Furshpan and Furukawa, 1962.)
(ب) نمودار رویدادهای سیناپسی در سلولهای ماوتنر یک ماهی در پاسخ به اختلال در آبی که از سمت راست میآید. (ج) پاسخهای مکمل سلولهای ماوتنر راست و چپ که واسطه پاسخ فرار هستند. زمانهای ۱ و ۲ مطابق با زمانهای نشان داده شده در شکل ب هستند. (برگرفته از Furshpan و Furukawa، ۱۹۶۲.)
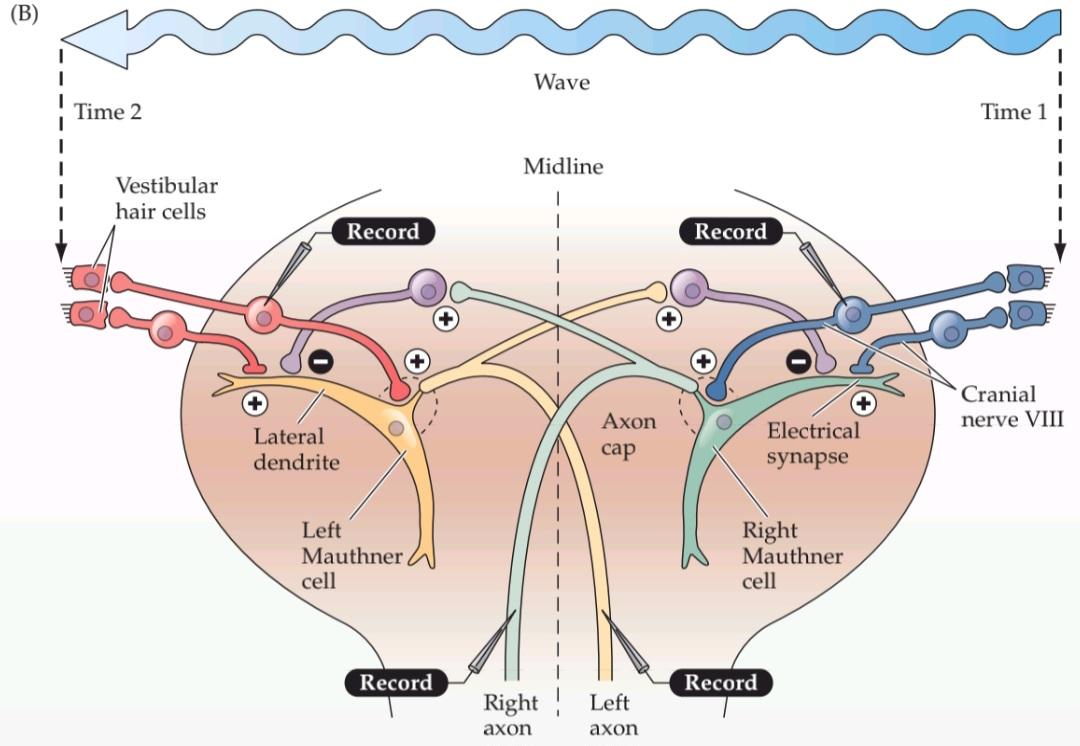
The optimization for speed and direction in the escape reflex also is reflected in the synapses that vestibular nerve afferents make on each Mauthner cell (Figure B). These connections are electrical synapses that allow rapid and faithful transmission of the vestibular signal.
بهینهسازی سرعت و جهت در رفلکس فرار همچنین در سیناپسهایی که آورانهای عصب دهلیزی روی هر سلول ماوتنر ایجاد میکنند، منعکس میشود (شکل B). این اتصالات، سیناپسهای الکتریکی هستند که امکان انتقال سریع و دقیق سیگنال دهلیزی را فراهم میکنند.
An appropriate direction for escape is promoted by two features: (1) each Mauthner cell projects only to contralateral motor neurons; and (2) a local network of bilaterally projecting inter- neurons inhibits activity in the Mauthner cell away from the side on which the vestibular activity originates. In this way. the Mauthner cell on one side faithfully generates action potentials that command contractions of contralateral tail musculature, thus moving the fish out of the path of the oncoming predator. Conversely, the local inhibitory network silences the Mauthner cell on the opposite side during the response (Figure C).
دو ویژگی، جهت مناسب برای فرار را تعیین میکنند: (1) هر سلول ماوتنر فقط به نورونهای حرکتی طرف مقابل سیگنال میدهد؛ و (2) یک شبکه محلی از اینترنورونهای دوطرفه، فعالیت سلول ماوتنر را از طرفی که فعالیت دهلیزی از آن سرچشمه میگیرد، مهار میکند. به این ترتیب، سلول ماوتنر در یک طرف، پتانسیلهای عملی را تولید میکند که انقباضات عضلات دم طرف مقابل را فرمان میدهند و در نتیجه ماهی را از مسیر شکارچیِ در حال نزدیک شدن خارج میکنند. برعکس، شبکه مهاری محلی، سلول ماوتنر در طرف مقابل را در طول پاسخ خاموش میکند (شکل C).
The Mauthner cells in fish are analogous to the reticulospinal and vestibulospinal pathways that control balance, posture, and orienting movements in mammals. The equivalent behavioral responses in humans are evident in a friendly game of tag, or more serious escape endeavors.
سلولهای ماوتنر در ماهیها مشابه مسیرهای رتیکولواسپینال و وستیبولواسپاینال هستند که تعادل، وضعیت بدن و حرکات جهتیابی را در پستانداران کنترل میکنند. پاسخهای رفتاری معادل در انسان در یک بازی دوستانه گرگم به هوا یا تلاشهای جدیتر برای فرار مشهود است.
Loss of the VOR can have severe consequences. A patient with vestibular damage finds it difficult or impossible to fixate on visual targets while the head is moving, a condition called oscillopsia (“bouncing vision”). If the damage is unilateral, the patient usually recovers the ability to fixate objects during head movements. However, a patient with bilateral loss of vestibular function has the persistent and disturbing sense that the world is moving when the head moves. The underlying problem in such cases is that information about head movements normally generated by the vestibular organs is not available to the oculomotor centers, so that compensatory eye movements cannot be made.
از دست دادن VOR میتواند عواقب شدیدی داشته باشد. بیماری که دچار آسیب دهلیزی شده است، در حین حرکت سر، تمرکز روی اهداف بصری را دشوار یا غیرممکن مییابد، وضعیتی که به آن اسیلوپسیا (“دید جهشی”) میگویند. اگر آسیب یک طرفه باشد، بیمار معمولاً توانایی تمرکز روی اشیاء را در حین حرکات سر بازیابی میکند. با این حال، بیماری که دچار از دست دادن دو طرفه عملکرد دهلیزی شده است، این حس مداوم و نگرانکننده را دارد که هنگام حرکت سر، دنیا در حال حرکت است. مشکل اساسی در چنین مواردی این است که اطلاعات مربوط به حرکات سر که معمولاً توسط اندامهای دهلیزی ایجاد میشوند، در دسترس مراکز حرکتی چشم قرار نمیگیرد، به طوری که حرکات جبرانی چشم نمیتوانند انجام شوند.
Descending projections from the vestibular nuclei are essential for postural adjustments of the head, mediated by the vestibulocervical reflex (VCR), and body, mediated by the vestibulospinal reflex (VSR). As with the VOR, these postural reflexes are extremely fast, in part due to the small number of synapses interposed between the vestibular organ and the relevant motor neurons (Box 14A). Like the VOR, the VCR and the VSR are both compromised in patients with bilateral damage to the vestibular periphery. Such patients exhibit diminished head and postural stability, resulting in gait deviations; they also have difficulty balancing. These balance defects become more pronounced in low light or while walking on uneven surfaces, indicating that balance normally is the product of vestibular, visual, and proprioceptive inputs.
برآمدگیهای نزولی از هستههای دهلیزی برای تنظیم وضعیت سر، که توسط رفلکس دهلیزی-گردنی (VCR) واسطهگری میشود، و بدن، که توسط رفلکس دهلیزی-نخاعی (VSR) واسطهگری میشود، ضروری هستند. همانند VOR، این رفلکسهای وضعیتی بسیار سریع هستند، که تا حدی به دلیل تعداد کم سیناپسهای قرار گرفته بین اندام دهلیزی و نورونهای حرکتی مربوطه است (کادر 14A). مانند VOR، VCR و VSR هر دو در بیمارانی که آسیب دو طرفه به حاشیه دهلیزی دارند، دچار اختلال میشوند. چنین بیمارانی کاهش پایداری سر و وضعیت را نشان میدهند که منجر به انحراف در راه رفتن میشود. آنها همچنین در حفظ تعادل مشکل دارند. این نقصهای تعادل در نور کم یا هنگام راه رفتن روی سطوح ناهموار بیشتر نمایان میشوند، که نشان میدهد تعادل به طور معمول محصول ورودیهای دهلیزی، بینایی و حس عمقی است.
The anatomical substrate for the VCR involves the medial vestibular nucleus; axons from this nucleus descend in the medial longitudinal fasciculus to reach the upper cervical levels of the spinal cord (Figure 14.11). This pathway regulates head position by reflex activity of neck muscles in response to stimulation of the semicircular canals caused by rotations of the head. For example, during a downward pitch of the body (e.g., tripping), the superior canals are activated and the head muscles reflexively pull the head up. The dorsal flexion of the head initiates other reflexes, such as forelimb extension and hindlimb flexion, to stabilize the body and protect against a fall (see Chapter 17).
زیرلایه آناتومیکی VCR شامل هسته دهلیزی داخلی است؛ آکسونهای این هسته در دسته طولی داخلی پایین میآیند تا به سطوح فوقانی گردنی نخاع برسند (شکل 14.11). این مسیر با فعالیت رفلکسی عضلات گردن در پاسخ به تحریک مجاری نیمدایرهای ناشی از چرخش سر، موقعیت سر را تنظیم میکند. به عنوان مثال، در طول یک حرکت رو به پایین بدن (مثلاً زمین خوردن)، مجاری فوقانی فعال میشوند و عضلات سر به صورت رفلکسی سر را به سمت بالا میکشند. خم شدن پشتی سر، رفلکسهای دیگری مانند باز شدن اندام جلویی و خم شدن اندام عقبی را برای تثبیت بدن و محافظت در برابر سقوط آغاز میکند (به فصل 17 مراجعه کنید).
The VSR is mediated by a combination of pathways, including the lateral and medial vestibulospinal tracts and the reticulospinal tract. The inputs from the otolith organs project mainly to the lateral vestibular nucleus, which in turn sends axons in the lateral vestibulospinal tract to the ipsilateral ventral horn of the spinal cord (see Figure 14.11). These axons terminate monosynaptically on extensor motor neurons, and they disynaptically inhibit flexor motor neurons; the net result is a powerful excitatory influence on the extensor (antigravity) muscles. When hair cells in the otolith organs are activated, signals reach the medial part of the ventral horn. By activating the ipsilateral pool of motor neurons innervating extensor muscles in the trunk and limbs, this pathway mediates balance and the maintenance of upright posture.
VSR توسط ترکیبی از مسیرها، از جمله مسیرهای دهلیزی-نخاعی جانبی و داخلی و مسیر مشبک-نخاعی، واسطهگری میشود. ورودیهای اندامهای اتولیتی عمدتاً به هسته دهلیزی جانبی هدایت میشوند که به نوبه خود آکسونهایی را در مسیر دهلیزی-نخاعی جانبی به شاخ شکمی همان طرف نخاع میفرستد (شکل 14.11 را ببینید). این آکسونها به صورت تک سیناپسی روی نورونهای حرکتی بازکننده خاتمه مییابند و به صورت غیرسیناپسی نورونهای حرکتی خمکننده را مهار میکنند. نتیجه نهایی، یک تأثیر تحریکی قدرتمند بر عضلات بازکننده (ضد جاذبه) است. هنگامی که سلولهای مویی در اندامهای اتولیتی فعال میشوند، سیگنالها به قسمت داخلی شاخ شکمی میرسند. با فعال کردن مجموعه نورونهای حرکتی همان طرف که عضلات بازکننده را در تنه و اندامها عصبدهی میکنند، این مسیر واسطه تعادل و حفظ وضعیت قائم است.
Decerebrate rigidity, characterized by rigid extension of the limbs, arises when the brainstem is transected above the level of the vestibular nucleus. Decerebrate rigidity in experimental animals is relieved when the vestibular nuclei are lesioned, underscoring the importance of the vestibular system to the maintenance of muscle tone. The tonic activation of extensor muscles in decerebrate rigidity suggests further that descending projections from higher levels of the brain, especially the cerebral cortex, normally suppress the vestibulospinal pathway (see also Chapter 17).
سفتی مخچهای، که با کشش سفت اندامها مشخص میشود، زمانی ایجاد میشود که ساقه مغز بالاتر از سطح هسته دهلیزی قطع شود. سفتی مخچهای در حیوانات آزمایشگاهی با آسیب دیدن هستههای دهلیزی برطرف میشود، که اهمیت سیستم دهلیزی را در حفظ تون عضلانی برجسته میکند. فعال شدن تونیک عضلات بازکننده در سفتی مخچهای بیشتر نشان میدهد که برآمدگیهای نزولی از سطوح بالاتر مغز، به ویژه قشر مغز، معمولاً مسیر دهلیزی-نخاعی را سرکوب میکنند (همچنین به فصل ۱۷ مراجعه کنید).
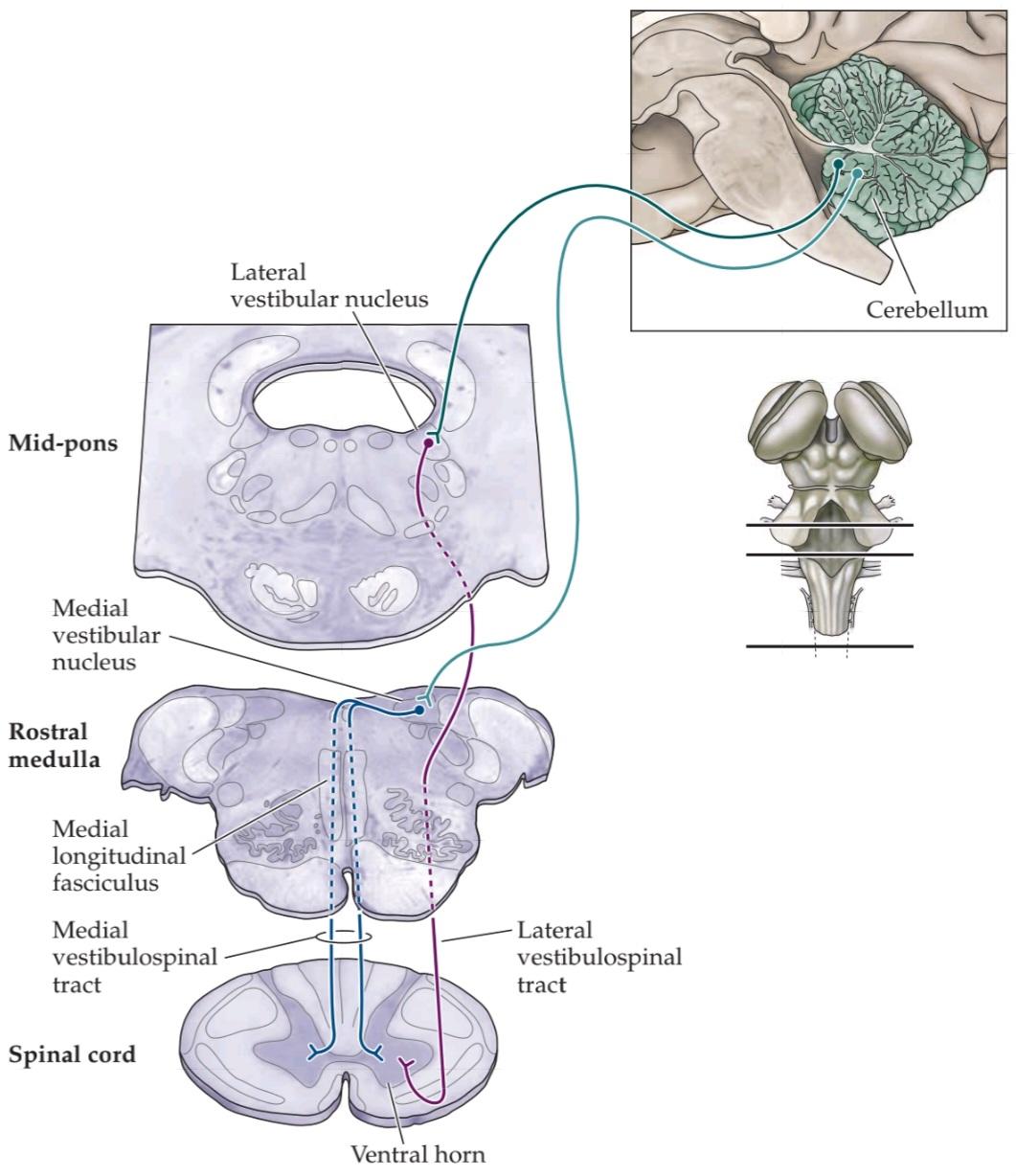
FIGURE 14.11 Descending projections from the medial and lateral vestibular nuclei to the spinal cord underlie the VCR and VSR. The medial vestibular nuclei project bilaterally in the medial longitudinal fasciculus to reach the medial part of the ventral horns and mediate head reflexes in response to activation of semicircular canals. The lateral vestibular nucleus sends axons via the lateral vestibular tract to contact ventral horn cells innervating the axial and proximal limb muscles. Neurons in the lateral vestibular nucleus receive input from the cerebellum, allowing the cerebellum to influence posture and equilibrium.
شکل ۱۴.۱۱ برآمدگیهای نزولی از هستههای دهلیزی داخلی و خارجی به نخاع، زیر VCR و VSR قرار دارند. هستههای دهلیزی داخلی به صورت دو طرفه در دسته طولی داخلی بیرون زدهاند تا به قسمت داخلی شاخهای شکمی برسند و در پاسخ به فعال شدن مجاری نیمدایرهای، رفلکسهای سر را واسطهگری کنند. هسته دهلیزی خارجی، آکسونها را از طریق مسیر دهلیزی خارجی به سلولهای شاخ شکمی که عضلات اندامهای محوری و پروگزیمال را عصبدهی میکنند، میفرستد. نورونهای هسته دهلیزی خارجی، ورودیها را از مخچه دریافت میکنند و به مخچه اجازه میدهند تا بر وضعیت بدن و تعادل تأثیر بگذارد.
Vestibular Cerebellar Pathways
The cerebellum is a major target of ascending vestibular pathways and also provides descending input to the vestibular nuclei, resulting in a recurrent circuit architecture that plays an important role in modulating vestibular activity. These vestibular cerebellar circuits play a critical role in integrating and modulating vestibular signals to enable adaptive changes to the VOR, distinguish head tilts from translational movements, and distinguish passive movements of the head and body from those that are self generated. Major vestibular targets in the cerebellum include the flocculus, paraflocculus, nodulus, uvula, and rostral fastigial nucleus, all of which play distinct roles in vestibular plasticity and multimodal integration.
مسیرهای دهلیزی-مخچهای
مخچه هدف اصلی مسیرهای دهلیزی صعودی است و همچنین ورودی نزولی به هستههای دهلیزی ارائه میدهد که منجر به معماری مدار بازگشتی میشود که نقش مهمی در تعدیل فعالیت دهلیزی ایفا میکند. این مدارهای مخچهای دهلیزی نقش مهمی در ادغام و تعدیل سیگنالهای دهلیزی دارند تا تغییرات تطبیقی در VOR را ممکن سازند، کج شدن سر را از حرکات انتقالی تشخیص دهند و حرکات غیرفعال سر و بدن را از حرکاتی که خود به خود ایجاد میشوند، تشخیص دهند. اهداف اصلی دهلیزی در مخچه شامل فلوکولوس، پارافلوکولوس، ندولوس، زبان کوچک و هسته فاستیژیال روسترال هستند که همگی نقشهای متمایزی در انعطافپذیری دهلیزی و ادغام چندوجهی دارند.
The VOR displays remarkable plasticity in response to altered visual or vestibular input, a feature that has ad- vanced the VOR as a major subject of sensorimotor learning studies. This plasticity manifests after unilateral damage to the vestibular periphery and can also be triggered experimentally by placing optics over the eyes that magnify, miniaturize, or even invert the optical image on the retina. Subjects wearing such optics adapt by changing the gain of the VOR to maintain a stable image on the retina and enable coordinated movements under visual control. In a famous series of experiments Ivo Kohler conducted in the 1950s, he found that after wearing inverting prisms for a week he could function well enough to ride a bicycle down a busy street. The flocculus and paraflocculus contain Purkinje cells that transmit inhibitory signals to the vestibular nucleus, and these neurons change their firing properties in a manner that accounts for adaptation of the VOR. Although most people do not elect to wear prism glasses except for experiments, VOR plasticity comes into play during development, when the rapid growth of the head changes the transform between head and eye movements, and also when people are fitted with corrective lenses.
VOR در پاسخ به ورودیهای بینایی یا دهلیزی تغییر یافته، انعطافپذیری قابل توجهی نشان میدهد، ویژگیای که VOR را به عنوان یک موضوع اصلی مطالعات یادگیری حسی-حرکتی ارتقا داده است. این انعطافپذیری پس از آسیب یک طرفه به حاشیه دهلیزی آشکار میشود و همچنین میتواند به صورت تجربی با قرار دادن اپتیکهایی روی چشمها که تصویر نوری روی شبکیه را بزرگ، کوچک یا حتی معکوس میکنند، فعال شود. افرادی که چنین اپتیکهایی را میپوشند، با تغییر بهره VOR برای حفظ تصویر پایدار روی شبکیه و فعال کردن حرکات هماهنگ تحت کنترل بینایی، سازگار میشوند. در یک سری آزمایشهای معروف که ایوو کوهلر در دهه 1950 انجام داد، دریافت که پس از پوشیدن منشورهای معکوس به مدت یک هفته، میتواند به اندازه کافی خوب عمل کند تا در یک خیابان شلوغ دوچرخهسواری کند. فلوکولوس و پارافلوکولوس حاوی سلولهای پورکنژ هستند که سیگنالهای مهاری را به هسته دهلیزی منتقل میکنند و این نورونها خواص شلیک خود را به گونهای تغییر میدهند که سازگاری VOR را توجیه میکند. اگرچه اکثر افراد به جز در آزمایشها، عینکهای منشوری را انتخاب نمیکنند، اما انعطافپذیری VOR در طول رشد، زمانی که رشد سریع سر، تبدیل بین حرکات سر و چشم را تغییر میدهد، و همچنین زمانی که افراد از لنزهای اصلاحی استفاده میکنند، نقش خود را ایفا میکند.
A previously mentioned, integration of signals from the otolith organs and semicircular canals is needed to disambiguate head tilts from purely translational movements. Recordings made in the nodulus and uvula reveal that individual Purkinje cells integrate signals from these two vestibular sources to unambiguously encode either head tilts or translational movements, suggesting that the nodulus and uvula are critical sites of computation for making this distinction. Another major function of vestibular cerebellar circuitry is helping distinguish vestibular signals that arise from self generated motion from those that are triggered by external forces. An interesting feature of neurons in the rostral fastigial nucleus is that they do not respond to self-generated head or body movements, despite receiving both vestibular and proprioceptive signals from the head and body. An influential idea to account for this specialized responsiveness is that predictive signals generated in the cerebellum cancel out vestibular and proprioceptive signals in rostral fastigial neurons generated from selfmotion, helping distinguish active from passive movements of the head.
همانطور که قبلاً ذکر شد، ادغام سیگنالهای اندامهای اتولیتی و مجاری نیمدایرهای برای تفکیک کج شدن سر از حرکات صرفاً انتقالی مورد نیاز است. ثبتهای انجام شده در گره و زبان کوچک نشان میدهد که سلولهای پورکنژ منفرد سیگنالهای این دو منبع دهلیزی را ادغام میکنند تا به طور واضح کج شدن سر یا حرکات انتقالی را رمزگذاری کنند، که نشان میدهد گره و زبان کوچک مکانهای حیاتی محاسبه برای ایجاد این تمایز هستند. یکی دیگر از عملکردهای اصلی مدار مخچه دهلیزی، کمک به تشخیص سیگنالهای دهلیزی است که از حرکت خود تولید شده از سیگنالهایی که توسط نیروهای خارجی ایجاد میشوند، ناشی میشوند. یکی از ویژگیهای جالب نورونها در هسته فاستیژیال روسترال این است که آنها با وجود دریافت سیگنالهای دهلیزی و حس عمقی از سر و بدن، به حرکات خود تولید شده سر یا بدن پاسخ نمیدهند. یک ایده تأثیرگذار برای توضیح این پاسخگویی تخصصی این است که سیگنالهای پیشبینیکننده تولید شده در مخچه، سیگنالهای دهلیزی و حس عمقی در نورونهای فاستیژیال روسترال تولید شده از خودحرکتی را خنثی میکنند و به تشخیص حرکات فعال از غیرفعال سر کمک میکنند.

FIGURE 14.12 Thalamocortical pathways carrying vestibular information. Unlike somatosensory, visual, and auditory systems, there is no single, canonical “primary vestibular cortex. Rather, there is a “vestibular cortical system” involving a distributed set of cortical areas in the parietal and posterior insular regions (purple ovals). Each of these areas contains neurons that are modulated by vestibular signals, interconnected across areas, and give rise to subcortical connections with the vestibular nuclear complex of the brainstem. Of particular importance is the parietoinsular vestibular cortex, which integrates multimodal proprioceptive signals and generates a “head inspace” frame for body orientation and motor control. Other areas contributing to this vestib- ular cortical system include the ventral premotor cortex and the cingulate motor area (not shown; see Chapter 17).
شکل ۱۴.۱۲ مسیرهای تالاموکورتیکال حامل اطلاعات دهلیزی. برخلاف سیستمهای حسی-پیکری، بینایی و شنوایی، هیچ «قشر دهلیزی اولیه» واحد و متعارفی وجود ندارد. در عوض، یک «سیستم قشر دهلیزی» وجود دارد که شامل مجموعهای توزیعشده از نواحی قشری در نواحی جداری و خلفی اینسولار (بیضیهای بنفش) است. هر یک از این نواحی حاوی نورونهایی است که توسط سیگنالهای دهلیزی تعدیل میشوند، در نواحی مختلف به هم متصل میشوند و باعث ایجاد ارتباطات زیرقشری با کمپلکس هستهای دهلیزی ساقه مغز میشوند. قشر دهلیزی جداری-اینسولار از اهمیت ویژهای برخوردار است که سیگنالهای حس عمقی چندوجهی را ادغام میکند و یک قاب «فضای سر» برای جهتگیری بدن و کنترل حرکتی ایجاد میکند. سایر نواحی مؤثر در این سیستم قشر دهلیزی شامل قشر پیشحرکتی شکمی و ناحیه حرکتی کمربندی است (نشان داده نشده است؛ به فصل ۱۷ مراجعه کنید).
Vestibular Pathways to the Thalamus and Cortex
In addition to the several projections of the vestibular nuclei already mentioned, the superior and lateral vestibular nuclei send axons to the ventral posterior nuclear complex of the thalamus. This, in turn, projects to a variety of cortical areas relevant to the perceptions arising from the processing of vestibular information, including Brodmann’s area 2v just posterior of the face representation in the somatosensory cortex, two regions in Brodmann’s area 3a in the fundus of the central sulcus, and the parietoinsular vestibular cortex (PIVC), which may be especially important to our sense of self motion and orientation in space (Figure 14.12). Indeed, in the 1950s Wilder Penfield found that electrically stimulating the PIVC could elicit strong vestibular sensations, and more modern imaging studies indicate that this region is activated by vestibular stimulation. Electrophysiological studies of individual neurons in these various cortical areas show that the relevant cells respond to proprioceptive and visual stimuli as well as to vestibular stimuli, reflecting the multisensory nature of central vestibular processing. Many of these cortical neurons are activated by moving visual stimuli as well as by rotation of the body (even with the eyes closed), suggesting that these cortical regions are involved in the perception of body orientation in extrapersonal space. Consistent with this interpretation, patients with lesions of the right parietal cortex, including the PIVC, suffer altered perception of personal and extrapersonal space, as discussed in greater detail in Chapter 26.
مسیرهای دهلیزی به تالاموس و قشر مغز
علاوه بر چندین برآمدگی هستههای دهلیزی که قبلاً ذکر شد، هستههای دهلیزی فوقانی و جانبی، آکسونهایی را به کمپلکس هستهای خلفی شکمی تالاموس میفرستند. این به نوبه خود، به نواحی قشری متنوعی مرتبط با ادراکات ناشی از پردازش اطلاعات دهلیزی، از جمله ناحیه 2v برودمن درست در خلف بازنمایی چهره در قشر حسی-پیکری، دو ناحیه در ناحیه 3a برودمن در فوندوس شیار مرکزی و قشر دهلیزی جداری-اینسولار (PIVC) که ممکن است به ویژه برای حس حرکت خود و جهتیابی ما در فضا مهم باشد، ارسال میکند (شکل 14.12). در واقع، در دهه 1950، وایلدر پنفیلد دریافت که تحریک الکتریکی PIVC میتواند احساسات دهلیزی قوی ایجاد کند و مطالعات تصویربرداری مدرنتر نشان میدهد که این ناحیه توسط تحریک دهلیزی فعال میشود. مطالعات الکتروفیزیولوژیک نورونهای منفرد در این نواحی مختلف قشر مغز نشان میدهد که سلولهای مربوطه به محرکهای حس عمقی و بینایی و همچنین محرکهای دهلیزی پاسخ میدهند که منعکسکننده ماهیت چندحسی پردازش دهلیزی مرکزی است. بسیاری از این نورونهای قشر مغز با حرکت محرکهای بینایی و همچنین چرخش بدن (حتی با چشمان بسته) فعال میشوند، که نشان میدهد این نواحی قشر مغز در درک جهتگیری بدن در فضای فراشخصی نقش دارند. مطابق با این تفسیر، بیماران مبتلا به ضایعات قشر جداری راست، از جمله PIVC، از درک تغییر یافته از فضای شخصی و فراشخصی رنج میبرند، همانطور که با جزئیات بیشتر در فصل 26 مورد بحث قرار گرفته است.
Spatial Orientation Perception and Multisensory Integration
Although the vestibular system contributes to many automatic reflexes and most of us remain unaware of its functioning unless it is damaged, it also plays an important role in our perception of spatial orientation and self motion. Indeed, the vestibular system underlies our ability to detect the direction and magnitude of self generated motion and distinguish this motion from the movement of objects around us. Working in concert with the visual system, the vestibular system provides critical information for spatial orientation and navigation.
ادراک جهتیابی فضایی و ادغام چندحسی
اگرچه سیستم دهلیزی در بسیاری از رفلکسهای خودکار نقش دارد و اکثر ما از عملکرد آن بیاطلاع هستیم مگر اینکه آسیب دیده باشد، اما نقش مهمی در درک ما از جهتیابی فضایی و حرکت خود به خودی دارد. در واقع، سیستم دهلیزی زیربنای توانایی ما در تشخیص جهت و بزرگی حرکت خود به خودی و تمایز این حرکت از حرکت اشیاء اطراف ما است. سیستم دهلیزی با همکاری سیستم بینایی، اطلاعات حیاتی را برای جهتیابی فضایی و ناوبری فراهم میکند.
As might be expected based on the stimuli that act on the vestibular end organs, the vestibular system contributes to our perception of head rotations, translational head movements, and head tilts. As with other percepts, these vestibular percepts do not simply mirror the physical attributes of the associated stimulus. For example, a blindfolded subject riding on a chair rotating at a constant speed will perceive that the rotation is slowing and, after about 30 seconds, that it has stopped entirely. Interestingly, the time course of this perceptual decay is similar to, but more prolonged than, the decrement of the signal transmitted from the semicircular canal to the brain, suggesting that the brain is somehow compensating for the diminishing signal to generate a percept that more closely approximates the actual rotation. In a similar manner, the vestibular system also contributes to our perception of the direction and velocity of translational movements, as well as the direction and magnitude of static tilts.
همانطور که بر اساس محرکهایی که بر اندامهای انتهایی دهلیزی عمل میکنند، انتظار میرود، سیستم دهلیزی در درک ما از چرخشهای سر، حرکات انتقالی سر و کج شدن سر نقش دارد. همانند سایر ادراکات، این ادراکات دهلیزی صرفاً منعکسکننده ویژگیهای فیزیکی محرک مرتبط نیستند. به عنوان مثال، یک فرد چشمبند زده که بر روی صندلی در حال چرخش با سرعت ثابت سوار است، درک میکند که چرخش در حال کند شدن است و پس از حدود 30 ثانیه، کاملاً متوقف شده است. جالب توجه است که سیر زمانی این زوال ادراکی مشابه، اما طولانیتر از، کاهش سیگنال منتقل شده از کانال نیمدایرهای به مغز است، که نشان میدهد مغز به نوعی در حال جبران سیگنال کاهشیافته است تا ادراکی ایجاد کند که به چرخش واقعی نزدیکتر باشد. به طور مشابه، سیستم دهلیزی همچنین در درک ما از جهت و سرعت حرکات انتقالی و همچنین جهت و بزرگی کج شدنهای استاتیک نقش دارد.
As mentioned, the vestibular and visual systems normally work in concert to provide an estimate of self-motion. As we walk along a forest trail, for example, translational movements activate the vestibular system and also generate visual flow across the retina. The contributions of visual flow to self-motion can be very compelling, as witnessed by riders waiting for their train to leave the station. If a train on an adjacent track starts to move, the resultant visual flow generates a strong sense of self-motion, a perceptual process known as vection. More systematic tests of vestibular-visual interactions using controlled visual stimulation suggest that the normal function of the vestibular system is to suppress the sense of self-generated motion that can arise from visual cues. Consistent with this idea, people with vestibular damage as well as astronauts working in space, who lack gravitational cues from the otolith organs-report heightened vection in response to visual flow information.
همانطور که گفته شد، سیستمهای دهلیزی و بینایی معمولاً با هماهنگی یکدیگر کار میکنند تا تخمینی از حرکت خود ارائه دهند. به عنوان مثال، هنگامی که در امتداد یک مسیر جنگلی قدم میزنیم، حرکات انتقالی سیستم دهلیزی را فعال میکنند و همچنین جریان بصری را در سراسر شبکیه ایجاد میکنند. سهم جریان بصری در حرکت خود میتواند بسیار جذاب باشد، همانطور که توسط مسافرانی که منتظر ترک قطار خود از ایستگاه هستند، مشاهده میشود. اگر قطاری در مسیر مجاور شروع به حرکت کند، جریان بصری حاصل، حس قوی از حرکت خود ایجاد میکند، یک فرآیند ادراکی که به عنوان انتقال شناخته میشود. آزمایشهای سیستماتیکتر از تعاملات دهلیزی-بینایی با استفاده از تحریک بصری کنترلشده نشان میدهد که عملکرد طبیعی سیستم دهلیزی، سرکوب حس حرکت خود تولید شده است که میتواند از نشانههای بصری ناشی شود. مطابق با این ایده، افراد مبتلا به آسیب دهلیزی و همچنین فضانوردانی که در فضا کار میکنند و فاقد نشانههای گرانشی از اندامهای اتولیت هستند، انتقال بیشتری را در پاسخ به اطلاعات جریان بصری گزارش میدهند.
Summary
The vestibular system provides information about the orientation of the head with respect to gravity and head mo- tion. The sensory receptor cells are located in the otolith organs and the semicircular canals of the inner ear. The otolith organs provide information for ocular reflexes and postural adjustments when the head tilts in various directions or undergoes translational movements, and for our perception of these tilts and translations. The semicircular canals, in contrast, provide information about head rotations; these stimuli initiate reflex movements that adjust the eyes, head, and body during motor activities. Among the best studied of these reflexes are eye movements that compensate for head movements, thereby stabilizing the visual scene when the head moves. Information from the vestibular system also plays a central role in our perception of spatial orientation and our ability to navigate through the environment. Vestibular processing is inherently multisensory: Input from all the vestibular organs is integrated with input from the visual and somatosensory systems to provide perceptions of body position and orientation in space.
خلاصه
سیستم دهلیزی اطلاعاتی در مورد جهت گیری سر نسبت به جاذبه و حرکت سر ارائه میدهد. سلولهای گیرنده حسی در اندامهای اتولیت و مجاری نیم دایرهای گوش داخلی قرار دارند. اندامهای اتولیت اطلاعاتی را برای رفلکسهای چشمی و تنظیمات وضعیتی هنگام کج شدن سر در جهات مختلف یا انجام حرکات انتقالی، و برای درک ما از این کج شدنها و حرکات انتقالی ارائه میدهند. در مقابل، مجاری نیم دایرهای اطلاعاتی در مورد چرخشهای سر ارائه میدهند. این محرکها حرکات رفلکسی را آغاز میکنند که چشمها، سر و بدن را در طول فعالیتهای حرکتی تنظیم میکنند. از جمله بهترین رفلکسهای مورد مطالعه، حرکات چشم هستند که حرکات سر را جبران میکنند و در نتیجه صحنه بصری را هنگام حرکت سر تثبیت میکنند. اطلاعات سیستم دهلیزی همچنین نقش محوری در درک ما از جهت گیری فضایی و توانایی ما برای پیمایش در محیط ایفا میکند. پردازش دهلیزی ذاتاً چند حسی است: ورودی از تمام اندامهای دهلیزی با ورودی از سیستمهای بینایی و حسی-پیکری ادغام میشود تا درک موقعیت و جهت گیری بدن در فضا را فراهم کند.
ADDITIONAL READING
کلیک کنید «Reviews»
Angelaki, D. E. and K. E. Cullen (2008) Vestibular system: The many facets of a multimodal sense. Annu. Rev. Neurosci. 31: 125-150.
Benson, A. (1982) The vestibular sensory system. In The Senses, H. B. Barlow and J. D. Mollon (eds.). New York: Cambridge University Press, pp. 333-368.
Brandt, T. (1991) Man in motion: Historical and clinical aspects of vestibular function. A review. Brain 114: 2159-2174.
Cullen, K. E. (2011) The neural encoding of self-motion. Curr. Opin. Neurobiol. 21: 587–595.
Cullen, K. E. (2012) The vestibular system: multimodal integration and encoding of self-motion for motor control. Trends Neurosci. 35: 185-196.
Cullen, K. E. and J. X. Brooks (2015) Neural correlates of sensory prediction errors in monkeys: evidence for internal mod- els of voluntary self motion in the cerebellum. Cerebellum 14:
31-34.
Eatock, R. A. and J. E. Songer (2011) Vestibular hair cells and afferents: two channels for head motion signals. Annu. Rev. Neurosci. 34: 501-534.
Furman, J. M. and R. W. Baloh (1992) Otolith ocular testing in human subjects. Ann. N. Y. Acad. Sci. 656: 431-451.
Goldberg, J. M. (2000) Afferent diversity and the organization of the central vestibular pathways. Exp. Brain Res. 130: 277-297.
Goldberg, J. M. and C. Fernandez (1984) The vestibular system. In Handbook of Physiology. Section 1: The Nervous System, Volume III: Sensory Processes, Part II, J. M. Brookhart, V. B. Mountcastle,I. Darian-Smith and S. R. Geiger (eds.). Bethesda, MD: Ameri- can Physiological Society, pp. 977-1022.
Green, A. M. and D. E. Angelaki (2010) Multisensory integration: resolving sensory ambiguities to build novel representa- tions. Curr. Opin. Neurobiol. 20: 353-360.
Hess, B. J. (2001) Vestibular signals in self orientation and eye movement control. News Physiolog. Sci. 16: 234-238.
Raphan, T. and B. Cohen (2002) The vestibulo-ocular reflex in three dimensions. Exp. Brain Res. 145: 1-27.
کلیک کنید «Important original papers»
Angelaki, D. E., A. G. Shaikh, A. M. Green and J. D. Dickman (2004) Neurons compute internal models of the physical laws of motion. Nature 430: 560-564.
Brooks, J. X., J. Carriot and K. E. Cullen (2015) Learning to expect the unexpected: rapid updating in primate cerebellum during voluntary self-motion. Nat. Neurosci. 18: 1310-1317.
Goldberg, J. M. and C. Fernandez (1971) Physiology of peripheral neurons innervating semicircular canals of the squirrel monkey, Parts 1, 2, 3. J. Neurophysiol. 34: 635-684.
Goldberg, J. M. and C. Fernandez (1976) Physiology of peripheral neurons innervating otolith organs of the squirrel monkey, Parts 1, 2, 3. J. Neurophysiol. 39: 970-1008.
Laurens, J., H. Meng and D. E. Angelaki (2013) Neural representation of orientation relative to gravity in the macaque cerebellum. Neuron 80: 1508-1518.
Lindeman, H. H. (1973) Anatomy of the otolith organs. Adv. Otorhinolaryngol. 20: 405-433.
Merfeld, D. M. (1995) Modeling the vestibular-ocular reflex of the squirrel monkey during eccentric rotation and roll tilt. Exp. Brain. Res. 106: 123-134.
کلیک کنید «Books»
Baloh, R. W. (1998) Dizziness, Hearing Loss, and Tinnitus. Phila- delphia: F. A. Davis Company.
Baloh, R. W. and V. Honrubia (2001) Clinical Neurophysiology of the Vestibular System, 3rd Edition. New York: Oxford University Press.
Go to the NEUROSCIENCE 6e Companion Website at oup-arc.com/access/purves-6e for Web Topics,- Animations, Flashcards, and more. Go to DASHBOARD for additional resources and assessments.
🚀 با ما همراه شوید!
تازهترین مطالب و آموزشهای مغز و اعصاب را از دست ندهید. با فالو کردن کانال تلگرام، از ما حمایت کنید!