نقش حواس چشایی و بویایی در درک طعم، انتخاب غذا و رمزگشایی محرکهای شیمیایی در سیستم عصبی

دعای مطالعه [ نمایش ]
بِسْمِ الله الرَّحْمنِ الرَّحیمِ
اَللّهُمَّ اَخْرِجْنى مِنْ ظُلُماتِ الْوَهْمِ
خدايا مرا بيرون آور از تاريكىهاى وهم،
وَ اَكْرِمْنى بِنُورِ الْفَهْمِ
و به نور فهم گرامى ام بدار،
اَللّهُمَّ افْتَحْ عَلَيْنا اَبْوابَ رَحْمَتِكَ
خدايا درهاى رحمتت را به روى ما بگشا،
وَانْشُرْ عَلَيْنا خَزائِنَ عُلُومِكَ بِرَحْمَتِكَ يا اَرْحَمَ الرّاحِمينَ
و خزانههاى علومت را بر ما باز كن به امید رحمتت اى مهربانترين مهربانان.
حواس چشایی (taste) و بویایی (smell) به ما این امکان را میدهند که غذاهای نامطلوب یا حتی کشنده را از غذاهایی که خوردن آنها خوشایند و مغذی هستند جدا کنیم. آنها همچنین باعث برانگیختن پاسخهای فیزیولوژیکی میشوند که در هضم و استفاده از غذاها نقش دارند. حس بویایی همچنین به جانوران اجازه میدهد تا نزدیکی سایر جانوران یا حتی افراد را در بین جانوران تشخیص دهند. در نهایت، هر دو حس به شدت با عملکردهای عاطفی و رفتارهای ابتدایی سیستم عصبی ما مرتبط هستند. در این فصل، چگونگی شناسایی محرکهای چشایی و بویایی و نحوه کدگذاری آنها در سیگنالهای عصبی ارسال شده به مغز را مورد بحث قرار میدهیم.
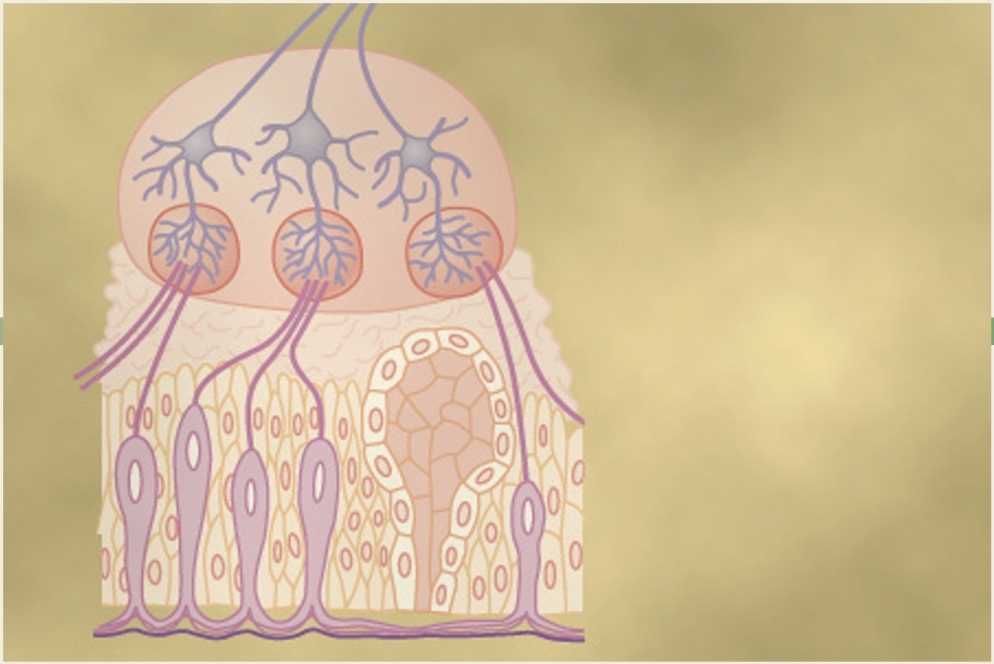
حس چشایی
چشایی عمدتاً عملکرد جوانههای چشایی (taste buds) در دهان است، اما تجربه رایجی است که حس بویایی فرد نیز به شدت به درک چشایی کمک میکند. علاوه بر این، بافت غذا (texture of food)، همانطور که توسط حواس لامسه (tactual senses) دهان تشخیص داده میشود، و وجود موادی مانند فلفل (pepper) در غذا که باعث تحریک انتهای درد (pain endings) میشوند، تجربه چشایی را تا حد زیادی تغییر میدهد. اهمیت چشایی از این واقعیت سرچشمه میگیرد که به فرد اجازه میدهد غذا را مطابق با خواستهها و اغلب مطابق با نیاز متابولیکی بافتهای بدن به مواد خاص انتخاب کند.
حواس اولیه چشایی
ماهیت مواد شیمیایی خاصی که گیرندههای مختلف چشایی را تحریک میکنند، به طور کامل مشخص نیست. با این حال، مطالعات سایکوفیزیولوژیک (psychophysiologic) و نوروفیزیولوژیک (neurophysiologic) حداقل 13 گیرنده شیمیایی احتمالی را در سلولهای چشایی شناسایی کرده اند، این گیرندهها به شرح زیر میباشند:
- 2 گیرنده سدیم (sodium receptors)،
- 2 گیرنده پتاسیم (potassium receptors)،
- 1 گیرنده کلرید (chloride receptor)،
- 1 گیرنده آدنوزین (adenosine receptor)،
- 1 گیرنده اینوزین (inosine receptor)،
- 2 گیرنده شیرینی (sweet receptors)،
- 2 گیرنده تلخی (bitter receptors)،
- 1 گیرنده گلوتامات (glutamate receptor) و
- 1 گیرنده یون هیدروژن (hydrogen ion receptor).
برای تحلیل کاربردی چشایی، قابلیتهای گیرندههای فوقالذکر نیز در پنج دسته کلی طبقهبندی شدهاند و حواس اولیه چشایی نامگذاری شدهاند. آنها عبارتند از مزههای ترشی (sour)، شوری (salty)، شیرینی (sweet)، تلخی (bitter) و «اومامی» (umami).
یک فرد میتواند صدها طعم مختلف را درک کند. همه آنها قرار است ترکیبی از حواس اولیه چشایی (elementary taste sensations) باشند، همانطور که تمام رنگهایی که میبینیم ترکیبی از سه رنگ اصلی هستند که در فصل 50 توضیح داده شد.
مزه ترشی
مزه ترشی توسط اسیدها ایجاد میشود، یعنی غلظت یون هیدروژن، و شدت احساس این مزه تقریباً متناسب با لگاریتم غلظت یون هیدروژن (logarithm of the hydrogen ion concentration) است. یعنی هر چه غذا اسیدیتر باشد، حس ترشی آن بیشتر است.
مزه شوری
مزه شوری توسط نمکهای یونیزه شده، عمدتاً توسط غلظت یون سدیم (sodium ion concentration)، ایجاد میشود. کیفیت مزه از نمکی به نمک دیگر تا حدودی متفاوت است، زیرا برخی از نمکها علاوه بر شوری، حس چشایی دیگری را نیز برمیانگیزند. کاتیون نمکها، به ویژه کاتیونهای سدیم، عمدتاً مسئول مزه شوری هستند، اما آنیونها (anions) نیز به میزان کمتری نقش دارند.
مزه شیرینی
مزه شیرینی توسط یک دسته ویژه از مواد شیمیایی ایجاد نمیشود. برخی از انواع مواد شیمیایی که باعث ایجاد این مزه میشوند عبارتند از قندها (sugars)، گلیکولها (glycols)، الکلها (alcohols)، آلدئیدها (aldehydes)، کتونها (ketones)، آمیدها (amides)، استرها (esters)، برخی اسیدهای آمینه (amino acids)، برخی پروتئینهای کوچک (small proteins)، اسیدهای سولفونیک (sulfonic acids)، اسیدهایهالوژنه (halogenated acids) و نمکهای معدنی سرب و بریلیم (inorganic salts of lead and beryllium). به طور خاص توجه داشته باشید که بیشتر موادی که باعث مزه شیرینی میشوند، مواد شیمیایی آلی هستند. به خصوص جالب است که تغییرات جزئی در ساختار شیمیایی، مانند افزودن یک رادیکال ساده، اغلب میتواند ماده را از شیرینی به تلخی تبدیل کند.
مزه تلخی
مزه تلخی، مانند مزه شیرینی، توسط تنها یک نوع عامل شیمیایی ایجاد نمیشود. در اینجا نیز موادی که مزه تلخی میدهند تقریباً کاملاً مواد آلی هستند. دو دسته خاص از مواد بهویژه باعث ایجاد مزه تلخی میشوند: (1) مواد آلی با زنجیره بلند که حاوی نیتروژن هستند و (2) آلکالوئیدها. آلکالوئیدها (alkaloids) شامل بسیاری از داروهای مورد استفاده در پزشکی مانند کینین (quinine)، کافئین (caffeine)، استریکنین (strychnine) و نیکوتین (nicotine) هستند.
برخی مواد در ابتدا مزه شیرینی دارند، سپس مزه تلخی ایجاد میکنند. این موضوع در مورد ساخارین (saccharin) صدق میکند که این ماده را برای برخی افراد ناخوشایند میکند.
مزه تلخی زمانی که شدت بالایی داشته باشد، معمولاً باعث میشود انسان یا حیوان از غذا اجتناب کنند. این مورد، بدون شک یکی از عملکردهای مهم حس چشایی تلخی است زیرا بسیاری از سموم کشنده (deadly toxins) موجود در گیاهان آلکالوئیدی هستند و تقریباً همه این آلکالوئیدها باعث ایجاد حس تلخی شدیدی میشوند و معمولاً باعث اجتناب از غذا میشود.
مزه اومامی
Umami یک کلمه ژاپنی (به معنی «خوشمزه» delicious) است که نشان دهنده یک مزهٔ دلپذیر و خوشایند است که از نظر کیفی با ترشی، شوری، شیرینی و یا تلخی متفاوت است. اومامیطعم غالب غذاهای حاوی ال-گلوتامات (l-glutamate) مانند عصاره گوشت (meat extracts) و پنیر مانده (aging cheese) است و برخی فیزیولوژیستها آن را جزء دسته پنجم محرکهای چشایی اولیه میدانند.
یک گیرنده چشایی برای L-گلوتامات ممکن است به یکی از گیرندههای گلوتامات مرتبط باشد که در سیناپسهای عصبی مغز نیز بیان میشود. با این حال، مکانیسمهای مولکولی دقیق مسئول مزه umami هنوز نامشخص است.
آستانه چشایی
آستانه تحریک (threshold for stimulation) مزه ترشی توسط اسید هیدروکلریک (hydrochloric acid) یا کلریدریک اسید به طور متوسط 0.0009 مولار [در منبع به اشتباه نوشته شده N] است. برای تحریک مزه شوری توسط کلرید سدیم، 0.01 مولار، برای مزه شیرینی توسط ساکارز، 0.01 مولار و برای مزه تلخی کینین، 0.000008 مولار. توجه داشته باشید که حساسیت چشایی به تلخی نسبت به سایر حسها بسیار زیاد است، که مورد انتظار هم میباشد، زیرا این حس عملکرد محافظتی مهمیدر برابر بسیاری از سموم خطرناک در غذا ایجاد میکند.
جدول 53-1 شاخصهای نسبی چشایی (معکوس آستانه چشایی) مواد مختلف را نشان میدهد. در این جدول، شدت چهار حس اولیه چشایی به ترتیب نسبت به شدت چشایی اسید کلریدریک، کینین، ساکارز و کلرید سدیم ارائه شده است و شدت چشایی هر یک از این چهار ماده به صورت قراردادی با شاخص چشایی (taste index) 1 در نظر گرفته شدهاند.
جدول 53-1 شاخصهای نسبی چشایی مواد مختلف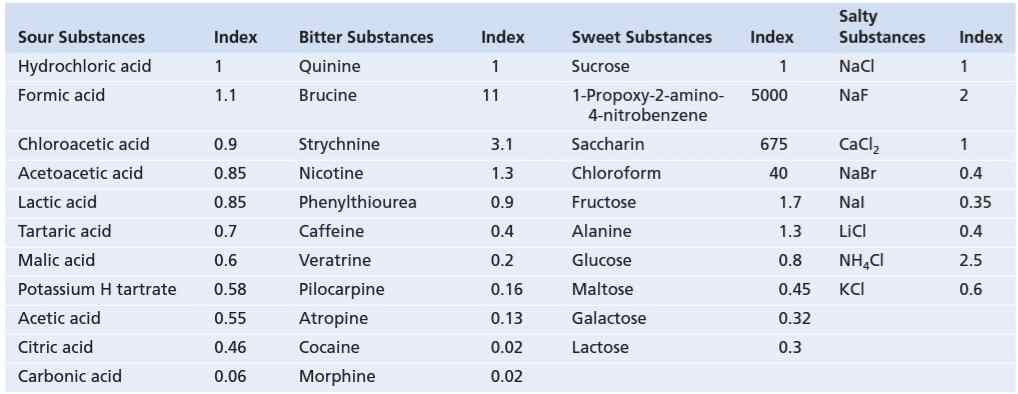
کوری چشایی
برخی از افراد نسبت به برخی از مواد، به ویژه برای انواع مختلف ترکیبات تیواوره (thiourea)، کوری چشایی (taste blindness) هستند. ماده ای که اغلب توسط روانشناسان برای نشان دادن کوری چشایی استفاده میشود فنیل تیوکاربامید (phenylthiocarbamide) است که حدود 15 تا 30 درصد از مردم نسبت به آن کوری چشایی نشان میدهند. درصد دقیق به روش آزمایش و غلظت ماده بستگی دارد.
جوانه چشایی و عملکرد آن
شکل 53-1 یک جوانه چشایی را نشان میدهد که قطر آن حدود ![]() میلیمتر و طول آن حدود
میلیمتر و طول آن حدود ![]() میلیمتر است. جوانه چشایی از حدود 50 سلول اپیتلیال اصلاح شده (modified epithelial cells) تشکیل شده است که برخی از آنها سلولهای پشتیبان به نام سلولهای sustentacular و برخی دیگر سلولهای چشایی هستند. سلولهای چشایی به طور مداوم با تقسیم میتوزی سلولهای اپیتلیال اطراف جایگزین میشوند، بنابراین برخی از سلولهای چشایی سلولهای جوان هستند. برخی دیگر سلولهای بالغی هستند که به سمت مرکز جوانه قرار دارند. اینها به زودی تخریب میشوند و از بین میروند. طول عمر هر سلول چشایی در پستانداران پستتر حدود 10 روز است اما برای انسان مشخص نیست.
میلیمتر است. جوانه چشایی از حدود 50 سلول اپیتلیال اصلاح شده (modified epithelial cells) تشکیل شده است که برخی از آنها سلولهای پشتیبان به نام سلولهای sustentacular و برخی دیگر سلولهای چشایی هستند. سلولهای چشایی به طور مداوم با تقسیم میتوزی سلولهای اپیتلیال اطراف جایگزین میشوند، بنابراین برخی از سلولهای چشایی سلولهای جوان هستند. برخی دیگر سلولهای بالغی هستند که به سمت مرکز جوانه قرار دارند. اینها به زودی تخریب میشوند و از بین میروند. طول عمر هر سلول چشایی در پستانداران پستتر حدود 10 روز است اما برای انسان مشخص نیست.
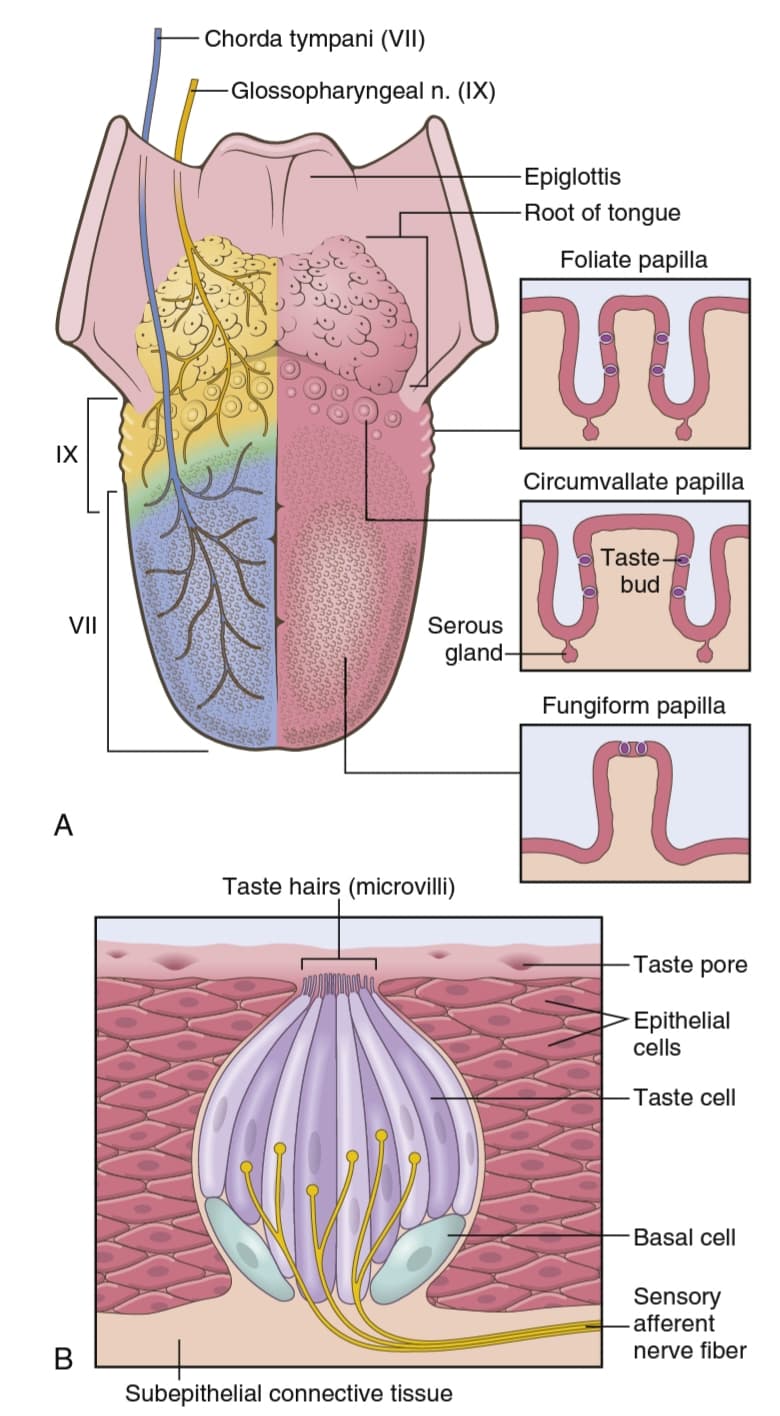 شکل 1-53 جوانه چشایی.
شکل 1-53 جوانه چشایی.
نوک خارجی سلولهای چشایی در اطراف یک منفذ کوچک چشایی (minute taste pore) قرار گرفته اند که در شکل 1-53 نشان داده شده است. از نوک هر سلول چشایی، چندین میکروویلی (microvilli) یا موی چشایی (taste hairs) به سمت خارج به درون منفذ چشایی بیرون زده تا به حفره دهان میرسند. این میکروویلیها سطح گیرنده را برای چشایی فراهم میکنند.
یک شبکه از انتهای منشعب شدهٔ رشتههای عصبی چشایی در اطراف تنه سلولهای چشایی وجود دارد که توسط سلولهای گیرنده چشایی تحریک میشوند. برخی از این رشتههای عصبی در چینهای غشای سلول چشایی فرو میروند. تعداد زیادی وزیکول (vesicles) در زیر غشای سلول در نزدیکی رشتههای عصبی وجود دارد. اعتقاد بر این است که این وزیکولها حاوی ماده انتقال دهنده عصبی (neurotransmitter substance) هستند که از غشای سلول آزاد میشوند تا پایانههای رشتهٔ عصبی را در پاسخ به تحریک چشایی، تحریک کند.
موقعیت جوانههای چشایی
جوانههای چشایی روی سه نوع پاپیلای زبان یافت میشوند که شرح آنها به صورت زیر است: (1) تعداد زیادی جوانه چشایی روی دیوارههای فرورفتهای وجود دارند که پاپیلاهای گنبدیشکل (circumvallate papillae) را احاطه کردهاند. این برجستگیها یک خط V شکل را در سطح خلفی زبان ایجاد میکنند. (2) تعداد متوسطی از جوانههای چشایی روی پاپیلاهای قارچی شکل (fungiform papillae) در سطح صاف قدامیزبان قرار دارند. (3) تعداد متوسطی از جوانههای چشایی روی پاپیلاهای برگیشکل (foliate papillae) در چینهایی قرار دارند که در سطوح جانبی زبان واقع شدهاند. تعدادی جوانه چشایی نیز روی کام (palate) قرار دارند و تعداد کمیروی ستونهای لوزهها (tonsillar pillars)، روی اپی گلوت (epiglottis) و حتی در پروگزیمال (ابتدای) مری (proximal esophagus) یافت میشوند. تعداد جوانههای چشایی در بزرگسالان 3000 تا 10000 و در کودکان کمیبیشتر است. بعد از سن 45 سالگی، بسیاری از جوانههای چشایی تحلیل میروند و باعث کاهش حس چشایی در سنین بالا میشود.
اختصاصی بودن جوانههای چشایی برای یک محرک چشایی اولیه
مطالعات میکروالکترودی از جوانههای چشایی منفرد نشان میدهد که زمانی که غلظت ماده تولید کننده حس چشایی کم است هر جوانه چشایی معمولاً به یکی از پنج محرک اولیه چشایی پاسخ میدهد. اما اگر غلظت ماده زیاد باشد، بیشتر جوانهها میتوانند توسط دو یا چند محرک چشایی اولیه و همچنین توسط چند محرک چشایی دیگر که در تقسیمبندی “محرکهای اولیه” قرار نمیگیرند، برانگیخته میشوند.
مکانیسم تحریک جوانههای چشایی
پتانسیل گیرنده
غشای سلول چشایی، مانند بسیاری از سلولهای گیرنده حسی دیگر، در داخل نسبت به خارج دارای بار منفی است. اعمال یک ماده چشایی روی موهای چشایی باعث کاهش نسبی این پتانسیل منفی میشود، یعنی سلول چشایی دپلاریزه میشود. در بیشتر موارد، کاهش پتانسیل، در محدوده وسیع، تقریباً با لگاریتم غلظت ماده محرک متناسب است. این تغییر در پتانسیل الکتریکی سلول چشایی را پتانسیل گیرنده چشایی (receptor potential for taste) مینامند.
مکانیسمیکه توسط آن اکثر مواد محرک با پرزهای چشایی واکنش میدهند تا پتانسیل گیرنده را ایجاد کنند، با اتصال ماده شیمیایی چشایی به یک مولکول گیرنده پروتئینی است که روی سطح خارجی سلول گیرنده چشایی در نزدیکی غشای پرزها و یا برآمده از غشای پرزها قرار دارد. این به نوبه خود کانالهای یونی را باز میکند که به یونهای سدیم یا یونهای هیدروژن با بار مثبت اجازه میدهد وارد شوند و سلول را دپولاریزه کنند. سپس خود ماده شیمیایی چشایی به تدریج توسط بزاق (saliva) از پرز چشایی (taste villus) شسته میشود تا محرک حذف شود.
نوع پروتئین گیرنده در هر پرز چشایی تعیین کننده نوع مزهای است که درک میشود. برای یونهای سدیم و یونهای هیدروژن که به ترتیب مزههای شوری و ترشی را ایجاد میکنند، پروتئینهای گیرنده کانالهای یونی خاصی را در غشای آپیکال (رأسی) سلولهای چشایی باز میکنند و در نتیجه گیرندهها را فعال میکنند. با این حال، در مورد مزههای شیرینی و تلخی، بخشهایی از مولکولهای پروتئین گیرنده که از غشاهای راسی (apical membranes) بیرون زدهاند، پیامبرهای ثانویه (second-messenger) داخل سلولی را فعال میکنند و این پیامبرهای ثانویه باعث ایجاد تغییرات شیمیایی درون سلول میشوند که سیگنالهای چشایی را برمیانگیزد.
تولید تکانههای عصبی توسط جوانه چشایی
در ابتدای اعمال محرک چشایی، سرعت تخلیه رشتههای عصبی از جوانههای چشایی در کسری از ثانیه به اوج خود میرسد، اما پس از آن، در چند ثانیه بعدی سازش یافته و به سطحی پایین و پایدار میرسد که تا زمانی که محرک تداوم دارد در همین حد میماند. بنابراین، یک سیگنال فوری قوی توسط عصب چشایی ارسال میشود و یک سیگنال پیوسته ضعیفتر تا زمانی که جوانه چشایی در معرض محرک چشایی باشد، منتقل میشود.
انتقال سیگنالهای چشایی به سیستم عصبی مرکزی
شکل 2-53، مسیرهای عصبی انتقال سیگنالهای چشایی از زبان و ناحیه حلق به سیستم عصبی مرکزی را نشان میدهد. تکانههای چشایی (Taste impulses) دو سوم قدامیزبان ابتدا به عصب زبانی (lingual nerve) وارد شده، سپس از طریق طناب صماخی (chorda tympani) به عصب صورت (facial nerve) و در نهایت به دسته منزوی (tractus solitarius) در ساقه مغز میرسند. حس چشایی از پاپیلاهای گنبدی شکل (circumvallate papillae) در پشت زبان و سایر نواحی خلفی دهان و گلو نیز توسط عصب گلوسوفارنژیال (glossopharyngeal nerve) به tractus solitarius منتقل میشود، اما کمیدر سطح خلفیتر قرار میگیرند. در نهایت، چند سیگنال چشایی به داخل مخابره میشود tractus solitarius از قاعده زبان و سایر قسمتهای ناحیه حلق از طریق عصب واگ.
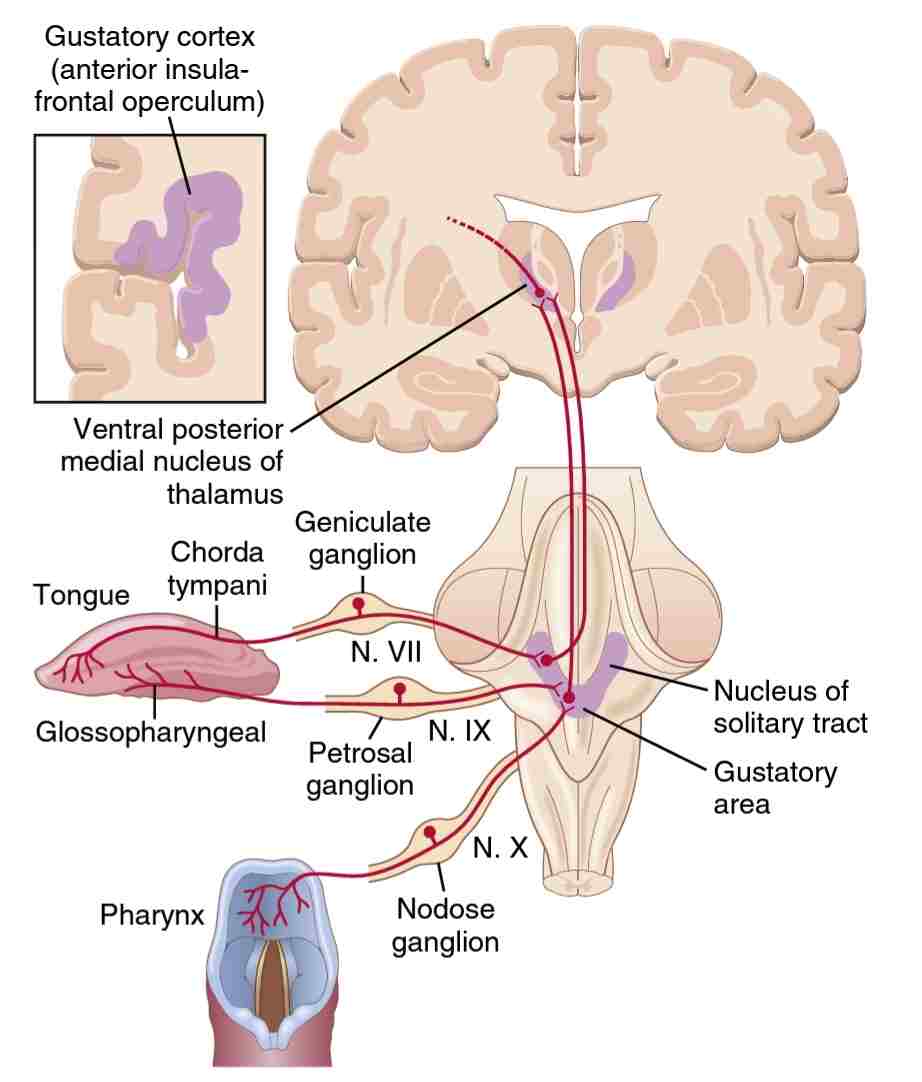 شکل 2-53 انتقال سیگنالهای چشایی به سیستم عصبی مرکزی.
شکل 2-53 انتقال سیگنالهای چشایی به سیستم عصبی مرکزی.
تمام رشتههای چشایی در هستههای دسته منزوی (tractus solitarius) در ناحیه خلفی ساقه مغز (posterior brain stem) سیناپس میدهند. این هستهها، نورونهای مرتبه دوم را به ناحیه کوچکی از هسته داخلی خلفی شکمی(ventral posterior medial nucleus) تالاموس میفرستند، که نسبت به محل ختم نواحی صورتی سیستم ستون خلفی-لمنیسکوس میانی کمیدر قسمت داخلیتر تالاموس قرار دارند. سپس از تالاموس، نورونهای درجه سوم به قسمت تحتانی شکنج پس مرکزی (lower tip of the postcentral gyrus) در قشر مغزی آهیانه (parietal cerebral cortex,) در محلی که به صورت عمیق وارد شیار سیلویوس (sylvian fissure) شده و ، ناحیه مجاور اپرکولوم اینسولار (adjacent opercular insular area) میروند. این قسمت کمیدر سمت جانبی، شکمیو روسترال (lateral, ventral, and rostral) نسبت به ناحیه سیگنالهای لمسی زبان (tongue tactile) در قشر پیکری I (cerebral somatic area I) قرار دارد. با این توصیف از مسیرهای چشایی، مشهود است که فییرهای نورونهای مسیر چشایی نزدیک و به موازات مسیرهای حسی-پیکری زبان (somatosensory pathways from the tongue) قرار دارند.
رفلکسهای چشایی در ساقه مغز ادغام شده اند
از مسیر منزوی (tractus solitarius)، درون ساقه مغز سیگنالهای چشایی زیادی مستقیماً به هستههای بزاقی فوقانی و تحتانی (superior and inferior salivatory nuclei) ارسال میشوند و این نواحی سیگنالهایی را به غدد زیر فکی (submandibular)، زیر زبانی (sublingual) و پاروتید (parotid glands) میفرستند تا طی بلع و هضم غذا به کنترل ترشح بزاق کمک کنند.
تطابق سریع چشایی
همه با این واقعیت آشنا هستند که حس چشایی به سرعت تطابق مییابد، اغلب در حضور مداوم محرک (continuous stimulation) تقریباً در عرض یک دقیقه یا بیشتر به طور کامل سازگار میشود. با این حال، از مطالعات الکتروفیزیولوژیک فیبرهای عصبی چشایی، مشخص است که معمولاً بیش از نیمیاز این سازشها مربوط به تطابق خود جوانههای چشایی نیست. بنابراین، حداکثر میزان سازگاری که در حس چشایی رخ میدهد، تقریباً به طور قطع در خود سیستم عصبی مرکزی ایجاد میشود، اگرچه مکانیسم و مکان آن هنوز مشخص نیست. به هر حال، این مکانیسم با بسیاری از سیستمهای حسی دیگر که سازش در آنها تقریباً به طور کامل در گیرندهها انجام میشود، متفاوت است.
ترجیح مزه و کنترل رژیم غذایی
ترجیح چشایی به صورت ساده به این معناست که یک حیوان انواع خاصی از غذاها را به سایر غذاها ترجیح میدهد، و حیوان به طور خودکار از آن برای کمک به کنترل رژیم غذایی خود استفاده میکند. علاوه بر این، این ترجیحات چشایی اغلب براساس نیاز بدن به برخی مواد خاص تغییر میکند.
مطالعات زیر این توانایی جانوران را در انتخاب غذا مطابق با نیازهای بدنشان نشان میدهد. ابتدا جانورانی که آدرنالکتومی (غده فوق کلیه آنها برداشته) شده و بنابراین نمک بدنشان به شدت کاهش یافته و نیازمند نمک میباشند، به طور خودکار آب آشامیدنی با غلظت بالای کلرید سدیم را به آب خالص ترجیح میدهند و اغلب این انتخاب برای تامین نیازهای بدن کافی بوده و از مرگ ناشی از کاهش نمک جلوگیری میکند. دوم، حیوانی که در اثر تزریق مقادیر بیش از حد انسولین (insulin)، دچار کاهش شدید قند خون (blood sugar) شده، بهطور خودکار شیرینترین غذا را از میان نمونههای زیاد غذاها انتخاب میکند. سوم، جانوران پاراتیروئیدکتومیشده (غده پاراتیروئید آنها برداشته شده: parathyroidectomized) و بنابراین دچار کمبود کمبود کلسیم شدهاند به طور خودکار آشامیدنی با غلظت بالای کلرید کلسیم را انتخاب میکنند.
همین پدیدهها در زندگی روزمره نیز مشاهده میشود. به عنوان مثال، «رسوبات نمکی» (salt licks) مناطق بیابانی سبب جذب جانوران از دور و نزدیک میشود. همچنین انسان هر غذایی که دارای طعم ناخوشایند (unpleasant affective sensation) باشد را رد میکند که در بسیاری از موارد از بدن ما در برابر مواد نامطلوب محافظت میکند.
پدیده ترجیح چشایی (phenomenon of taste preference) تقریباً به طور قطع ناشی از مکانیزمیاست که در سیستم عصبی مرکزی قرار دارد و مربوط به گیرندههای چشایی (taste receptors) نیست، اگرچه گیرندهها اغلب نسبت به یک ماده مغذی مورد نیاز حساس میشوند. یک دلیل مهم برای این باور که ترجیح چشایی عمدتاً یک پدیده سیستم عصبی مرکزی است، این است که تجربه قبلی با طعمهای ناخوشایند یا خوشایند (unpleasant or pleasant tastes) نقش عمده ای در تعیین ترجیحات چشایی فرد دارد. به عنوان مثال، اگر فردی بلافاصله پس از خوردن نوع خاصی از غذا مریض شود، معمولاً پس از آن نسبت به آن غذای خاص ترجیح چشایی منفی (negative taste preference) یا به اصطلاح تنفر چشایی (taste aversion) پیدا میکند. همین اثر را میتوان در جانوران پستتر (lower animals) نشان داد.
کلیک کنید «Bibliography: فهرست کتب مربوطه»
Augustine V, Gokce SK, Oka Y: Peripheral and central nutrient sensing underlying appetite regulation. Trends Neurosci 41:526, 2018.
Avau B, Depoortere 1: The bitter truth about bitter taste receptors: beyond sensing bitter in the oral cavity. Acta Physiol (Oxf) 216:407, 2016.
Besnard P, Passilly-Degrace P, Khan NA: Taste of fat: a sixth taste modality? Physiol Rev 96:151, 2016.
Buck LB. The molecular architecture of odor and pheromone sensing in mammals. Cell 100:611, 2000.
Chandrashekar J, Hoon MA, Ryba NJ, Zuker CS: The receptors and cells for mammalian taste. Nature 444:288, 2006.
Lodovichi C, Belluscio L: Odorant receptors in the formation of the olfactory bulb circuitry. Physiology (Bethesda) 27:200, 2012.
Mizrahi A: The hard and soft wired nature of the olfactory map. Trends Neurosci 41:872, 2018.
Mori K, Takahashi YK, Igarashi KM, Yamaguchi M: Maps of odorant molecular features in the mammalian olfactory bulb. Physiol Rev 86:409, 2006.
Palmer RK: A Pharmacological perspective on the study of taste. Phar- macol Rev 71:20, 2019.
Roper SD: The taste of table salt. Pflugers Arch. 467:457, 2015. Roper SD, Chaudhari N: Taste buds: cells, signals and synapses. Nat
Rev Neurosci 18:485, 2017.
Schier LA, Spector AC: The functional and neurobiological properties of bad taste. Physiol Rev 99:605, 2019.
Smith DV, Margolskee RF: Making sense of taste. Sci Am 284:32, 2001.
Tizzano M, Finger TE: Chemosensors in the nose: guardians of the airways. Physiology (Bethesda) 28:51, 2013.
Yarmolinsky DA, Zuker CS, Ryba NJ: Common sense about taste: from mammals to insects. Cell 16;139:234, 2009.