اساس سلولی یادگیری و حافظه: نقش LTP، قانون هب و پلاستیسیته سیناپسی در هیپوکامپ

دعای مطالعه [ نمایش ]
بِسْمِ الله الرَّحْمنِ الرَّحیمِ
اَللّهُمَّ اَخْرِجْنى مِنْ ظُلُماتِ الْوَهْمِ
خدايا مرا بيرون آور از تاريكىهاى وهم،
وَ اَكْرِمْنى بِنُورِ الْفَهْمِ
و به نور فهم گرامى ام بدار،
اَللّهُمَّ افْتَحْ عَلَيْنا اَبْوابَ رَحْمَتِكَ
خدايا درهاى رحمتت را به روى ما بگشا،
وَانْشُرْ عَلَيْنا خَزائِنَ عُلُومِكَ بِرَحْمَتِكَ يا اَرْحَمَ الرّاحِمينَ
و خزانههاى علومت را بر ما باز كن به امید رحمتت اى مهربانترين مهربانان.
کتاب «علوم اعصاب شناختی: زیستشناسی ذهن» اثر مایکل گازانیگا و همکاران، همچنان مرجع طلایی این حوزه است که با ترکیب تازهترین پژوهشها و کاربردهای بالینی، پلی میان دانش پایه و عمل پزشکی میسازد.
تلاش علمی و تدریس ارزشمند استاد محترم آقای دکتر محمدعلی نظری الهامبخش نگارش و ترجمه این اثر بوده است.
ترجمه دقیق این اثر توسط برند علمی آیندهنگاران مغز به مدیریت داریوش طاهری، دسترسی فارسیزبانان به مرزهای نوین علوم اعصاب را ممکن ساخته و رسالتی علمی برای شناخت عمیقتر مغز و ساختن آیندهای روشنتر فراهم آورده است.
»» فصل ۹: فصل حافظه؛ قسمت یازدهم
»» CHAPTER 9: Memory
9.7 Cellular Basis of Learning
9.7 اساس سلولی یادگیری
Learning and Memory
یادگیری و حافظه
Researchers have long believed the synapse, with its dynamic connections, to be a likely structure involved in the mechanics of memory. Most models of the cellular bases of memory hold that memory is the result of changes in the strength of synaptic interactions among neurons in neural networks. How would synaptic strength be altered to enable learning and memory?
محققان مدتها بر این باور بودند که احتمالا ساختار سیناپس، با اتصالات دینامیکی آن، در مکانیک حافظه درگیر باشد. اکثر مدلهای اساس سلولی حافظه بر این باورند که حافظه نتیجه تغییر در قدرت تعاملات سیناپسی بین نورونها در شبکههای عصبی است. چگونه میتوان قدرت سیناپسی را برای فعال کردن یادگیری و حافظه تغییر داد؟
In 1949, Donald Hebb, the father of neural network theory, proposed that synaptic connections between coactivated cells change in a manner dependent on their activity. This theory, known as Hebb’s law, is commonly summarized as “Cells that fire together wire together.” Hebb proposed that the mechanism underlying learning was the strengthening of synaptic connections that results when a weak input and a strong input act on a cell at the same time. This learning theory has been dubbed Hebbian learning.
در سال 1949، دونالد هب، پدر نظریه شبکههای عصبی، پیشنهاد کرد که ارتباطات سیناپسی بین سلولهای فعال شده به شیوه ای وابسته به فعالیت آنها تغییر میکند. این نظریه که به قانون هب معروف است، معمولاً به عنوان «سلولهایی که با هم آتش میزنند به هم متصل میشوند» خلاصه میشود. هب پیشنهاد کرد که مکانیسم زیربنایی یادگیری، تقویت اتصالات سیناپسی است که زمانی حاصل میشود که یک ورودی ضعیف و یک ورودی قوی به طور همزمان روی یک سلول عمل کنند. این نظریه یادگیری را یادگیری هب نامیده اند.

FIGURE 9.39 Synaptic organization of the rat hippocampus. The major projection pathways of the rat hippocampus are analogous to those in humans.
شکل 9.39 سازماندهی سیناپسی هیپوکامپ موش. مسیرهای پروجکشن اصلی هیپوکامپ موش مشابه مسیرها در انسان است.
Long-Term Potentiation and the Hippocampus
تقویت طولانی مدت و هیپوکامپ
Because of the role that the hippocampal formation plays in memory, it has long been hypothesized that neurons in the hippocampus must be plastic that is, able to change their synaptic interactions. Although it is now clear that storage itself does not reside in the hippocampus, this fact does not invalidate the hippocampal models that we will examine, because the same cellular mechanisms can operate in various cortical and subcortical areas.
به دلیل نقشی که تشکیلات هیپوکامپ در حافظه ایفا میکند، مدتهاست این فرضیه وجود دارد که نورونهای هیپوکامپ باید انعطافپذیر باشند که قادر به تغییر برهمکنشهای سیناپسی خود باشند. اگرچه اکنون واضح است که خود ذخیره سازی در هیپوکامپ قرار ندارد، اما این واقعیت مدلهای هیپوکامپ را که ما بررسی خواهیم کرد، بی اعتبار نمیکند، زیرا مکانیسمهای سلولی مشابهی میتوانند در نواحی مختلف قشری و زیر قشری عمل کنند.
First let’s establish the three major excitatory neural pathways of the hippocampus that extend from the subiculum to the CA1 cells: the perforant pathway, the mossy fibers, and the Schaffer collaterals (Figure 9.39). Neocortical association areas project via the parahippocampal cortex or perirhinal cortex to the entorhinal cortex. Neurons from the entorhinal cortex travel via the subiculum along the perforant pathway to synapse with granule cells of the dentate gyrus with excitatory inputs. The granule cells have distinctive-looking unmyelinated axons, known as mossy fibers, which connect the dentate gyrus to the dendritic spines of the hippocampal CA3 pyramidal cells. The CA3 pyramidal cells are connected to the CA1 pyramidal cells by axon collaterals, known as the Schaffer collaterals. This hippocampal system is used to examine synaptic plasticity as the mechanism of learning at the cellular level.
ابتدا اجازه دهید سه مسیر عصبی تحریکی اصلی هیپوکامپ را که از سابیکولوم تا سلولهای CA1 گسترش مییابد، معین کنیم: مسیر پرفورنت، فیبرهای خزهای و شاخههای جانبی شافر (شکل 9.39). نواحی ارتباطی نئوکورتیکال از طریق قشر پاراهیپوکامپ یا قشر پریرینال به قشر انتورینال پیش میرود. نورونهای قشر انتورینال از طریق سابیکولوم در امتداد مسیر پرفورنت به سمت سلولهای گرانول شکنج دندانهدار با ورودیهای تحریکی حرکت میکنند تا با آنها سیناپس کنند. سلولهای گرانول دارای آکسونهای غیرمیلین با ظاهری متمایز هستند که به فیبرهای خزهای معروف هستند که شکنج دندانهدار را به خارهای دندریتی سلولهای هرمی CA3 هیپوکامپ متصل میکنند. سلولهای هرمی CA3 توسط شاخههای جانبی آکسونی به سلولهای هرمی CA1 متصل میشوند که به شاخههای جانبی شافر معروف هستند. این سیستم هیپوکامپ برای بررسی انعطافپذیری سیناپسی به عنوان مکانیسم یادگیری در سطح سلولی استفاده میشود.
Physiological studies in rabbits done by Timothy Bliss and Terje Lømo (1973) provided evidence for Hebb’s law. Bliss and Lømo found that stimulating the axons of the perforant pathway resulted in a long-term increase in the magnitude of excitatory postsynaptic potentials (EPSPs). That is, stimulation led to greater synaptic strength in the perforant pathway so that, when the axons were stimulated again later, larger postsynaptic responses resulted in the granule cells of the dentate gyrus. This phenomenon is known as long-term potentiation (LTP) (potentiate means “to strengthen or make more potent”), and its discovery confirmed Hebb’s law.
مطالعات فیزیولوژیکی در خرگوشها که توسط تیموتی بلیس و ترجه لومو (1973) انجام شد، شواهدی برای قانون هب ارائه کرد. بلیس و لومو دریافتند که تحریک آکسونهای مسیر پرفورنت منجر به افزایش طولانی مدت در میزان پتانسیلهای پس سیناپسی تحریکی (EPSPs) میشود. یعنی تحریک منجر به قدرت سیناپسی بیشتر در مسیر پرفورنت شد به طوری که وقتی آکسونها بعداً دوباره تحریک شدند، پاسخهای پس سیناپسی بزرگتری در سلولهای گرانول شکنج دندانه دار ایجاد شد. این پدیده به عنوان تقویت طولانی مدت (LTP) شناخته میشود (تقویت به معنای تشدید یا قوی تر کردن است) و کشف آن قانون هب را تأیید کرد.
LTP can be recorded by the combination of a stimulating electrode placed on the perforant pathway and at recording electrode placed in a granule cell of the dentate gyrus (Figure 9.40). A single pulse is presented, and the resulting EPSP is measured. The size of this first recording is the strength of the connection before the LTP is induced. Then the perforant pathway is stimulated with a burst of pulses; early studies used approximately 100 pulses/s, but more recent studies have used as few as 5 pulses/s. After LTP is induced, a single pulse is sent again, and the magnitude of the EPSP in the postsynaptic cell is measured. The magnitude of the EPSP increases after the LTP is induced, signaling the greater strength of the synaptic effect (Figure 9.40, red curve). A fascinating finding is that, when the pulses are presented slowly (as low-frequency pulses), the opposite effect- long-term depression (LTD)-develops (Figure 9.40, blue curve).
LTP را میتوان با ادغام دو الکترود ثبت کرد: یک الکترود تحریکی قرار داده شده در مسیر پرفورنت و یک الکترود ثبت قرار داه شده در سلول گرانولی شکنج دندانهدار (شکل 9.40). یک پالس منفرد ارائه میشود و EPSP حاصل اندازهگیری میشود. اندازه این ثبت، قدرت اتصال قبل از القای LTP است. سپس مسیر پرفورنت با پالسهای پی در پی تحریک میشود. در مطالعات اولیه تقریباً 100 پالس در ثانیه استفاده میشد، اما مطالعات جدیدتر از 5 پالس در ثانیه استفاده میکردند. پس از القای LTP، یک پالس منفرد دوباره ارسال میشود و بزرگی EPSP در سلول پس سیناپسی اندازه گیری میشود. بزرگی EPSP پس از القای LTP افزایش مییابد که نشاندهنده قدرت بیشتر اثر سیناپسی است (شکل 9.40، منحنی قرمز). یک یافته شگفتانگیز این است که وقتی پالسها به آرامی ارائه میشوند (بهعنوان پالسهای فرکانس پایین)، اثر معکوس – تضعیف طولانیمدت (LTD) – ایجاد میشود (شکل 9.40، منحنی آبی).

FIGURE 9.40 Stimulus and recording setup for the study of LTP In perforant pathways.
شکل 9.40 ساز و کار محرک و ثبت برای مطالعه LTP در مسیرهای پرفورنت.
The pattern of responses (in millivolts) before and after the induction of long-term potentiation (LTP) is shown as the red curve in the oscilloscope reading. The pattern of responses in long-term depression (LTD) is shown as the blue curve. PSPs = postsynaptic potentials.
الگوی پاسخها (بر حسب میلی ولت) قبل و بعد از القای تقویت طولانی مدت (LTP) به عنوان منحنی قرمز در خوانش اسیلوسکوپ نشان داده شده است. الگوی پاسخها در تضعیف طولانی مدت (LTD) به صورت منحنی آبی نشان داده شده است. PSPها = پتانسیلهای پس سیناپسی.
LTP was also found to occur in the other two excitatory projection pathways of the hippocampus: the mossy fibers and the Schaffer collaterals. The changes could last for hours in isolated slices of hippocampal tissue placed in dishes, and for weeks in living animals. LTP also takes place in other brain regions, including the amygdala, basal ganglia, cerebellum, and cortex-all involved with learning.
LTP همچنین در دو مسیر پروجکشن تحریکی دیگر هیپوکامپ رخ میدهد: فیبرهای خزهای و شاخههای جانبی شافر. این تغییرات میتواند ساعتها در برشهای جدا شده از بافت هیپوکامپ در ظروف و برای هفتهها در حیوانات زنده ادامه داشته باشد. LTP همچنین در سایر نواحی مغز، از جمله آمیگدال، عقدههای قاعدهای، مخچه و قشر مغز رخ میدهد که همگی درگیر یادگیری هستند.
HEBBIAN LEARNING By manipulating LTP in the CA1 neurons of the hippocampus, Bliss and Lømo found that if two weak inputs (W1 and W2) and a strong input (S1) are given to the same cell, then only when the weak (say, W1) and strong (S1) are active together is the weak input strengthened. Subsequently, if W2 and $1 are active together, then W1 is not affected by the LTP induced from the coactivation of W2 and S1. These findings sug- gest the following three properties of LTP in the CA1 synapses:
یادگیری هب: با دستکاری LTP در نورونهای CA1 هیپوکامپ، بلیس و لومو دریافتند که اگر دو ورودی ضعیف (W1 و W2) و یک ورودی قوی (S1) به یک سلول داده شود، تنها زمانی که ضعیف (مثلاً W1) و قوی (S1) با هم فعال هستند ورودی ضعیف تقویت شده است. پس از آن، اگر W2 و S1 با هم فعال باشند، W1 تحت تأثیر LTP ناشی از همفعال شدن W2 و S1 قرار نمیگیرد. این یافتهها سه ویژگی زیر را از LTP در سیناپسهای CA1 نشان میدهد:
1. Cooperativity. More than one input must be active at the same time.
1. همیاری. بیش از یک ورودی باید همزمان فعال باشد.
2. Associativity. Weak inputs are potentiated when co-occurring with stronger inputs.
2. شرکتپذیری. ورودیهای ضعیف زمانی که با ورودیهای قویتر اتفاق میافتند، تقویت میشوند.
3. Specificity. Only the stimulated synapse shows potentiation.
3. تخصص یافتگی. فقط سیناپس تحریک شده تقویت را نشان میدهد.
To account for these properties, University of Göteborg researchers Bengt Gustafsson and Holger Wigström (1988) found that for LTP to be produced, in addition to receiving simultaneous excitatory inputs the postsynaptic cells must be depolarized. In fact, LTP is reduced by inhibitory inputs to postsynaptic cells, which is what happens when habituation occurs. Moreover, when postsynaptic cells are hyperpolarized, LTP is prevented.
برای توضیح این ویژگیها، محققان دانشگاه گوتبورگ، بنگت گوستافسون و هولگر ویگستروم (1988) دریافتند که برای تولید LTP، علاوه بر دریافت ورودیهای تحریکی همزمان، سلولهای پس سیناپسی باید دپلاریزه شوند. در واقع، LTP توسط ورودیهای بازدارنده به سلولهای پس سیناپسی کاهش مییابد، این همان چیزی است که زمانی اتفاق میافتد که خوگیری اتفاق میافتد. علاوه بر این، هنگامی که سلولهای پس سیناپسی هیپرپلاریزه میشوند، از LTP جلوگیری میشود.
Conversely, when postsynaptic inhibition is prevented, LTP is facilitated. If an input that is normally not strong enough to induce LTP is paired with a depolarizing current to the postsynaptic cell, LTP can be induced. Thus, through associative LTP, weak pathways become strengthened and specifically associated with other pathways. This process supports learning in the way that Hebb proposed. The patterns of connectivity established by the strengthening and weakening of the synaptic connections are generally thought to encode information.
برعکس، وقتی از مهار پس سیناپسی جلوگیری میشود، LTP تسهیل میشود. اگر یک ورودی که معمولاً به اندازه کافی قوی برای القای LTP نیست، با یک جریان دپلاریز کننده به سلول پس سیناپسی جفت شود، میتوان LTP را القا کرد. بنابراین، مسیرهای ضعیف به کمک ویژگی شرکتپذیری LTP، تقویت شده و به طور خاص با مسیرهای دیگر مرتبط میشوند. این فرآیند از یادگیری، از روشی که هب پیشنهاد کرد حمایت میکند. به طور کلی تصور میشود که الگوهای اتصال ایجاد شده توسط تقویت و تضعیف اتصالات سیناپسی، اطلاعات را رمزگذاری میکنند.
What the code is remains a mystery. While some progress has been made at the level of the neuron, mechanisms at the subcellular level are even more complicated. Consider that each neuron has thousands of synapses, and each can be changed through multiple mechanisms, including the amount of neurotransmitter released; the number of receptors that are active can change, as can their properties, and new synapses can be formed. Whether any of these mechanisms contribute to memory is unknown.
اینکه کد چیست یک راز باقی مانده است. با وجودی که پیشرفتهایی در سطح نورون حاصل شده است، اما مکانیسمها در سطح درون سلولی پیچیده تر هستند. در نظر بگیرید که هر نورون دارای هزاران سیناپس است و هر کدام را میتوان از طریق مکانیسمهای متعددی از جمله میزان انتقال دهنده عصبی آزاد شده تغییر داد. تعداد گیرندههای فعال میتواند تغییر کند، همچنین خواص آنها تغییر میکند و سیناپسهای جدید میتوانند تشکیل شوند. اینکه آیا هر یک از این مکانیسمها به حافظه کمک میکند یا نه ناشناخته است.
What has been known since the 1960s is that long-term memory formation requires the expression of new genes. When a protein synthesis inhibitor (which inhibits the expression of new genes) is given to mice right after a training session, they are amnesic for the training experience. Yet recent research has shown that the memory remained in their brains! The implications of this finding are rocking the foundations of current theories about the cellular basis of learning (see Box 9.1).
آنچه از دهه 1960 شناخته شده است این است که شکل گیری حافظه بلندمدت مستلزم بیان ژنهای جدید است. هنگامی که یک مهارکننده سنتز پروتئین (که بیان ژنهای جدید را مهار میکند) بلافاصله پس از یک جلسه آموزشی به موشها داده میشود، آنها آن تجربه آموزشی را فراموش میکنند. با این حال تحقیقات اخیر نشان داده است که حافظه در مغز آنها باقی مانده است! پیامدهای این یافته پایههای تئوریهای کنونی در مورد اساس سلولی یادگیری را میلرزاند (به کادر 9.1 مراجعه کنید).
THE NMDA RECEPTOR The molecular mechanism that mediates LTP is fascinating. It depends on the neurotransmitter glutamate, the major excitatory transmitter in the hippocampus. Glutamate binds to two types of glutamate receptors. Normal synaptic transmissions are mediated by the AMPA (a-amino-3-hydroxyl-5-methyl-4-isoxazole propionate) receptor. LTP is initially mediated by the NMDA (N-methyl-D-aspartate) receptors (Figure 9.41), located on the dendritic spines of post-synaptic neurons. When the NMDA receptors of CA1 neurons are blocked with the chemical AP5 (2-amino-5-phosphonopentanoate), then LTP induction is prevented. Once LTP is established in these cells, however, AP5 treatment has no effect. Therefore, NMDA receptors are central to producing LTP, but not maintaining it. Maintenance of LTP probably depends on the AMPA receptors, although the mechanisms are not fully understood.
گیرنده NMDA: مکانیسم مولکولی که واسطه LTP میشود بسیار جذاب است. این به انتقال دهنده عصبی گلوتامات به عنوان انتقال دهنده اصلی تحریک کننده در هیپوکامپ بستگی دارد. گلوتامات به دو نوع گیرنده گلوتامات متصل میشود. انتقال سیناپسی معمولی توسط گیرنده AMPA (α-amino-3-hydroxyl-5-methyl-4-isoxazole propionate) انجام میشود. LTP در ابتدا با میانجیگری گیرندههای NMDA (N-methyl-D-aspartate) انجام میشود (شکل 9.41)، که بر روی خارهای دندریتی نورونهای پس سیناپسی قرار دارند. هنگامی که گیرندههای NMDA نورونهای CA1 با ماده شیمیایی AP5 (2-amino-5-phosphonopentanoate) مسدود میشوند، از القای LTP جلوگیری میشود. با این حال، هنگامی که LTP در این سلولها ایجاد شد، تیمار AP5 هیچ تاثیری ندارد. بنابراین، گیرندههای NMDA در تولید LTP نقش اساسی دارند، اما آن را حفظ نمیکنند. حفظ LTP احتمالا به گیرندههای AMPA بستگی دارد، اگرچه مکانیسمها به طور کامل شناخته نشده اند.
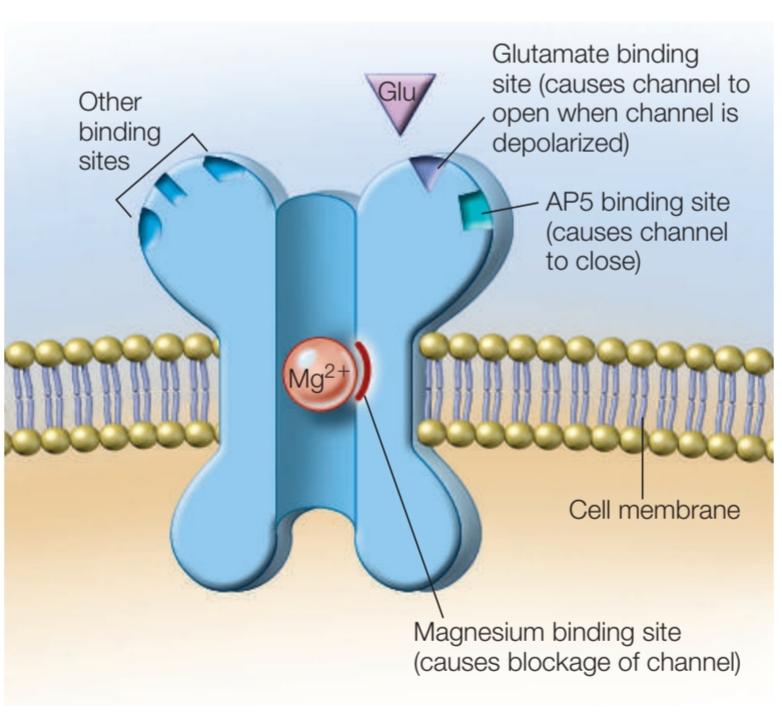
FIGURE 9.41 The NMDA receptor.
شکل 9.41 گیرنده NMDA
As this simplified cross-sectional schematic shows, the NMDA receptor is naturally blocked by Mg2+ ions. Unblocking (channel opening) occurs when the proteins that form the channel shift after glutamate binds to the glutamate binding site.
همانطور که این طرح مقطعی ساده نشان میدهد، گیرنده NMDA به طور طبیعی توسط یونهای +Mg2 مسدود میشود. رفع انسداد (باز شدن کانال) زمانی اتفاق میافتد که پروتئینهای تشکیلدهنده کانال به دلیل اتصال گلوتامات به محل اتصال گلوتامات جابجا شوند.
NMDA receptors are also blocked by magnesium ions (Mg2+), which prevent other ions from entering the postsynaptic cell. The Mg2+ ions can be ejected from the NMDA receptors only when the cell is depolarized. Thus, the ion channel opens only when two conditions are met: (a) the neurotransmitter glutamate binds to the receptors, and (b) the membrane is depolarized. These two conditions are another way of saying that the NMDA receptors are transmitter- and voltage-dependent (also called gated, Figure 9.42).
گیرندههای NMDA نیز توسط یونهای منیزیم (+Mg2) مسدود میشوند که از ورود یونهای دیگر به سلول پس سیناپسی جلوگیری میکند. تنها زمانی که سلول دپلاریزه شده است یونهای +Mg2 را میتوان از گیرندههای NMDA خارج کرد. بنابراین، کانال یونی تنها زمانی باز میشود که دو شرط برآورده شود: (الف) انتقال دهنده عصبی گلوتامات به گیرندهها متصل شود و (ب) غشاء دپلاریزه شده باشد. این دو شرط راه دیگری برای بیان این مطلب هستند که گیرندههای NMDA وابسته به ترانسمیتر و ولتاژ هستند (همچنین کانال دریچهدار نامیده میشوند، شکل 9.42).
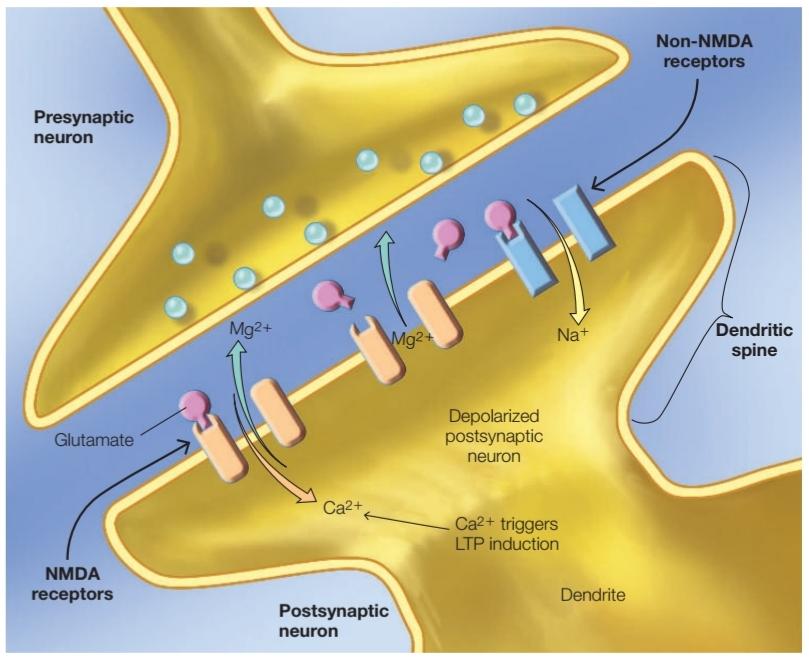
FIGURE 9.42 The role of Mg2+ and Ca2+ in the functioning of the NMDA receptor. See text for details.
شکل 9.42 نقش +Mg2 و +Ca2 در عملکرد گیرنده NMDA. برای جزئیات به متن مراجعه کنید.
The open ion channel allows Ca2+ ions to enter the postsynaptic cell. The effect of Ca2+ influx via the NMDA receptor is critical in the formation of LTP. The Ca2+ acts as an intracellular messenger conveying the signal, which changes enzyme activities that influence synaptic strength. Despite rapid advances in understanding the mechanisms of LTP at physiological and biochemical levels, the molecular mechanisms of synaptic strengthening in LTP are still the subject of extensive debate.
کانال یونی باز اجازه میدهد تا یونهای +Ca2 وارد سلول پس سیناپسی شوند. اثر هجوم +Ca2 از طریق گیرنده NMDA در تشکیل LTP حیاتی است. +Ca2 به عنوان یک پیام رسان درون سلولی سیگنال را منتقل میکند، که فعالیت آنزیمی را تغییر میدهد که بر قدرت سیناپسی تأثیر میگذارد. با وجود پیشرفتهای سریع در درک مکانیسمهای LTP در سطوح فیزیولوژیکی و بیوشیمیایی، مکانیسمهای مولکولی تقویت سیناپسی در LTP هنوز موضوع بحثهای گسترده است.
The synaptic changes that create a stronger synapse after LTP induction likely include presynaptic and postsynaptic mechanisms. One hypothesis is that LTP raises the sensitivity of postsynaptic AMPA glutamate receptors and prompts more glutamate to be released presynaptically. Or perhaps changes in the physical characteristics of the dendritic spines transmit EPSPs more effectively to the dendrites. Finally, via a message from the postsynaptic cell to the presynaptic cell, the efficiency of presynaptic neurotransmitter release is increased.
تغییرات سیناپسی که پس از القای LTP، سیناپس قویتری ایجاد میکند احتمالاً شامل مکانیسمهای پیشسیناپسی و پسسیناپسی میشود. یک فرضیه این است که LTP حساسیت گیرندههای گلوتاماتی AMPA پسسیناپسی را افزایش میدهد و نیز باعث میشود گلوتامات بیشتری به صورت پیشسیناپسی آزاد شود. یا شاید تغییرات در ویژگیهای فیزیکی خارهای دندریتی، EPSPها را به طور موثرتری به دندریتها منتقل میکند. در نهایت، از طریق یک پیام از سلول پسسیناپسی به سلول پیشسیناپسی، کارایی آزادسازی انتقال دهنده عصبی پیشسیناپسی افزایش مییابد.
BOX 9.1 | HOT SCIENCE
کادر ۹.۱ | علم داغ
i know you get there!
من میدانم که شما آنجا هستید!
Early in the 20th century, Richard Semon, a German evolutionary biologist, came up with the term engram to mean the enduring modification to the nervous system produced by an experience. The memory engram is the learned information stored in the brain-the chunk of neural tissue that must be reactivated in order to recall that information.
در اوایل قرن بیستم، ریچارد سمون، زیستشناس تکاملی آلمانی، اصطلاح انگرام را به معنای تغییر پایدار در سیستم عصبی ناشی از یک تجربه مطرح کرد. انگرام حافظه، اطلاعات آموخته شده ای است که در مغز ذخیره شده است – بخشی از بافت عصبی که باید دوباره فعال شود تا آن اطلاعات را به خاطر بیاورد.
Researchers have recently been able to isolate memory engram-bearing cells in the hippocampus. Tomás Ryan and his colleagues (2015) compared the physiological properties of engram cells with non-engram cells in the hip- pocampus of mice while they were learning the response to a stimulus. The researchers identified an increase of synaptic strength and dendritic spine density specifically in consolidated-memory engram cells. Next they used a recently developed technique to interfere with memory consolidation: They gave a protein synthesis inhibitor to the mice right after a training session, resulting in amnesia for the training. At the level of the cells, they found that the treatment that induced amnesia also abolished the increases of synaptic strength and dendritic spine density in the engram.
محققان اخیراً توانسته اند سلولهای حامل حافظه را در هیپوکامپ جدا کنند. توماس رایان و همکارانش (2015) خواص فیزیولوژیکی سلولهای انگرام را با سلولهای غیر انگرام در هیپوکامپ موشها در حالی که پاسخ به یک محرک را یاد میگرفتند، مقایسه کردند. محققان افزایش قدرت سیناپسی و تراکم خارهای دندریتی را به طور خاص در سلولهای انگرام حافظه تثبیت شده شناسایی کردند. سپس آنها از یک تکنیک اخیراً توسعه یافته برای تداخل در تثبیت حافظه استفاده کردند: آنها بلافاصله پس از یک جلسه آموزش به موشها یک مهارکننده سنتز پروتئین دادند که منجر به فراموشی تجربه آموزشی شد. آنها در سطح سلولی دریافتند که تیماری که باعث فراموشی میشود، افزایش قدرت سیناپسی و تراکم خارهای دندریتی در انگرام را نیز از بین میبرد.
Next, to test whether amnesia is caused by a consolidation failure, as many studies suggest, they optogenetically activated labeled CA3 engram cells in the two groups of trained mice: those that had been induced with amnesia for their training and those that had not. This is when they stumbled onto something unexpected: Directly activating the engram cells optogenetically produced the same behavior in both groups of mice; both groups demonstrated the trained response! The amnesic mice were able to perform the trained task even when their engram cells showed no increases in synaptic strength. The memory was not abolished even when the enhanced synaptic strength between the engram cells was!
در مرحله بعد، همانطور که بسیاری از مطالعات نشان میدهند، برای آزمایش اینکه آیا فراموشی ناشی از ناکارآمدی تثبیت است یا خیر، سلولهای CA3 انگرام برچسبگذاری شده اپتوژنتیکی را در دو گروه از موشهای آموزش دیده فعال کردند: موشهایی که تجربه آموزشی خود را فراموش کرده بودند و موشهایی که این تجربه را فراموش نکرده بودند. آنها در این موقع به چیزی غیرمنتظره، برخورد کردند: فعال کردن مستقیم سلولهای انگرام از نظر اپتوژنتیکی رفتار یکسانی را در هر دو گروه موش ایجاد کرد. هر دو گروه پاسخ آموزش دیده را نشان دادند! موشهای مبتلا به فراموشی حتی زمانی که سلولهای انگرام آنها هیچ افزایشی در قدرت سیناپسی نداشتند، میتوانستند این کار را انجام دهند. حتی زمانی که قدرت سیناپسی بین سلولهای انگرام افزایش یافته بود، حافظه از بین نرفت!
This finding confirms that synaptic strength appears to be necessary for memory retrieval. That is, no matter how memory is stored, when synapses are not operational, information cannot be extracted from cells (Queenan et al., 2017). Shockingly, however, synaptic strength, which has been the top contender as the mechanism for memory storage, has to be scrubbed (T. J. Ryan et al., 2015). So, how is memory stored if not by the synaptic changes? The search is on!
این یافته تأیید میکند که به نظر میرسد قدرت سیناپسی برای بازیابی حافظه ضروری است. یعنی مهم نیست که چقدر حافظه ذخیره میشود، وقتی سیناپسها عملیاتی نمیشوند، نمیتوان اطلاعات را از سلولها استخراج کرد (کوئینان و همکاران، 2017). با این حال، به طرز تکان دهنده ای، قدرت سیناپسی، که مدعی برتری به عنوان مکانیسم ذخیرهسازی حافظه بوده است، باید باطل شود (تی جی رایان و همکاران 2015). بنابراین، اگر ذخیرهسازی حافظه با تغییرات سیناپسی نباشد، حافظه چگونه ذخیره میشود؟ جستجو ادامه دارد!
🚀 با ما همراه شوید!
تازهترین مطالب و آموزشهای مغز و اعصاب را از دست ندهید. با فالو کردن کانال تلگرام آیندهنگاران مغز، از ما حمایت کنید!

 ورود/ ثبت نام با جیمیل
ورود/ ثبت نام با جیمیل
