کتاب الکترونیکی «پرسشهای چند گزینهای علوم اعصاب شامل تمامیمباحث نوروآناتومی»
انتشارات: موسسه آموزشی تالیفی ارشدان
تعداد صفحه: ۸۴
این کتاب شامل تمامیپرسشهای نوروآناتومیدکتری علوم اعصاب از سال ۱۳۸۷ تا سال ۱۴۰۰ همراه پاسخ کلیدی پرسشها
آزمون شامل ۲۵ پرسش است.
سیستم عصبی انسان پیچیده ترین و همه کاره ترین دستاورد تکامل است. سیستم عصبی همه جانوران برای تشخیص تغییرات در محیطهای خارجی و داخلی و ایجاد پاسخهای مناسب در عضلات، اندامها و غدد عمل میکند. با صعود به مقیاس تکاملی، علاوه بر این، ظرفیت فزاینده ای برای “عملکردهای بالاتر” سیستم عصبی مانند یادگیری، حافظه، شناخت و در نهایت خودآگاهی، عقل و شخصیت وجود دارد. اساس آناتومیکی، فیزیولوژیکی، بیوشیمیایی و مولکولی عملکرد عصبی حوزههای فعالیت تحقیقاتی شدید در علوم پایه و بالینی است. در حالی که اطلاعات زیادی در مورد نحوه عملکرد سیستم عصبی وجود دارد، هنوز چیزهای زیادی در انتظار روشن شدن هستند.
سیستم عصبی میتواند توسط ناهنجاریهای ارثی یا رشدی، فرآیندهای بیماری و آسیبهای تروماتیک آسیب ببیند. بنابراین، پیشگیری، تشخیص و درمان اختلالات عصبی از اهمیت اجتماعی و اقتصادی بسیار بالایی برخوردار است. درک نوروآناتومی و ارتباط آن با عملکرد و اختلال در عملکرد معاصر علوم اعصاب بالینی و چشم انداز پیشرفتهای آینده در پیشگیری و درمان اختلالات عصبی اساسی است.
نورونها و نوروگلیا
واحد اصلی ساختاری و عملکردی سیستم عصبی سلول عصبی یا نورون است (شکل ۱.۱، ۱.۲)، که سیستم عصبی انسان حدود ۱۰۱۰ تخمین زده میشود. وظایف نورون دریافت و ادغام اطلاعات دریافتی از گیرندههای حسی یا سایر نورونها و انتقال اطلاعات به سایر نورونها یا اندامهای موثر است. ساختار عصبی برای انجام این وظایف بسیار تخصصی است. هر نورون یک موجودیت جداگانه با یک غشای سلولی محدود کننده است. اطلاعات بین نورونها در نواحی تخصصی به نام سیناپس که در آن غشای سلولهای مجاور در کنار هم قرار دارند، منتقل میشود (شکل ۱.۱).
 شکل ۱.۱ نمایش شماتیک ساختار اصلی نورون و سیناپس.
شکل ۱.۱ نمایش شماتیک ساختار اصلی نورون و سیناپس.

شکل ۱.۲ بازسازی سه بعدی شبه رنگی یک نورون از هیپوکامپ، تصویربرداری شده توسط میکروسکوپ اسکن لیزری کانفوکال. یکی از فرآیندهای پایه نورون، آکسون است.
(از دکتر RA McKinney، موسسه تحقیقات مغز، دانشگاه زوریخ، سوئیس.)
تنوع گسترده ای در شکل و اندازه نورونها در بخشهای مختلف سیستم عصبی وجود دارد، اما همه ویژگیهای مشترک خاصی دارند. یک جسم سلولی وجود دارد که تعداد متغیری از فرآیندهای انشعاب از آن خارج میشود. اکثر این فرآیندها از نظر عملکرد پذیرا هستند و به عنوان دندریت شناخته میشوند. آنها دارای تخصصهای سیناپسی هستند، گاهی اوقات هزاران مورد، که از طریق آن اطلاعات را از سایر سلولهای عصبی دریافت میکنند. در نورونهای حسی، دندریتها ممکن است برای تشخیص تغییرات در محیط خارجی یا داخلی تخصصیتر شوند. یکی از فرآیندهای خروج از بدن سلولی آکسون نام دارد (فیبر عصبی) و این اطلاعات را از بدن سلولی دور میکند. طول آکسونها بسیار متغیر است و ممکن است به چندین شاخه یا وثیقه تقسیم شوند که از طریق آنها میتوان اطلاعات را به چندین مقصد مختلف به طور همزمان توزیع کرد. در انتهای آکسون، تخصصهای سیناپسی به نام پایانههای عصبی (پایانهای پیش سیناپسی، بوتونهای انتهایی) رخ میدهد. از اینها، اطلاعات به دندریت سایر نورونها منتقل میشود.
اطلاعات درون نورونها با تغییرات انرژی الکتریکی کدگذاری میشود. نورون در حالت سکون دارای پتانسیل الکتریکی (پتانسیل استراحت) در سراسر غشای خود در حد ۶۰ تا ۷۰ میلی ولت (mV) است که داخل آن نسبت به بیرون منفی است. هنگامیکه یک نورون بالاتر از یک سطح آستانه خاص تحریک یا برانگیخته میشود، یک معکوس مختصر از قطبیت پتانسیل غشایی آن وجود دارد که به آن پتانسیل عمل میگویند. پتانسیلهای عمل در آکسون منتشر میشوند و به پایانههای عصبی حمله میکنند. انتقال اطلاعات بین نورونها تقریباً همیشه با روشهای شیمیایی و نه الکتریکی اتفاق میافتد. حمله به پایانههای عصبی توسط یک پتانسیل عمل باعث آزاد شدن عوامل شیمیایی خاصی میشود که در وزیکولهای سیناپسی ذخیره میشوند. در پایان پیش سیناپسی این مواد شیمیایی به عنوان انتقال دهندههای عصبی شناخته میشوند و در شکاف باریک بین غشاهای پیش و پس سیناپسی پخش میشوند تا به گیرندههای سلول پس سیناپسی متصل شوند و باعث ایجاد تغییرات در پتانسیل غشاء شوند. این تغییر ممکن است یا برای دپولاریز کردن غشاء، در نتیجه حرکت به سمت آستانه تولید پتانسیلهای عمل، یا به سمت هیپرپلاریزه شدن و در نتیجه تثبیت سلول باشد.
دیگر اجزای سلولی اصلی سیستم عصبی، سلولهای نوروگلیال یا گلیا هستند که از نظر قدر از نورونها بیشتر هستند. بر خلاف نورونها، نوروگلیا نقش مستقیمیدر پردازش اطلاعات ندارند، اما تعدادی از نقشهای کمکی ضروری برای عملکرد طبیعی سلولهای عصبی را ایفا میکنند. سه نوع اصلی سلول نوروگلیال شناخته شده است:
۱. الیگودندروگلیا (الیگودندروسیت)، که غلاف میلین را تشکیل میدهد که بسیاری از آکسونهای عصبی را احاطه کرده است (شکل ۱.۱)، که باعث افزایش سرعت انتقال پتانسیلهای عمل میشود.
۲. آستروگلیا (آستروسیتها)، که تصور میشود یک مانع نفوذ پذیر انتخابی بین سیستم گردش خون و نورونهای مغز و نخاع ایجاد میکنند. این به عنوان”سد خونی مغزی“شناخته میشود و عملکرد محافظتی دارد.
۳. میکروگلیا که نقش فاگوسیتیزی در پاسخ بافت محلی به آسیب سیستم عصبی دارند.
سیستم عصبی مرکزی و محیطی
سیستم عصبی (شکل ۱.۳) به سیستم عصبی مرکزی (CNS) و سیستم عصبی محیطی (PNS) تقسیم میشود. سیستم عصبی مرکزی از مغز و نخاع تشکیل شده است، به ترتیب در محافظت از جمجمه و ستون مهره قرار دارد. این پیچیده ترین بخش سیستم عصبی است. این شامل اکثر اجسام سلولهای عصبی و اتصالات سیناپسی است. سیستم عصبی محیطی پیوند بین CNS و ساختارهای اطراف بدن را تشکیل میدهد که از آن اطلاعات حسی دریافت میکند و تکانههای کنترلی را به آن میفرستد. سیستم عصبی محیطی شامل اعصاب متصل به مغز و نخاع (اعصاب جمجمه ای و نخاعی) و شاخههای آنها در بدن است. اعصاب نخاعی که به اندامهای فوقانی یا تحتانی خدمت میکنند به ترتیب به هم میپیوندند و شبکه بازویی یا کمری را تشکیل میدهند که در آن رشتهها به نامهای مختلف توزیع میشوند. اعصاب محیطی. PNS همچنین شامل گروههایی از سلولهای عصبی محیطی است که در ساختارهایی به نام گانگلیون تجمع یافته اند.
 شکل ۱.۳ سیستم عصبی مرکزی و محیطی.
شکل ۱.۳ سیستم عصبی مرکزی و محیطی.
سامانه ی عصبی خودمختار
نورونهایی که تغییرات را در احشاء تشخیص داده و فعالیت آن را کنترل میکنند، در مجموع به عنوان سیستم عصبی خودمختار (ANS) شناخته میشوند. اجزای آن در هر دو سیستم عصبی مرکزی و محیطی وجود دارد. سیستم عصبی خودمختار از نظر تشریحی و عملکردی به دو بخش مجزا تقسیم میشود، یعنی بخشهای سمپاتیک و پاراسمپاتیک، که عموماً اثرات متضاد (متضاد) بر ساختارهایی که عصب میکنند دارند. سیستم عصبی خودمختار عضله صاف، عضله قلب و غدد ترشحی را عصب دهی میکند. این بخش مهمی از مکانیسمهای هموستاتیک است که محیط داخلی بدن را کنترل میکند.
اجزاء و سازماندهی سیستم عصبی
![]() واحد ساختاری و عملکردی سیستم عصبی سلول عصبی یا نورون است. نورونها پتانسیل غشای استراحتی در حدود ۷۰- میلی ولت دارند.
واحد ساختاری و عملکردی سیستم عصبی سلول عصبی یا نورون است. نورونها پتانسیل غشای استراحتی در حدود ۷۰- میلی ولت دارند.
![]() یک نورون اطلاعات را عمدتاً از طریق دندریتهای خود دریافت میکند و آن را با پتانسیلهای عمل، که توسط آکسون از بدن سلولی دور میشوند، منتقل میکند.
یک نورون اطلاعات را عمدتاً از طریق دندریتهای خود دریافت میکند و آن را با پتانسیلهای عمل، که توسط آکسون از بدن سلولی دور میشوند، منتقل میکند.
![]() اطلاعات بین نورونها در سیناپسها با آزادسازی انتقال دهندههای عصبی از پایانههای پیش سیناپسی منتقل میشود. اینها روی گیرندههای غشای پس سیناپسی عمل میکنند و باعث دپلاریزاسیون یا هیپرپولاریزاسیون سلول پس سیناپسی میشوند.
اطلاعات بین نورونها در سیناپسها با آزادسازی انتقال دهندههای عصبی از پایانههای پیش سیناپسی منتقل میشود. اینها روی گیرندههای غشای پس سیناپسی عمل میکنند و باعث دپلاریزاسیون یا هیپرپولاریزاسیون سلول پس سیناپسی میشوند.
![]() تعداد سلولهای نوروگلیال از سلولهای عصبی بیشتر است، اما نقشهای فرعی دارند و مستقیماً در پردازش اطلاعات نقشی ندارند.
تعداد سلولهای نوروگلیال از سلولهای عصبی بیشتر است، اما نقشهای فرعی دارند و مستقیماً در پردازش اطلاعات نقشی ندارند.
![]() الیگودندروسیتها غلاف میلین را تشکیل میدهند که آکسونها را احاطه کرده و سرعت هدایت آنها را افزایش میدهد.
الیگودندروسیتها غلاف میلین را تشکیل میدهند که آکسونها را احاطه کرده و سرعت هدایت آنها را افزایش میدهد.
![]() آستروسیتها ممکن است سد خونی مغزی را تشکیل دهند.
آستروسیتها ممکن است سد خونی مغزی را تشکیل دهند.
![]() هنگامیکه سیستم عصبی آسیب میبیند، میکروگلیا عملکرد فاگوسیتی دارد.
هنگامیکه سیستم عصبی آسیب میبیند، میکروگلیا عملکرد فاگوسیتی دارد.
![]() سیستم عصبی به سیستم عصبی مرکزی (CNS) که شامل مغز و نخاع است و سیستم عصبی محیطی (PNS) که از اعصاب جمجمه ای و نخاعی و شاخههای آنها تشکیل شده است، تقسیم میشود.
سیستم عصبی به سیستم عصبی مرکزی (CNS) که شامل مغز و نخاع است و سیستم عصبی محیطی (PNS) که از اعصاب جمجمه ای و نخاعی و شاخههای آنها تشکیل شده است، تقسیم میشود.
![]() سیستم عصبی خودمختار (ANS) ساختارهای احشایی را عصب میکند و در هموستاز محیط داخلی مهم است.
سیستم عصبی خودمختار (ANS) ساختارهای احشایی را عصب میکند و در هموستاز محیط داخلی مهم است.
![]() نورونهای منفرد ممکن است به عنوان آوران یا وابران با توجه به CNS یا به عنوان نورونهای داخلی تعریف شوند.
نورونهای منفرد ممکن است به عنوان آوران یا وابران با توجه به CNS یا به عنوان نورونهای داخلی تعریف شوند.
![]() در CNS، نواحی غنی از سلولهای عصبی یا رشتههای عصبی به ترتیب ماده خاکستری و سفید را تشکیل میدهند.
در CNS، نواحی غنی از سلولهای عصبی یا رشتههای عصبی به ترتیب ماده خاکستری و سفید را تشکیل میدهند.
![]() خوشههایی از اجسام سلولی با عملکردهای مشابه به عنوان هسته شناخته میشوند.
خوشههایی از اجسام سلولی با عملکردهای مشابه به عنوان هسته شناخته میشوند.
![]() مسیرها یا مسیرهای رشتههای عصبی مناطق دوردست را به هم متصل میکنند.
مسیرها یا مسیرهای رشتههای عصبی مناطق دوردست را به هم متصل میکنند.
![]() به طور کلی، مسیرهای حرکتی حسی صعودی و نزولی در CNS در طول مسیر خود مورد بحث قرار میگیرند، به طوری که هر طرف مغز از نظر عملکردی با نیمه طرف مقابل بدن مرتبط است.
به طور کلی، مسیرهای حرکتی حسی صعودی و نزولی در CNS در طول مسیر خود مورد بحث قرار میگیرند، به طوری که هر طرف مغز از نظر عملکردی با نیمه طرف مقابل بدن مرتبط است.
نورونهای آوران، نورونهای وابران و نورونهای داخلی
سلولهای عصبی که اطلاعات را از گیرندههای محیطی به CNS حمل میکنند به عنوان نورونهای آوران نامیده میشوند (شکل ۱.۴). اگر اطلاعاتی که آنها حمل میکنند در نهایت به سطح خودآگاه برسد، آنها را نورونهای حسی نیز مینامند. نورونهای وابران تکانه ها را از CNS دور میکنند و اگر عضله اسکلتی را عصب دهی کنند تا باعث حرکت شوند، به آنها نورونهای حرکتی نیز گفته میشود. با این حال، اکثریت قریب به اتفاق نورونها به طور کامل در CNS قرار دارند و به آنها نورونهای داخلی میگویند (گاهی اوقات از اصطلاحات جایگزین internuncial یا relay neurones استفاده میشود). اصطلاحات «آوران» و «وابران» معمولاً برای نشان دادن قطبیت برآمدگیها به و از ساختارهای درون CNS استفاده میشوند، حتی اگر برجستگیها به طور کامل در مغز و نخاع وجود داشته باشند. به عنوان مثال، برآمدگیها به قشر مغز و از آن به ترتیب آورانهای قشری و وابران نامیده میشوند.
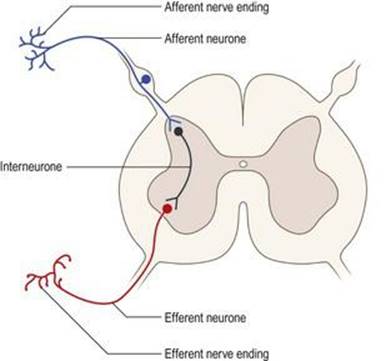 شکل ۱.۴ آرایش کلی آوران، وابران و نورونهای داخلی.
شکل ۱.۴ آرایش کلی آوران، وابران و نورونهای داخلی.
ماده خاکستری و سفید، هستهها و مجاری
CNS یک ساختار بسیار ناهمگن از نظر توزیع اجسام سلولهای عصبی و فرآیندهای آنها است (شکل ۱.۵). برخی از نواحی از نظر بدن سلولهای عصبی (مثلاً بخش مرکزی نخاع و سطح نیمکره مغز) نسبتاً غنی شده اند و به آنها ماده خاکستری میگویند. سایر نواحی عمدتاً شامل فرآیندهای عصبی (معمولاً آکسون) هستند. اینها اغلب میلین دار هستند (پوشش در میلین)، که رنگ کم رنگ تری ایجاد میکند – از این رو ماده سفید را به آن میگویند.
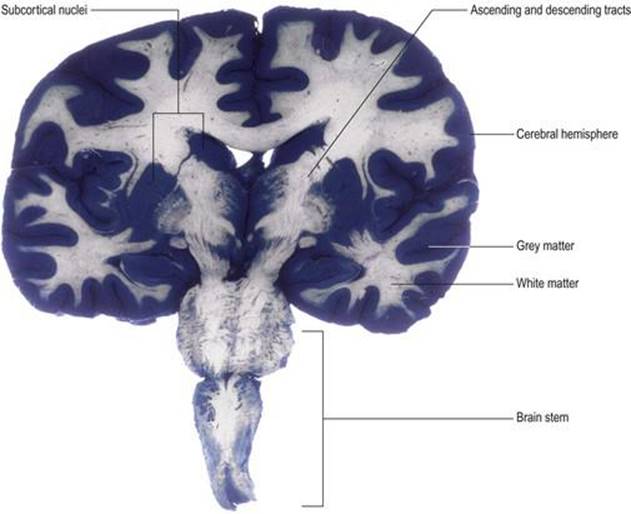 شکل ۱.۵ بخش تاجی از طریق مغز که توزیع ماده خاکستری و سفید را نشان میدهد. این بخش با تکنیک مولیگان رنگ آمیزی شده است که ماده خاکستری را آبی رنگ میکند و ماده سفید را نسبتاً بدون رنگ میگذارد.
شکل ۱.۵ بخش تاجی از طریق مغز که توزیع ماده خاکستری و سفید را نشان میدهد. این بخش با تکنیک مولیگان رنگ آمیزی شده است که ماده خاکستری را آبی رنگ میکند و ماده سفید را نسبتاً بدون رنگ میگذارد.
اجسام سلولهای عصبی با اتصالات و عملکردهای تشریحی مشابه (مثلاً نورونهای حرکتی که گروهی از عضلات مرتبط را عصب دهی میکنند) تمایل دارند در کنار هم در گروههایی به نام هسته قرار گیرند . به طور مشابه، فرآیندهای عصبی که اتصالات و عملکردهای مشترک را به اشتراک میگذارند، تمایل دارند مسیر مشابهی را دنبال کنند و در مسیرها یا مجاری اجرا میشوند (شکل ۱.۵ و شکل ۱.۱۹ را ببینید).
بحث از مسیرهای حسی و حرکتی
این یک اصل کلی از سازماندهی CNS است که مسیرهایی که اطلاعات حسی را به سطح خودآگاه (نیمکره مغزی) منتقل میکند، از یک طرف CNS به سمت دیگر متقاطع یا جدا میشود. همین امر در مورد مسیرهای نزولی از نیمکره مغزی که حرکت را کنترل میکنند نیز صادق است. بنابراین، به طور کلی، هر نیمکره مغزی احساسات را از طرف مقابل (مقابل) بدن درک میکند و حرکات آن را کنترل میکند.
توسعه سیستم عصبی مرکزی
با آغاز هفته دوم رشد جنینی انسان، سه لایه سلول زاینده ایجاد میشود: اکتودرم، مزودرم و اندودرم. متعاقباً، هر کدام باعث ایجاد بافتها و اندامهای خاصی در بزرگسالان میشوند. اکتودرم باعث ایجاد پوست و سیستم عصبی میشود. مزودرم بافتهای اسکلتی، عضلانی و همبند را تشکیل میدهد. اندودرم مجاری گوارشی، تنفسی و دستگاه تناسلی را ایجاد میکند. فرآیند تشکیل سیستم عصبی جنینی به عنوان نورولاسیون شناخته میشود. در هفته سوم رشد جنینی، اکتودرم خط میانی پشتی برای تشکیل صفحه عصبی ضخیم میشود (شکل ۱.۶، ۱.۷). حاشیههای جانبی صفحه عصبی بالا رفته و تشکیل میشود چینهای عصبی در دو طرف یک فرورفتگی طولی خط وسط، شیار عصبی. سپس چینهای عصبی به هم متصل میشوند و به هم جوش میخورند، بنابراین شیار عصبی را میبندند و لوله عصبی را ایجاد میکنند. برخی از سلولها از نوک چینهای عصبی جدا میشوند و گروههایی را تشکیل میدهند که پشتی جانبی لوله عصبی قرار دارند. اینها به عنوان تاجهای عصبی شناخته میشوند. تشکیل لوله عصبی تقریباً در اواسط هفته چهارم رشد جنینی کامل میشود.
 شکل ۱.۶ میکروگرافهای الکترونی روبشی مقاطع عرضی از طریق اکتودرم پشتی جنین جوجه که مراحل (از بالا به پایین) تشکیل لوله عصبی را نشان میدهد (۱۴۰×).
شکل ۱.۶ میکروگرافهای الکترونی روبشی مقاطع عرضی از طریق اکتودرم پشتی جنین جوجه که مراحل (از بالا به پایین) تشکیل لوله عصبی را نشان میدهد (۱۴۰×).
(با اجازه دکتر GC Schoenwolf، بخش نوروبیولوژی و آناتومی، دانشکده پزشکی دانشگاه یوتا، سالت لیک سیتی، ایالات متحده آمریکا.)

شکل ۱.۷ نمایش شماتیک از تشکیل لوله عصبی از اکتودرم جنینی.
رشد عظیم، اعوجاج و تمایز سلولی در طول تبدیل بعدی لوله عصبی به CNS بالغ رخ میدهد. این حداکثر در قسمت منقاری است که در مغز رشد میکند و قسمت دمیبه نخاع تبدیل میشود. حفره مرکزی درون لوله عصبی به کانال مرکزی نخاع و بطنهای مغز تبدیل میشود. تاجهای عصبی عقدههای حسی اعصاب نخاعی و جمجمه ای و همچنین عقدههای اتونومیک را تشکیل میدهند.
با ادامه رشد، یک شیار طولی به نام sulcus limitans در سطح داخلی دیوارههای جانبی نخاع جنینی و قسمت دمیمغز ظاهر میشود (شکل ۱.8A). گروهبندی سلولهای پشتی و شکمیکه بدین ترتیب مشخص شدهاند، به ترتیب به عنوان صفحه آلار و صفحه پایه شناخته میشوند. سلولهای عصبی که در صفحه آلار رشد میکنند، عمدتاً عملکردهای حسی دارند، در حالی که سلولهای موجود در صفحه پایه عمدتاً حرکتی هستند. توسعه بیشتر همچنین باعث تمایز ماده خاکستری و سفید میشود. ماده خاکستری در مرکز اطراف کانال مرکزی قرار دارد و ماده سفید یک پوشش بیرونی را تشکیل میدهد. این الگوی اصلی رشد هنوز هم به راحتی در نخاع بزرگسالان قابل تشخیص است.شکل ۱.8B).
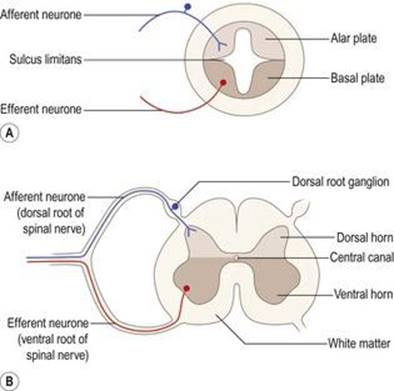 شکل ۱.۸ نمایش شماتیک مقاطع عرضی از طریق (A) لوله عصبی در حال رشد و (B) نخاع بالغ. اتصالات عصبی با ساختارهای محیطی تنها در یک طرف نشان داده شده است.
شکل ۱.۸ نمایش شماتیک مقاطع عرضی از طریق (A) لوله عصبی در حال رشد و (B) نخاع بالغ. اتصالات عصبی با ساختارهای محیطی تنها در یک طرف نشان داده شده است.
در طول رشد جنینی، بخش منقاری لوله عصبی تحت تمایز و رشد گسترده قرار میگیرد تا مغز را تشکیل دهد (شکل ۱.۹). در حدود هفته پنجم، سه وزیکول مغزی اولیه را میتوان شناسایی کرد: پروسنفالون (جلو مغز)، مزانسفالون (مغز میانی) و رومبنسفالون (مغز عقبی). محور طولی CNS در حال رشد (نوراکسیس) مستقیم نمیماند بلکه توسط یک خمیدگی مغز میانی یا سفالیک خم میشود که در محل اتصال مغز میانی و جلو مغز و یک خم شدن گردنی بین مغز و نخاع ایجاد میشود.
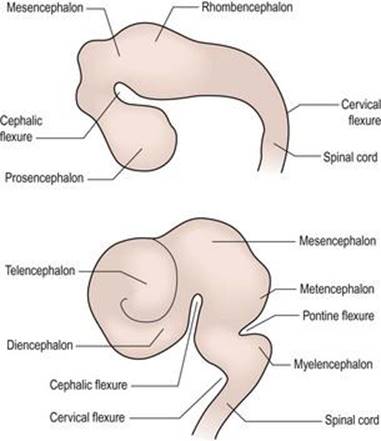 شکل ۱.۹ رشد اولیه مغز که (الف) وزیکولهای اولیه مغز را در حدود ۴-۵ هفته و (B) وزیکولهای مغزی ثانویه را در حدود ۷-۸ هفته نشان میدهد.
شکل ۱.۹ رشد اولیه مغز که (الف) وزیکولهای اولیه مغز را در حدود ۴-۵ هفته و (B) وزیکولهای مغزی ثانویه را در حدود ۷-۸ هفته نشان میدهد.
در هفته هفتم، تمایز بیشتر، پنج وزیکول مغزی ثانویه را که با تقسیم پروسنفالون به تلانسفالن و دیانسفالون و تقسیم رومبنسفالون به متانسفالون و میلنسفالون تولید میشوند، متمایز میکند. محل اتصال بین دومیبا یک خم اضافی در نوراکسیس مشخص میشود که خمش پونتین نامیده میشود.
برخی از نامهای زیربخشهای جنینشناسی مغز معمولاً برای اهداف توصیفی استفاده میشوند و بنابراین، دانستن قسمتهایی از مغز بالغ که متعاقباً در آن رشد میکنند مفید است (جدول ۱.۱). از میان سه بخش اصلی مغز، پیش مغزی یا پیش مغزی به مراتب بزرگترین است. به آن مغز نیز میگویند. در داخل مخ، تلانسفالن بیشترین رشد بیشتری را تجربه میکند و باعث ایجاد دو نیمکره مغزی میشود. اینها از یک لایه بیرونی ماده خاکستری (قشر مخ) و یک توده درونی ماده سفید تشکیل شده اند که در داخل آن گروههای مختلفی از هستهها مدفون شده اند (بزرگترین آنها جسم مخطط است ). دیانسفالون عمدتاً از تالاموس که شامل گروههای سلولی متعددی است و ارتباط نزدیکی با قشر مغز دارد. مزانسفالون یا مغز میانی نسبتاً تمایز نیافته است (هنوز یک حفره لوله مانند مرکزی که توسط ماده خاکستری احاطه شده است را حفظ میکند). متانسفالون به سمت پونز و مخچه پوشاننده رشد میکند، در حالی که میلنسفالون بصل النخاع (بصل النخاع) را تشکیل میدهد. مدولا، پونز و مغز میانی در مجموع به عنوان ساقه مغز شناخته میشوند (شکل ۱.۱۰).
جدول ۱.۱ رشد جنینی مغز
| وزیکولهای مغزی اولیه | وزیکولهای مغزی ثانویه | مشتقات در مغز بالغ |
| پروسنفالون (پیش مغز) | تلنسفالون | نیمکره مغزی |
| دیانسفالون | تالاموس | |
| مزانسفالون (مغز میانی) | مزانسفالون | مغز میانی |
| Rhombencephalon (مغز عقبی) | متانسفالون | پونز، مخچه |
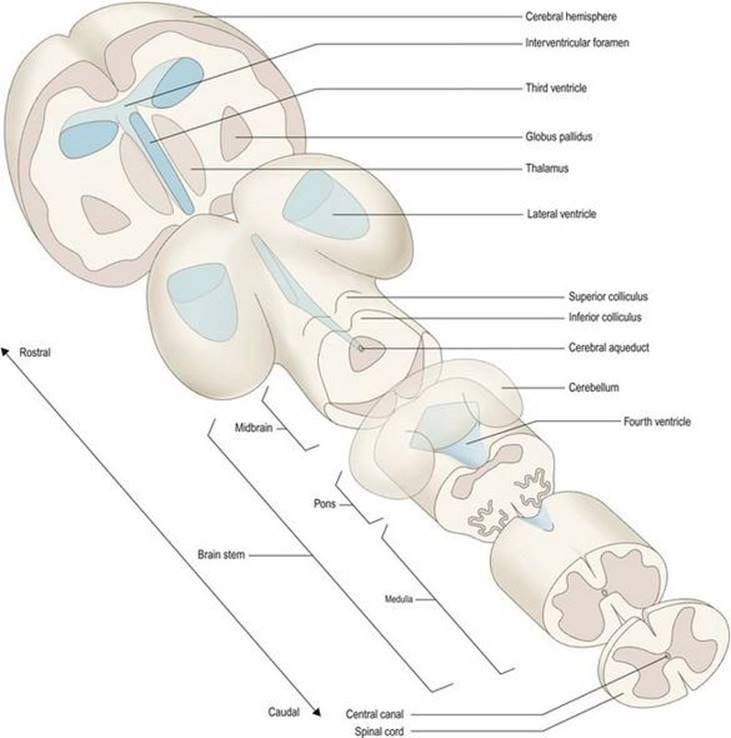 شکل ۱۰-۱ نمایش شماتیک زیربخشهای اصلی و نشانههای مغز.
شکل ۱۰-۱ نمایش شماتیک زیربخشهای اصلی و نشانههای مغز.
همانطور که مغز رشد میکند، حفره مرکزی آن نیز دستخوش تغییرات قابل توجهی در اندازه و شکل میشود و سیستمیاز حفرهها یا بطنها را تشکیل میدهد (شکل ۱.۱۰ و شکل ۱.۱۸ را ببینید) که حاوی مایع مغزی نخاعی (CSF) است.
شباهتهایی بین رشد جنینی مغز و تغییرات عمده ای که مغز در طی صعود به مقیاس فیلوژنتیک یا تکاملی از حیوانات ساده به پیچیده تر متحمل شده است ترسیم شده است. در حالی که این مطمئناً یک سادهسازی بیش از حد است، این مفهوم دارای شایستگی معرفی برخی از بخشهای اصلی مغز، و روابط آنها با یکدیگر، به شیوهای گرافیکی و به یاد ماندنی است (شکل ۱.۱۰).
سادهترین جانوران وتردار (مثلاً آمفیوکسوس)، که مهرهداران از آن تکامل یافتهاند، دارای طناب عصبی لولهای پشتی هستند که یادآور لوله عصبی جنین پستانداران در حال رشد است. در طول فیلوژنی، انتهای منقاری سیستم عصبی لولهای دستخوش تغییرات و تغییرات عظیمیشده است. در نتیجه، مغز انسان بالغ شباهت آشکار کمیبه اجداد تکاملی خود دارد.
تخصص منطقه ای موضوع مهمیدر تکامل مغز بوده است و این امر به ویژه در رابطه با حواس و کنترل حرکت آشکار است. مدتها پیش در فیلوژنی، مراکزی که به این عملکردها اختصاص یافته بودند، بهعنوان انبساط یا رشد از جنبه پشتی مغز لولهای ساده ایجاد شدند (شکل ۱.۱۰) از نظر شکل، آنها شامل یک قشر بیرونی از اجسام سلولهای عصبی با یک هسته زیرین رشتههای عصبی بودند. مراکز جفت دو طرفه در رابطه با حواس بویایی، بینایی و شنوایی ایجاد شدند و یک مرکز متقارن خط وسط در ارتباط با عملکرد دهلیزی و حفظ تعادل ایجاد شد. هر یک از این مراکز تحت تغییرات تکاملی بعدی قرار گرفتند، اما این بیشتر در قسمت منقاری،”بویایی”مغز، که به نیمکرههای عظیم مغزی تبدیل شد، مشهود بود (شکل ۱.۱۱، ۱.۱۲). در طول این فرآیند پروانسفالیزه شدن، نیمکرههای مغز نقش اجرایی در بسیاری از زمینههای عملکرد مغز به عهده گرفتند. به عنوان مثال، بالاترین سطح برای درک و تفسیر ورودی از تمام روشهای حسی، در نهایت در سطح قشر مغز نیمکرههای مغزی، و بالاترین سطح برای کنترل حرکتی، محلی شد. این امر با این واقعیت منعکس میشود که تنها بخش کوچکی از نیمکره مغزی انسان بالغ به عملکرد بویایی اختصاص دارد.
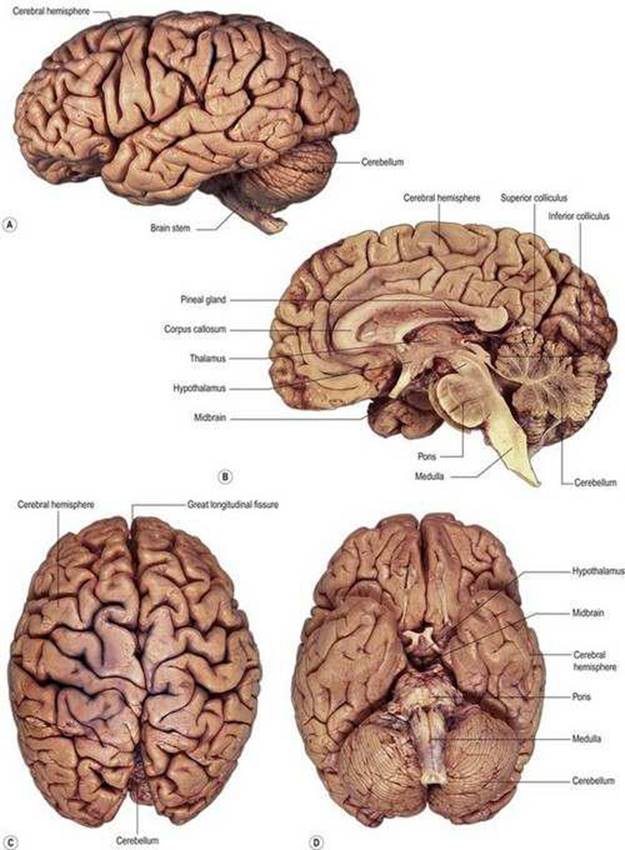 شکل ۱.۱۱ عکسهای مغز. الف) جنبه جانبی؛ (ب) بخش ساژیتال میانه. (ج) جنبه پشتی؛ (د) جنبه شکمی.
شکل ۱.۱۱ عکسهای مغز. الف) جنبه جانبی؛ (ب) بخش ساژیتال میانه. (ج) جنبه پشتی؛ (د) جنبه شکمی.


شکل ۱.۱۲ تقسیمات فرعی اصلی و برخی از نشانههای مهم در مغز بالغ. (الف) بخش ساژیتال میانه؛ (ب) جنبه شکمی. اعصاب اعصاب جمجمه ای با رنگ زرد نشان داده شده اند.
فرآیند پرونسفالیزاسیون به این معنی بود که سایر مراکز به تدریج تابع نیمکرههای مغزی شدند. برای مثال، آنهایی که برای بینایی و شنوایی هستند، رشد نسبتا کمیداشتند و عملکردهای بازتابی و خودکار را در مغز انسان انجام میدهند. با این حال، هنوز هم ممکن است به عنوان چهار تورم کوچک در سطح پشتی مغز میانی شناسایی شوند: corpora quadrigemina یا کولیکولهای فوقانی و تحتانی (شکلهای ۱.۱۰ – ۱.۱۲). مرکز حرکتی نزدیک انتهای دمیمغز به مخچه توسعه یافته است (شکلهای ۱.۱۰ – ۱.۱۲) که نقش اصلی را در حفظ تعادل و هماهنگی حرکت حفظ میکند.
![]() ناهنجاریهای رشدی
ناهنجاریهای رشدی
اختلالات رشد، رشد طبیعی و سازمان ساختاری نخاع و مغز را مختل میکند. از آنجایی که سیستم عصبی از اکتودرم جنینی مشتق شده است، این ناهنجاریهای رشدی پوششهای سیستم عصبی (پوست و استخوان) را نیز درگیر میکند. در آنسفالی، مغز و جمجمه کوچک هستند و نوزاد معمولا زنده نمیماند. در اسپینا بیفیدا، نخاع تحتانی و ریشههای عصبی توسعه نیافته اند و ممکن است بدون پوشش پوست یا ستون فقرات استخوانی در پشت نوزاد قرار بگیرند (مننگومیلوسل). چنین نوزادانی با اندام تحتانی پژمرده، فلج و بیهوش همراه با بی اختیاری روده و مثانه باقی میمانند.
مروری بر آناتومی سیستم عصبی مرکزی
پوششها و خون رسانی
مغز و نخاع به ترتیب توسط استخوانهای جمجمه و ستون مهرهها حمایت و محافظت میشوند. در داخل این پوششهای استخوانی، CNS به طور کامل توسط سه لایه غشاء به نام مننژ پوشانده شده است (شکل ۱.۱۳). بیرونی ترین غشای سخت شامه، پوششی محکم و فیبری است که مغز و نخاع را مانند یک کیسه گشاد احاطه کرده است. دورای نخاعی و قسمت اعظم سختی جمجمه ای جدا از پریوستئوم است که استخوانهای اطراف را میپوشاند. با این حال، در مکانهای خاصی، مانند کف حفره جمجمه، سختشکم و پریوستوم به هم میپیوندند و سختی دور جمجمه به داخل جمجمه میچسبد. دو ورقه بزرگ دورا وارد حفره جمجمه میشوند و به طور ناقص آن را به بخشهایی تقسیم میکنند.شکل ۱.۱۴). مغز فاکس در صفحه ساژیتال بین دو نیمکره مغز قرار دارد. مرز آزاد آن در بالای جسم پینه ای قرار دارد. مخچه تنتوریوم به صورت افقی قرار دارد و در زیر لوبهای پس سری نیمکرههای مغزی و بالای مخچه قرار دارد. سخت شامه را میتوان متشکل از دو لایه در نظر گرفت. اینها به جز در مکانهای خاصی با هم ترکیب میشوند، جایی که از هم جدا میشوند و فضاهایی را تشکیل میدهند، سینوسهای وریدی دورال، که به عنوان کانالهایی برای تخلیه وریدی مغز عمل میکنند. سینوسهای دورال مهم رخ میدهد:
![]() در کف حفره جمجمه
در کف حفره جمجمه
![]() در امتداد خطوط اتصال فالک سربری و تنتوریوم مخچه به داخل جمجمه (سینوس ساژیتال فوقانی، شکل ۱.۱۴؛ سینوس عرضی، شکل ۷.۹، ۷.۱۰)
در امتداد خطوط اتصال فالک سربری و تنتوریوم مخچه به داخل جمجمه (سینوس ساژیتال فوقانی، شکل ۱.۱۴؛ سینوس عرضی، شکل ۷.۹، ۷.۱۰)
![]() در امتداد خط اتصال فالک سربری و تنتوریوم مخچه به یکدیگر (سینوس مستقیم) Figs 7.9, ۷.۱۰).
در امتداد خط اتصال فالک سربری و تنتوریوم مخچه به یکدیگر (سینوس مستقیم) Figs 7.9, ۷.۱۰).
 شکل ۱.۱۳ بخشی از جمجمه که روابط بین مننژها و CNS را نشان میدهد.
شکل ۱.۱۳ بخشی از جمجمه که روابط بین مننژها و CNS را نشان میدهد.
 شکل ۱.۱۴ بخش پاراساژیتال سر که وضعیت فالکس سربری و تنتوریوم مخچه را نشان میدهد.
شکل ۱.۱۴ بخش پاراساژیتال سر که وضعیت فالکس سربری و تنتوریوم مخچه را نشان میدهد.
پوششها و خون رسانی سیستم عصبی مرکزی
![]() مغز و نخاع توسط سه لایه مننژی پوشانده میشوند: سخت شامه، عنکبوتیه ماتر و پیا ماتر.
مغز و نخاع توسط سه لایه مننژی پوشانده میشوند: سخت شامه، عنکبوتیه ماتر و پیا ماتر.
![]() دو ورقه سخت شامه جمجمه به نامهای falx cerebri و tentorium cerebelli به طور ناقص حفره جمجمه را به بخشهایی تقسیم میکنند.
دو ورقه سخت شامه جمجمه به نامهای falx cerebri و tentorium cerebelli به طور ناقص حفره جمجمه را به بخشهایی تقسیم میکنند.
![]() سخته جمجمه ای حاوی سینوسهای وریدی دورال است که به عنوان کانالی برای تخلیه وریدی مغز عمل میکنند.
سخته جمجمه ای حاوی سینوسهای وریدی دورال است که به عنوان کانالی برای تخلیه وریدی مغز عمل میکنند.
![]() در زیر ماده عنکبوتیه فضای زیر عنکبوتیه قرار دارد که مایع مغزی نخاعی (CSF) در آن گردش میکند.
در زیر ماده عنکبوتیه فضای زیر عنکبوتیه قرار دارد که مایع مغزی نخاعی (CSF) در آن گردش میکند.
![]() مغز از طریق شریانهای کاروتید داخلی و مهره ای خون تامین میشود.
مغز از طریق شریانهای کاروتید داخلی و مهره ای خون تامین میشود.
![]() طناب نخاعی توسط عروق ناشی از شریانهای مهره ای تامین میشود که توسط شریانهای رادیکولار مشتق شده از عروق سگمنتال تقویت میشود.
طناب نخاعی توسط عروق ناشی از شریانهای مهره ای تامین میشود که توسط شریانهای رادیکولار مشتق شده از عروق سگمنتال تقویت میشود.
در زیر دورا ماده عنکبوتیه قرار دارد که این دو توسط یک فضای نازک ساب دورال از هم جدا شده اند. عنکبوتیه یک غشای کلاژنی شفاف است که مانند سخت شامه مغز و نخاع را به طور شل میپوشاند. داخلی ترین مننژها پیا ماتر است، غشایی ظریف با ضخامت میکروسکوپی که محکم به سطح مغز و نخاع میچسبد و خطوط آنها را به دقت دنبال میکند. بین عنکبوتیه و پیا فضای زیر عنکبوتیه قرار دارد که CSF از طریق آن گردش میکند.
خون شریانی مغز توسط شریانهای کاروتید داخلی و مهرهای تامین میشود، که آناستوموز میکنند و سیرکولوس شریانی (دایره ویلیس) را در پایه مغز تشکیل میدهند. طناب نخاعی توسط عروق ناشی از شریانهای مهره ای تامین میشود که توسط شریانهای رادیکولار مشتق شده از عروق سگمنتال تقویت میشود. شریانها و وریدهایی که به CNS خدمت میکنند بخشی از مسیر خود را در فضای زیر عنکبوتیه میگذرانند (شکل ۱.۱۳). مننژها توسط تعدادی رگ تامین میشوند که مهمترین عروق داخل جمجمهای، شریان مننژیال میانی است که بهطور گسترده بین جمجمه و سختشکم در قسمت جانبی نیمکره مغز منشعب میشود.
آناتومی نخاع
طناب نخاعی در کانال مهره ای (نخاعی) ستون مهره قرار دارد و به صورت منقاری با بصل النخاع ساقه مغز پیوسته است (شکل ۱.۱۵). نخاع اطلاعات را از تنه و اندام دریافت میکند و آن را کنترل میکند. این امر از طریق ۳۱ جفت اعصاب نخاعی حاصل میشود که در فواصل زمانی در طول آن به طناب میپیوندند و حاوی رشتههای عصبی آوران و وابران هستند که با ساختارهای پیرامونی متصل میشوند. در نزدیکی طناب، اعصاب نخاعی به ریشههای پشتی و شکمیتقسیم میشوند که به ترتیب در امتداد مرزهای پشتی و بطنی به طناب متصل میشوند (شکل ۱.۱۶). ریشههای پشتی دارای الیاف آوران هستند که بدنه سلولی آن در گانگلیونهای ریشه پشتی قرار دارد. ریشههای شکمیحامل الیاف وابران با اجسام سلولی در داخل ماده خاکستری ستون فقرات هستند. اعصاب نخاعی از طریق سوراخهای کوچکی به نام سوراخ بین مهره ای که بین مهرههای مجاور قرار دارد، کانال مهره را ترک میکنند . به دلیل تفاوت در سرعت رشد طناب نخاعی و ستون فقرات در طول رشد، نخاع در بزرگسالان تمام طول کانال مهره را گسترش نمیدهد، اما به سطح دیسک بین مهرهای بین L1 و L2 ختم میشود. بنابراین، اعصاب نخاعی کمری و ساکرال به ترتیبی بند مانند، دم اسب (شکل ۱.۱۵)، پایین میآیند تا به سوراخ خروجی خود برسند.
 شکل ۱.۱۵ جنبه پشتی نخاع در محل.
شکل ۱.۱۵ جنبه پشتی نخاع در محل.
 شکل ۱.۱۶ نمودار شماتیک یک مقطع عرضی از طناب نخاعی، اتصال ریشههای عصبی نخاعی و آرایش ماده خاکستری و سفید را نشان میدهد.
شکل ۱.۱۶ نمودار شماتیک یک مقطع عرضی از طناب نخاعی، اتصال ریشههای عصبی نخاعی و آرایش ماده خاکستری و سفید را نشان میدهد.
نخاع ساختاری نسبتاً تمایز نیافته در مقایسه با مغز است. در نتیجه، اصول اساسی سازمان، که در اوایل رشد جنینی ایجاد شده است، میتواند به آسانی حتی در بند ناف انسان بالغ شناسایی شود (شکل ۱.۱۶). طناب نخاعی تقریباً استوانه ای شکل است که در مرکز آن یک کانال مرکزی وستیژیال قرار دارد. جداسازی اجسام سلولی از رشتههای عصبی، شکل مشخصه «H» یا «پروانه» را به هسته مرکزی ماده خاکستری که کانال مرکزی را احاطه کرده است میدهد. چهار امتداد ماده خاکستری مرکزی به صورت پشتی و بطنی به سمت خطوط اتصال ریشههای پشتی و شکمیاعصاب نخاعی پیش میروند. اینها به نام شاخ پشتی و شاخ شکمیشناخته میشوند، به ترتیب. شاخ پشتی محل خاتمه بسیاری از نورونهای آوران است که تکانههای گیرندههای حسی را در سراسر بدن منتقل میکند و محل منشأ مسیرهای صعودی است که تکانههای حسی را به مغز منتقل میکنند. شاخ شکمیحاوی نورونهای حرکتی است که عضله اسکلتی را عصب دهی میکند. علاوه بر این، تنها در سطوح قفسه سینه و کمری فوقانی بند ناف، مجموعه دیگری از اجسام سلولی کوچکتر شامل شاخ جانبی است که حاوی نورونهای پیش گانگلیونی متعلق به بخش سمپاتیک سیستم عصبی خودمختار است.
آناتومی نخاع
![]() طناب نخاعی در داخل کانال مهره قرار دارد. دارای ۳۱ جفت اعصاب نخاعی است که از طریق آنها الیاف را از اطراف دریافت میکند و الیاف را به اطراف میفرستد.
طناب نخاعی در داخل کانال مهره قرار دارد. دارای ۳۱ جفت اعصاب نخاعی است که از طریق آنها الیاف را از اطراف دریافت میکند و الیاف را به اطراف میفرستد.
![]() در نزدیکی طناب، اعصاب نخاعی تقسیم میشوند و ریشههای پشتی و شکمیرا تشکیل میدهند. ریشههای پشتی دارای الیاف آوران با بدن سلولی در گانگلیونهای ریشه پشتی هستند و ریشههای شکمیحامل الیاف وابران هستند.
در نزدیکی طناب، اعصاب نخاعی تقسیم میشوند و ریشههای پشتی و شکمیرا تشکیل میدهند. ریشههای پشتی دارای الیاف آوران با بدن سلولی در گانگلیونهای ریشه پشتی هستند و ریشههای شکمیحامل الیاف وابران هستند.
![]() نخاع از یک هسته مرکزی ماده خاکستری، حاوی اجسام سلولهای عصبی و یک لایه بیرونی ماده سفید یا رشتههای عصبی تشکیل شده است.
نخاع از یک هسته مرکزی ماده خاکستری، حاوی اجسام سلولهای عصبی و یک لایه بیرونی ماده سفید یا رشتههای عصبی تشکیل شده است.
![]() در داخل ماده خاکستری، شاخ پشتی حاوی نورونهای حسی، شاخ شکمیحاوی نورونهای حرکتی و شاخ جانبی حاوی نورونهای سمپاتیک پیش گانگلیونی است.
در داخل ماده خاکستری، شاخ پشتی حاوی نورونهای حسی، شاخ شکمیحاوی نورونهای حرکتی و شاخ جانبی حاوی نورونهای سمپاتیک پیش گانگلیونی است.
![]() در داخل ماده سفید مسیرهای فیبر عصبی صعودی و نزولی وجود دارد که نخاع را با مغز پیوند میدهد.
در داخل ماده سفید مسیرهای فیبر عصبی صعودی و نزولی وجود دارد که نخاع را با مغز پیوند میدهد.
![]() مسیرهای صعودی اصلی ستونهای پشتی، مسیرهای اسپینوتالاموس و مجاری نخاعی مخچه هستند. دستگاه قشر نخاعی یک مسیر نزولی مهم است.
مسیرهای صعودی اصلی ستونهای پشتی، مسیرهای اسپینوتالاموس و مجاری نخاعی مخچه هستند. دستگاه قشر نخاعی یک مسیر نزولی مهم است.
محیط بند ناف از ماده سفید تشکیل شده است که شامل رشتههای عصبی درازا میباشد. اینها به یک سری از مسیرهای صعودی سازماندهی میشوند که اطلاعات را از تنه و اندامها به مغز میرسانند، و مسیرهای نزولی که توسط آنها مغز فعالیتهای نورونهای نخاع را کنترل میکند (شکل ۱.۱۷). مسیرهای صعودی اصلی ستونهای پشتی (fasciculus gracilis و fasciculus cuneatus) هستند که لمس ظریف و حس عمقی را حمل میکنند، مجاری اسپینوتالاموس که حامل درد، دما، لمس و فشار درشت هستند، و مجاری نخاعی مخچه ای هستند.که اطلاعات را از گیرندههای عضلانی و مفصلی به مخچه منتقل میکند. در میان مسیرهای نزولی، یکی از مهمترین آنها، دستگاه قشر نخاعی جانبی است که حرکات ارادی ماهرانه را کنترل میکند.
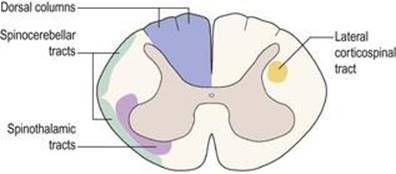 شکل ۱.۱۷ مقطع عرضی از طناب نخاعی که محل مسیرهای فیبر عصبی اصلی صعودی (سمت چپ) و نزولی (سمت راست) را نشان میدهد.
شکل ۱.۱۷ مقطع عرضی از طناب نخاعی که محل مسیرهای فیبر عصبی اصلی صعودی (سمت چپ) و نزولی (سمت راست) را نشان میدهد.
آناتومی مغز
ویژگیها و نشانههای اصلی
مغز تحت سلطه نیمکرههای مغزی است (شکلهای ۱.۱۱، ۱.۱۲). اینها یک گوشته بیرونی بسیار پیچیده از ماده خاکستری و یک هسته داخلی از ماده سفید دارند. برخی از پیچشهای سطحی عملکردهای حسی یا حرکتی خاصی دارند که در زیر توضیح داده شده است. دو نیمکره مغزی به طور ناقص توسط یک شکاف طولی بزرگ از هم جدا شده اند. شکاف به طور معمول توسط فلکس سربری اشغال میشود و در اعماق آن جسم پینه ای قرار دارد که حاوی الیاف همسو است که مناطق متناظر دو نیمکره را متحد میکند.
هنگامیکه مغز به صورت شکمیمشاهده میشود، ساقه مغز به وضوح دیده میشود، اگرچه روابط مغز میانی به بهترین وجه در بخش ساژیتال نشان داده میشود. ساقه مغز منشا اعصاب جمجمه III-XII است. پشتی (خلفی) به ساقه مغز مخچه قرار دارد. مخچه تنتوریوم معمولاً بین مخچه و قسمت خلفی (لوبهای پس سری) نیمکرههای مغز قرار دارد.
سیستم بطنی
طرح بسیار ساده شده مغز پایه، که در بالا توضیح داده شد، طرح مفیدی است که در آن میتوان وضعیت کلی سیستم بطنی را در نظر گرفت (شکلهای ۱.۱۰، ۱.۱۲، ۱.۱۸). همانطور که کانال مرکزی نخاع به سمت ساقه مغز بالا میرود، به تدریج در جهت پشتی حرکت میکند و در نهایت باز میشود تا یک فرورفتگی کم عمق و لوزی شکل در سطح پشتی مدولا و پونز (قسمت مغزی عقبی ساقه مغز) ایجاد شود.) زیر مخچه. این بطن چهارم است.
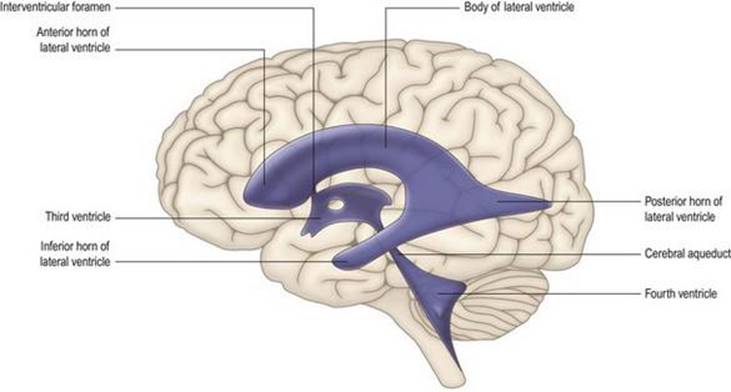 شکل ۱.۱۸ سیستم بطنی مغزی.
شکل ۱.۱۸ سیستم بطنی مغزی.
در مرز منقاری حوضچهها، دیوارههای بطن چهارم به هم میرسند و بار دیگر یک لوله باریک به نام قنات مغزی را تشکیل میدهند. قنات مغزی به درون ماده ای از ساقه مغز که در طول مغز میانی در زیر کولیکولهای تحتانی و فوقانی قرار دارد فرو میرود. در محل اتصال مغز میانی و جلوی مغز، قنات به بطن سوم باز میشود، محفظهای شکافمانند، باریک از پهلو به پهلو، اما در ابعاد پشتی و شکمیو روسترو دمیگسترده است. دیوارههای جانبی بطن سوم توسط تالاموس و هیپوتالاموس دیانسفالون تشکیل شده است. در نزدیکی انتهای منقاری بطن سوم، یک روزنه کوچک، سوراخ بین بطنی (فورامن مونرو)، با یک محفظه وسیع، بطن جانبی ارتباط برقرار میکند.، در هر نیمکره مغزی. سیستم بطنی حاوی CSF است که توسط شبکه کوروئید ترشح میشود.
ساقه مغز
هنگامیکه مغز به صورت خارجی مشاهده میشود، نیمکرههای بزرگ مغز بسیاری از ساختارهای دیگر را مبهم میکند، اما یک بخش ساژیتال وسط (شکل ۱.11B، ۱.12A) بیشتر ویژگیهای اصلی مغز اصلی را نشان میدهد. ساقه مغز را میتوان به وضوح در هر دو بخش ساژیتال وسط و نمای شکمی(شکل ۱.۱۱، ۱.۱۲) مغز مشاهده کرد. ساقه مغز از بصل النخاع، پونز و مغز میانی تشکیل شده است.
ساقه مغز تنها بخش کوچکی از کل مغز را تشکیل میدهد، اما بسیار مهم است. از طریق آن، مسیرهای فیبر عصبی صعودی و نزولی که مغز و نخاع را به هم پیوند میدهند، میگذرد (شکل ۱.۱۹). اینها اطلاعات حسی را از تنه و اندامها حمل میکنند و به آنها اجازه حرکت میدهند. ساقه مغز همچنین شامل محلهای منشأ و پایان بسیاری از اعصاب جمجمه ای است که مغز از طریق آنها سر را عصب میکند. علاوه بر این، در داخل ساقه مغز مراکزی قرار دارند که عملکردهای حیاتی مانند تنفس، سیستم قلبی عروقی و سطح هوشیاری را کنترل میکنند.
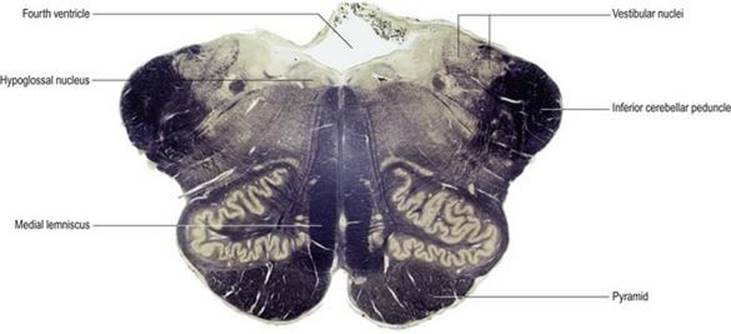 شکل ۱.۱۹ مقطع عرضی از طریق ساقه مغز در سطح بصل النخاع برش به روش وایگرت پال رنگ آمیزی شده است. نواحی غنی از فیبرهای عصبی تیره رنگ میشوند در حالی که نواحی غنی از بدن سلولی نسبتاً کم رنگ تر هستند. این هرم شامل فیبرهای حرکتی نزولی است که از قشر مغز به نخاع میروند. لمنیسکوس داخلی شامل آکسونهای صعودی است که اطلاعات حسی را از اندامها به مراکز بالاتر در مغز حمل میکند. پایه مخچه تحتانی حاوی الیاف نخاعی مخچه ای است که اطلاعات را از مفاصل و ماهیچهها به مخچه منتقل میکند. هستههای دهلیزی محل خاتمه عصب دهلیزی هستند. هسته هیپوگلوسال محل منشاء رشتههای عصبی هیپوگلاس است.
شکل ۱.۱۹ مقطع عرضی از طریق ساقه مغز در سطح بصل النخاع برش به روش وایگرت پال رنگ آمیزی شده است. نواحی غنی از فیبرهای عصبی تیره رنگ میشوند در حالی که نواحی غنی از بدن سلولی نسبتاً کم رنگ تر هستند. این هرم شامل فیبرهای حرکتی نزولی است که از قشر مغز به نخاع میروند. لمنیسکوس داخلی شامل آکسونهای صعودی است که اطلاعات حسی را از اندامها به مراکز بالاتر در مغز حمل میکند. پایه مخچه تحتانی حاوی الیاف نخاعی مخچه ای است که اطلاعات را از مفاصل و ماهیچهها به مخچه منتقل میکند. هستههای دهلیزی محل خاتمه عصب دهلیزی هستند. هسته هیپوگلوسال محل منشاء رشتههای عصبی هیپوگلاس است.
بصل النخاع به صورت دمیبا طناب نخاعی پیوسته است و به صورت منقاری تا حوضچه امتداد دارد. اتصال دوم را میتوان به وضوح در نماهای شکمییا ساژیتال مشاهده کرد زیرا قسمت شکمیپونز یک برآمدگی برجسته در سطح ساقه مغز تشکیل میدهد. در بخش ساژیتال (شکل ۱.11B، ۱.12A)، لومن بطن چهارم بین پونز و مدولا به صورت شکمیو مخچه در پشت ظاهر میشود، که سقف چادری شکل آن به داخل آن کشیده میشود.
اعصاب جمجمه ای
مغز به طور مستقیم اطلاعات حسی را از ساختارهای محیطی، به ویژه سر و گردن، دریافت میکند و آن را کنترل میکند. رشتههای عصبی آوران و وابران در ۱۲ جفت اعصاب جمجمه ای قرار دارند که با نامهای فردی و اعداد رومیI-XII مشخص میشوند. برخی از اعصاب جمجمه ای فقط دارای رشتههای عصبی حسی یا حرکتی هستند، اما اکثر آنها مانند اعصاب نخاعی حاوی مخلوطی هستند. دو عصب اول جمجمه ای (I Olfactory, II optic) مستقیماً به مغز جلویی و بقیه به ساقه مغز متصل میشوند. در درون ساقه مغز تعدادی از گروههای بدن سلولی وجود دارد که هستههای عصبی جمجمه نامیده میشوند. اینها محل خاتمه رشتههای حسی و منشاء رشتههای حرکتی (شکل ۱.۱۹) هستند که در اعصاب جمجمه ای قرار دارند.
مخچه
مخچه توسط توده بزرگی از رشتههای عصبی که در طرفین بطن چهارم قرار دارند به ساقه مغز متصل میشود. توده به طور اسمی به سه قسمت تقسیم میشود: پایه مخچه تحتانی، میانی و فوقانی.
اینها فیبرهای عصبی را به ترتیب بین مدولا، پونز و مغز میانی و مخچه حمل میکنند.
مخچه از یک لایه بیرونی ماده خاکستری به نام قشر مخچه تشکیل شده است که هسته مرکزی ماده سفید را احاطه کرده است. سطح قشر مغز بسیار پیچیده است تا یک الگوی منظم از چینهای باریک، موازی یا برگها را تشکیل دهد. ماده سفید مخچه متشکل از رشتههای عصبی است که به سمت قشر مخچه و از آن خارج میشوند. ماده سفید دارای چینش درخت مانندی در بخش (شکل ۱.11B) است، زیرا شاخههای آن به سمت سطح میرسد. مخچه مربوط به هماهنگی حرکت است و در سطحی کاملاً ناخودآگاه عمل میکند.
مغز میانی نسبتاً کوچکی در منقاری بر روی پونز قرار دارد. در سطح پشتی آن میتوان برجستگیهای گرد کولیکولهای فوقانی و تحتانی را مشاهده کرد که در زیر آنها قنات مغزی قرار دارد (شکلهای ۱.۱۰ – ۱.۱۲).
دیانسفالون و نیمکرههای مغزی
منقاری به ساقه مغز، جلوی مغز قرار دارد که از دیانسفالون و نیمکرههای مغزی تشکیل شده است. دیانسفالون و نیمکره مغز در هر طرف مغز تا حد زیادی از نظر فیزیکی از همتایان خود در طرف دیگر جدا هستند، اگرچه اتصالات متقابل مهمیوجود دارد، همانطور که در زیر توضیح داده شده است. دو طرف دیانسفالون توسط لومن بطن سوم که دیوارههای جانبی آن را تشکیل میدهند از هم جدا میشوند.
دیانسفالون از چهار بخش اصلی در جهت پشتی شکمیتشکیل شده است: اپی تالاموس، تالاموس، ساب تالاموس و هیپوتالاموس. اپیتالاموس کوچک است و قابل توجه ترین جزء آن غده صنوبری است که در خط وسط قرار دارد و بلافاصله به سمت کولیکولهای فوقانی مغز میانی قرار دارد (شکل ۱.12A). تالاموس بزرگترین قسمت دیانسفالون است و بیشتر دیواره جانبی بطن سوم را تشکیل میدهد. تالاموس نقش مهمیدر عملکردهای حسی، حرکتی و شناختی دارد و ارتباطات متقابل گسترده ای با قشر مغز دارد. ساب تالاموس ناحیه کوچکی است که در عمق دیواره بطن قرار دارد. حاوی هسته ساب تالاموس استکه از نظر عملکردی نزدیک به عقدههای قاعده ای مرتبط است (فصل ۱۴). هیپوتالاموس قسمت پایینی دیوارهها و کف بطن سوم را تشکیل میدهد. این یک منطقه پیچیده و بسیار مهم است زیرا در سیستم عصبی خودمختار (فصل ۴)، سیستم لیمبیک و سیستم عصبی غدد درون ریز (فصل ۱۶) دخالت دارد. از قسمت شکمیهیپوتالاموس در خط میانی، ساقه زیرین یا هیپوفیز بیرون میآید که غده هیپوفیز به آن متصل است (شکل ۱.۱۲).
نیمکره مغز تا حد زیادی بزرگترین بخش مغز است. مانند مخچه، از یک لایه بیرونی یا قشر از ماده خاکستری و یک توده داخلی از ماده سفید تشکیل شده است (شکل ۱.۲۰، ۱.۲۱). علاوه بر این، تا حدی در داخل ماده سفید زیر قشری چندین توده بزرگ از اجسام سلولی قرار دارند که در مجموع به آنها عقدههای پایه میگویند (شکلهای ۱.۵، ۱.۲۰، ۱.۲۱). دو نیمکره مغزی توسط یک شکاف خط میانی عمیق، شکاف طولی بزرگ (شکل ۱.۲۰) از هم جدا میشوند که فالکس سربری (falx cerebri) را در خود جای میدهد. در اعماق شکاف جسم پینه ای قرار دارد (شکل ۱.12A، ۱.۲۰)، ورقه بزرگی از رشتههای عصبی متقاطع (فیبرهای کمسیورال) که نواحی متناظر دو قشر مغز را به هم متصل میکند.
 شکل ۱.۲۰ برش تاج از نیمکره مغزی
شکل ۱.۲۰ برش تاج از نیمکره مغزی
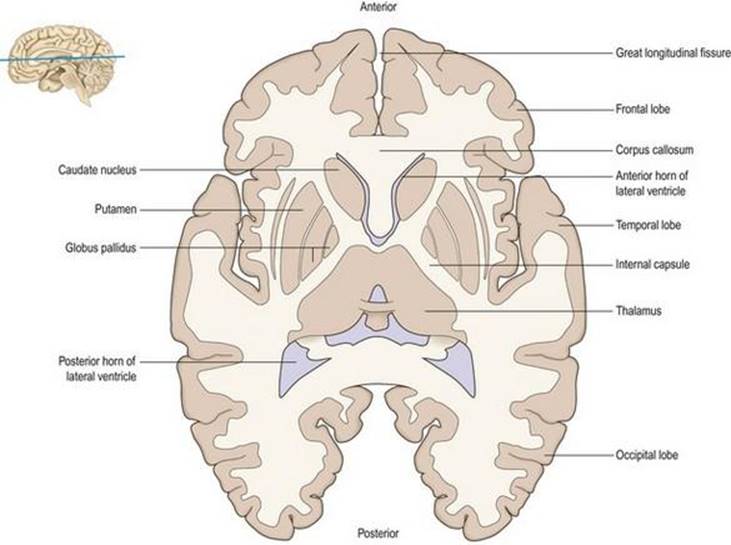 شکل ۱.۲۱ برش افقی از نیمکره مغزی.
شکل ۱.۲۱ برش افقی از نیمکره مغزی.
قشر مغز بسیار پیچیده است. این تأثیر به حداکثر رساندن سطح قشر مغز دارد که حدود ۱ متر مربع برای هر نیمکره است. به پیچیدگیها gyri (مفرد: gyrus) و شیارهای بین آنها sulci (مفرد: sulcus) میگویند. برخی از شکافها و شکافها پیکربندی نسبتاً ثابتی بین افراد دارند و مکان مناطق عملکردی مهم را مشخص میکنند.
در سطح جانبی نیمکره، یک شکاف عمیق، شکاف جانبی (شکل ۱.۲۰، ۱.۲۲)، یک نقطه عطف مهم است. این، همراه با شیارهای خاص، مرزهایی را تشکیل میدهد که نیمکره را به چهار لوب تقسیم میکند (شکل ۱.۲۲). لوبها نام استخوانهای جمجمه را دارند که زیر آن قرار دارند.

 شکل ۱.۲۲ شکنج اصلی، شیارها و نواحی عملکردی قشر مغز. الف) جنبه جانبی؛ (ب) جنبه داخلی.
شکل ۱.۲۲ شکنج اصلی، شیارها و نواحی عملکردی قشر مغز. الف) جنبه جانبی؛ (ب) جنبه داخلی.
قدامیترین قسمت نیمکره مغز را لوب فرونتال مینامند که بیشترین تحدب قدامیآن قطب پیشانی است. مرز خلفی لوب فرونتال شیار مرکزی است که میتواند به عنوان یک شیار منفرد و پیوسته که بر روی کل سطح جانبی نیمکره از شکاف طولی بزرگ تا شکاف جانبی قرار دارد شناسایی شود. در خلف شیار مرکزی، لوب جداری قرار دارد که توسط شکاف جانبی از لوب تمپورال زیر جدا میشود. نوک لوب تمپورال قطب تمپورال نامیده میشود. قسمت خلفی نیمکره لوب پس سری است که به قطب اکسیپیتال ختم میشود.مرزهای بین لوب آهیانه و تمپورال و لوب پس سری در سطح جانبی نیمکره نامشخص است، زیرا با هیچ شکاف خاصی مطابقت ندارند. با این حال، در سطح داخلی، لوب جداری و پس سری توسط یک شیار جداری-اکسیپیتال عمیق جدا شده است. در قسمت داخلی نیمکره مغزی، شیار سینگوله موازی با حاشیه بالایی جسم پینه ای قرار دارد. این یک ناحیه از قشر را مشخص میکند که همراه با بخشهایی از قسمت داخلی قشر تمپورال، گاهی اوقات به عنوان لوب لیمبیک شناخته میشود.
وظایف قشر مغز با جزئیات بیشتر در فصل ۱۳ توضیح داده شده است. با این حال، در ابتدا، شناسایی چهار ناحیه عملکردی مهم قشر، یکی در هر لوب مفید خواهد بود (شکل ۱.۲۲).
![]() در لوب فرونتال، شکنجی که در جلوی شیار مرکزی قرار دارد، از نظر تشریحی به عنوان شکنج پیش مرکزی شناخته میشود. از نظر عملکردی، این شامل قشر حرکتی اولیه است که بالاترین سطح در مغز برای کنترل حرکت است. در اینجا، در هر نیمکره، نیمه مخالف بدن به شکلی بسیار دقیق نشان داده شده است.
در لوب فرونتال، شکنجی که در جلوی شیار مرکزی قرار دارد، از نظر تشریحی به عنوان شکنج پیش مرکزی شناخته میشود. از نظر عملکردی، این شامل قشر حرکتی اولیه است که بالاترین سطح در مغز برای کنترل حرکت است. در اینجا، در هر نیمکره، نیمه مخالف بدن به شکلی بسیار دقیق نشان داده شده است.
![]() در لوب جداری، رو به قشر حرکتی اولیه در سراسر شیار مرکزی، شکنج پس مرکزی یا قشر حسی تنی اولیه قرار دارد. این محل خاتمه مسیرهایی است که مدالیتههای لمس، فشار، درد و دما را از نیمه مخالف بدن حمل میکنند و این منطقه ای است که آنها آگاهانه درک میشوند. حواس خاص بالاترین سطح بازنمایی خود را در سایر زمینهها دارند.
در لوب جداری، رو به قشر حرکتی اولیه در سراسر شیار مرکزی، شکنج پس مرکزی یا قشر حسی تنی اولیه قرار دارد. این محل خاتمه مسیرهایی است که مدالیتههای لمس، فشار، درد و دما را از نیمه مخالف بدن حمل میکنند و این منطقه ای است که آنها آگاهانه درک میشوند. حواس خاص بالاترین سطح بازنمایی خود را در سایر زمینهها دارند.
![]() قشر بینایی در لوب اکسیپیتال، بیشتر در قسمت داخلی نیمکره در شکم در بالا و پایین شیار آهکی با جهت افقی قرار دارد.
قشر بینایی در لوب اکسیپیتال، بیشتر در قسمت داخلی نیمکره در شکم در بالا و پایین شیار آهکی با جهت افقی قرار دارد.
![]() در لوب تمپورال قشر شنوایی قرار دارد. در شکنج گیجگاهی فوقانی که در زیر شکاف جانبی و به موازات آن قرار دارد، موضعی است .
در لوب تمپورال قشر شنوایی قرار دارد. در شکنج گیجگاهی فوقانی که در زیر شکاف جانبی و به موازات آن قرار دارد، موضعی است .
![]() لوب لیمبیک عمدتاً از شکنج سینگوله تشکیل شده است که در قسمت داخلی نیمکره قرار دارد و به موازات جسم پینه ای قرار دارد و تشکیل هیپوکامپ و آمیگدال که در داخل لوب تمپورال قرار دارند. این ساختارهای پیچیده با جنبههای عاطفی رفتار و حافظه سروکار دارند.
لوب لیمبیک عمدتاً از شکنج سینگوله تشکیل شده است که در قسمت داخلی نیمکره قرار دارد و به موازات جسم پینه ای قرار دارد و تشکیل هیپوکامپ و آمیگدال که در داخل لوب تمپورال قرار دارند. این ساختارهای پیچیده با جنبههای عاطفی رفتار و حافظه سروکار دارند.
در طی رشد، نیمکره مغز در نتیجه مهاجرت رو به جلو لوب گیجگاهی، پیکربندی C شکل به خود میگیرد، به طوری که قطب تمپورال در مجاورت لوب فرونتال قرار میگیرد و توسط شکاف جانبی از آن جدا میشود. بنابراین، بطن جانبی در نیمکره نیز اساساً به شکل C است و «شاخهایی» به سمت لوبهای پیشانی، اکسیپیتال و گیجگاهی کشیده شده است (شکل ۱.۱۸).
ساختار اصلی نیمکره مغزی یک گوشته بیرونی از ماده خاکستری، قشر مغز است که در زیر آن توده بزرگ و پیچیده ای از ماده سفید قرار دارد که از رشتههای عصبی به سمت و سوی قشر مغز تشکیل شده است (شکلهای ۱.۲۰، ۱.۲۱، ۱.۲۳ ).
 شکل ۱.۲۳ تشریح سمت راست مغز از جنبه جانبی آن، تشریح تاج و کپسول داخلی را نشان میدهد.
شکل ۱.۲۳ تشریح سمت راست مغز از جنبه جانبی آن، تشریح تاج و کپسول داخلی را نشان میدهد.
الیاف آوران و وابران قشری که بین قشر مغز و ساختارهای زیر قشری مانند جسم مخطط، تالاموس، ساقه مغز و نخاع عبور میکنند در یک الگوی تشعشعی مشخص قرار گرفتهاند، تاج رادیاتا، که به پیچشهای سطح قشر مغز میرسد. شکل ۱.۲۳). الیاف در عمق بیشتری در داخل نیمکره در یک صفحه متراکم از ماده سفید متمرکز میشوند که به عنوان کپسول داخلی شناخته میشود (شکلهای ۱.۲۰، ۱.۲۱، ۱.۲۳).
در اعماق نیمکره، هم داخلی و هم جانبی کپسول داخلی، تودههای اضافی از ماده خاکستری وجود دارد که معمولاً در مجموع به آن عقدههای بازال میگویند. بزرگترین آنها جسم مخطط است که از هسته دمی، پوتامن و گلوبوس پالیدوس تشکیل شده است (شکل ۱.۲۰، ۱.۲۱). هسته دمیدر تمام وسعت خود در دیواره بطن جانبی قرار دارد و مانند بطن C شکل است. عقدههای قاعده ای به کنترل تون عضلانی، وضعیت و حرکت مربوط میشوند (فصل ۱۴).
سازماندهی اولیه مغز
![]() مغز به طور معمول به مغز عقبی، مغز میانی و پیش مغزی تقسیم میشود.
مغز به طور معمول به مغز عقبی، مغز میانی و پیش مغزی تقسیم میشود.
![]() مغز عقبی بیشتر به بصل النخاع، پونز و مخچه تقسیم میشود.
مغز عقبی بیشتر به بصل النخاع، پونز و مخچه تقسیم میشود.
![]() مدولا، پونز و مغز میانی ساقه مغز را تشکیل میدهند.
مدولا، پونز و مغز میانی ساقه مغز را تشکیل میدهند.
![]() پیش مغز از دیانسفالون (تالاموس و هیپوتالاموس) و نیمکره مغز تشکیل شده است.
پیش مغز از دیانسفالون (تالاموس و هیپوتالاموس) و نیمکره مغز تشکیل شده است.
![]() در داخل نیمکره مغز چندین هسته بزرگ به نام عقدههای پایه یا جسم مخطط وجود دارد.
در داخل نیمکره مغز چندین هسته بزرگ به نام عقدههای پایه یا جسم مخطط وجود دارد.
![]() مغز حاوی سیستمیاز حفرهها یا بطنهای حاوی CSF است که توسط شبکه کوروئید تولید میشود.
مغز حاوی سیستمیاز حفرهها یا بطنهای حاوی CSF است که توسط شبکه کوروئید تولید میشود.
![]() مغز دارای ۱۲ جفت عصب جمجمه ای است که الیاف آوران و وابران را حمل میکنند.
مغز دارای ۱۲ جفت عصب جمجمه ای است که الیاف آوران و وابران را حمل میکنند.
![]() دو نیمکره مغزی توسط فیبرهای جسم پینه ای به هم متصل شده اند.
دو نیمکره مغزی توسط فیبرهای جسم پینه ای به هم متصل شده اند.
![]() سطح نیمکره مغز از ماده خاکستری قشر مغز تشکیل شده است که به شکل گیره و شکاف چین خورده است. در زیر سطح، تودههای الیاف متراکم تاج رادیاتا و کپسول داخلی قرار دارند. سطح به لوبها تقسیم میشود:
سطح نیمکره مغز از ماده خاکستری قشر مغز تشکیل شده است که به شکل گیره و شکاف چین خورده است. در زیر سطح، تودههای الیاف متراکم تاج رادیاتا و کپسول داخلی قرار دارند. سطح به لوبها تقسیم میشود:
![]() لوب فرونتال حاوی موتور اولیه و قشر پیش حرکتی
لوب فرونتال حاوی موتور اولیه و قشر پیش حرکتی
![]() لوب جداری حاوی قشر حسی جسمیاولیه
لوب جداری حاوی قشر حسی جسمیاولیه
![]() لوب تمپورال حاوی قشر شنوایی اولیه
لوب تمپورال حاوی قشر شنوایی اولیه
![]() لوب اکسیپیتال حاوی قشر بینایی اولیه است
لوب اکسیپیتال حاوی قشر بینایی اولیه است
![]() لوب لیمبیک حاوی مناطقی برای حافظه و جنبههای احساسی رفتار است.
لوب لیمبیک حاوی مناطقی برای حافظه و جنبههای احساسی رفتار است.
مسیرهای حسی اصلی
اطلاعات حسی در مورد محیط داخلی و خارجی در رشتههای عصبی آوران که در اعصاب جمجمه و نخاعی جریان دارند به CNS منتقل میشود. اطلاعات حسی را میتوان تحت عنوان «حواس خاص» و «حواس عمومی» طبقه بندی کرد. حواس خاص همگی در اعصاب جمجمه ای حمل میشوند و از بویایی (عصب جمجمه ای I)، بینایی (II)، چشایی (VII و IX) و شنوایی و عملکرد دهلیزی (VIII) تشکیل شده اند. حواس خاص در جای دیگری با جزئیات بیشتر مورد بررسی قرار میگیرد.
حواس عمومیشامل حالتهای لمس، فشار، درد و دما (که از گیرندههای بیرونی در پوست و گیرندههای درونی در احشاء منتقل میشود)، و آگاهی از وضعیت و حرکت (از گیرندههای عمقی در مفاصل، تاندونها و ماهیچهها) است. اطلاعات حسی عمومی از تنه و اندامها در اعصاب نخاعی حمل میشود. از سر در عصب سه قلو (عصب جمجمه ای V) حمل میشود.
برای تمام روشهای دستهبندی حس عمومی، یک توالی از سه نورون بین گیرنده حسی واقع در حاشیه و درک حس در سطح قشر مغز وجود دارد (شکل ۱.۲۴). اولین نورون (نورون مرتبه اول یا نورون آوران اولیه) از طریق یک عصب نخاعی یا عصب سه قلو در همان سمت بدن که گیرنده محیطی آن قرار دارد وارد نخاع یا ساقه مغز میشود. بدنه سلولی نورون مرتبه اول در گانگلیون ریشه پشتی یک عصب نخاعی یا در گانگلیون سه قلو قرار دارد. در CNS، نورون مرتبه اول در همان سمت باقی میماند (همان طرف) و روی نورون دوم (نورون مرتبه دوم) سیناپس میکند. نورون مرتبه دوم دارای بدنه سلولی خود در نخاع یا ساقه مغز است که محل دقیق آن بسته به روش مورد نظر است. آکسون آن از طرف دیگر CNS عبور میکند و به سمت تالاموس بالا میرود و در آنجا خاتمه مییابد. سومین نورون در این دنباله (نورون مرتبه سوم) بدن سلولی خود را در تالاموس دارد و آکسون آن به قشر حسی تنی که در شکنج پس مرکزی لوب جداری نیمکره مغز قرار دارد، قرار دارد.
 شکل ۱.۲۴ نمای کلی از مسیرهای اصلی برای احساس عمومی.
شکل ۱.۲۴ نمای کلی از مسیرهای اصلی برای احساس عمومی.
به طور خاص، آورانهای نخاعی اولیه که اطلاعات لمس/فشار، درد و دما را از اندامها و تنه حمل میکنند نزدیک به سطح ورودشان به نخاع ختم میشود. آنها با نورونهای مرتبه دوم سیناپس میشوند، آکسونهای آنها در چند بخش جدا میشوند و سپس دستگاه اسپینوتلاموس را تشکیل میدهند. در مقابل، آورانهای نخاعی اولیه حامل اطلاعات حس عمقی و لمس متمایز (ریز) بدون وقفه در همان سمت طناب هنگام ورودشان بالا میروند و ستونهای پشتی (fasciculus gracilis و fasciculus cuneatus) را تشکیل میدهند. آنها به هستههای ستون پشتی (nuclei gracilis و cuneatus) واقع در مدولا ختم میشوند. از اینجا، نورونهای مرتبه دوم بحث میشوند و به عنوان تالاموس به سمت تالاموس صعود میکنند لمنیسکوس داخلی. نورونهای آوران اولیه که در عصب سه قلو وارد ساقه مغز میشوند، به صورت همان طرف در هسته حسی سه قلو، یکی از هستههای عصب جمجمه، خاتمه مییابند. از اینجا، نورونهای مرتبه دوم به عنوان مجرای تریژمینوتالاموس به تالاموس میروند . نورونهای حسی مرتبه دوم، با منشا نخاع یا ساقه مغز، در همان ناحیه تالاموس (هسته خلفی شکمی) همگرا میشوند و روی نورونهای مرتبه سوم سیناپس میشوند که به قشر حسی تنی در شکنج پس مرکزی لوب جداری میرسند.در سراسر برجستگیهای مرکزی سیستم حسی تنی درجه بالایی از تفکیک فضایی نورونهای نشان دهنده قسمتهای مختلف بدن وجود دارد (به اصطلاح سوماتوتوپیک). سازمان). این به طور چشمگیری در سطح قشر مغز نشان داده شده است ( شکل ۱۳.۲۰ را ببینید). در اینجا ناحیه حسی تنی نواری از قشر مغز را اشغال میکند که از جنبه داخلی نیمکره (ناحیه پا) تا قسمت تحتانی لوب جداری (ناحیه سر) امتداد دارد.
مسیرهای اصلی موتور
نورونهای حرکتی که مستقیماً عضله اسکلتی را عصب دهی میکنند دارای بدنه سلولی هستند که در ماده خاکستری نخاع و ساقه مغز قرار دارند و اغلب به عنوان نورونهای حرکتی تحتانی شناخته میشوند. آنها به اصطلاح”مسیر مشترک نهایی”را تشکیل میدهند که توسط آن سیستم عصبی حرکت را کنترل میکند. در مقابل، نورونهایی که فعالیت نورونهای حرکتی تحتانی را کنترل میکنند، در مجموع به عنوان نورونهای حرکتی فوقانی شناخته میشوند. آنها تعدادی مسیر نزولی را تشکیل میدهند که از طریق ساقه مغز و نخاع میگذرد. از جمله مهمترین آنها، دستگاههای قشر نخاعی و کورتیکوبولبار هستند (شکل ۱.۲۵). این مسیرها تا حدی از نورونهای ناحیه حرکتی قشر مغز منشأ میگیرند، جایی که کل بدن به شکل سوماتوتوپیک نشان داده میشود (شکل ۱۳.۲۰). آکسونها از کپسول داخلی عبور کرده و وارد ساقه مغز میشوند، جایی که اکثر آنها به سمت دیگر تجزیه میشوند. این بدان معنی است که حرکات یک طرف بدن توسط قشر مغز مخالف کنترل میشود. فیبرهای کورتیکوبولبار فعالیت نورونهای حرکتی واقع در هستههای عصبی جمجمه را کنترل میکنند که عضلات اسکلتی سر و گردن را از طریق اعصاب جمجمه عصب دهی میکنند. فیبرهای قشر نخاعی فعالیت نورونهای حرکتی در نخاع را کنترل میکنند که عضلات تنه و اندام را عصب دهی میکنند. محل عبور فیبرهای قشر نخاعی به طرف دیگر سیستم عصبی را میتوان در قسمت شکمیمدولا مشاهده کرد. شکل ۹.۴) و به عنوان بحث از اهرام شناخته میشود. به همین دلیل، دستگاه قشر نخاعی به عنوان دستگاه هرمینیز شناخته میشود.
 شکل ۱.۲۵ مروری بر مسیرهای اصلی موتور.
شکل ۱.۲۵ مروری بر مسیرهای اصلی موتور.
عملکرد اصلی مسیرهای کورتیکوبولبار و قشر نخاعی کنترل حرکات ارادی و ماهرانه است. بنابراین، بخش بزرگی از قشر حرکتی و مسیرهای نزولی آن به آن قسمتهایی از بدن اختصاص دارد که قادر به انجام حرکات ظریف و به اصطلاح «قطعشده» هستند، مانند ماهیچههای گفتار و بیان صورت و ماهیچههایی که دست را کنترل میکنند.
ساختارهای مغزی متعددی جدا از سیستم قشر نخاعی یا هرمیدر کنترل حرکت، وضعیت بدن و تون عضلات نقش دارند. اینها گاهی اوقات مجموعاً به عنوان مسیرهای خارج هرمیشناخته میشوند. آنها شامل هستههای خاصی در ساقه مغز، مانند هستههای دهلیزی و هستههای مشبک (تشکیل مشبک)، و همچنین عقدههای قاعدهای و هستههای زیر قشری مربوطه واقع در جلو مغز هستند. هستههای دهلیزی و شبکهای بر نورونهای حرکتی نخاعی از طریق اتصالات نزولی در مجاری دهلیزی و نخاعی تأثیر میگذارند.آنها در کنترل تون عضلات و وضعیت بدن مهم هستند. عقدههای قاعده ای اعمال خود را بر روی نورونهای حرکتی تحتانی ساقه مغز و طناب نخاعی طرف مقابل از طریق مسیرهای پیچیده و غیر مستقیم اعمال میکنند (شکل ۱.۲۶). اینها شامل برآمدگی از طریق تالاموس به نواحی حرکتی قشر مغز و برآمدگی به تشکیل مشبک ساقه مغز است. عقدههای پایه در تسهیل رفتار حرکتی مناسب و مهار حرکات ناخواسته مهم هستند (فصل ۱۴).
 شکل ۱.۲۶ نمای کلی اتصالات عقدههای قاعده ای.
شکل ۱.۲۶ نمای کلی اتصالات عقدههای قاعده ای.
مخچه مرکز مهمیاست که در آن برنامههای حرکتی تولید شده در ناحیه حرکتی قشر مخ با بازخورد حسی در مورد سرعت و جهت حرکات فعال اندامها، سر و گردن در فضا مقایسه میشود. این برای حرکت دقیق، هماهنگ و هدفمند ضروری است. مخچه اتصالات آوران را از طناب نخاعی از طریق مجاری نخاعی مخچه و از سیستم دهلیزی و قشر حرکتی دریافت میکند. اتصالات وابران آن پیچیده است، اما عمدتاً به شکل بازخورد به تالاموس و سپس به قشر حرکتی است (شکل ۱.۲۷) آورانها به هر طرف مخچه از نیمه همان طرف نخاع و ساقه مغز و از قشر مغز طرف مقابل میآیند. برجستگیهای وابران به تالاموس طرف مقابل و قشر مغز از طریق بحث در مغز میانی هدایت میشوند. به همین دلیل و بحث در مورد مسیرهای حرکتی نزولی قشر مغز، هر طرف مخچه حرکات سمت همان طرف بدن را هماهنگ میکند (فصل ۱۱).
 شکل ۱.۲۷ نمای کلی اتصالات مخچه.
شکل ۱.۲۷ نمای کلی اتصالات مخچه.
اصول اولیه تشخیص بالینی
دانش نوروآناتومی یک پیش نیاز برای تشخیص بالینی اختلالات سیستم عصبی است. فرآیند دستیابی به تشخیص با گرفتن شرح حال، سپس معاینه عصبی و در نهایت با بررسیهای تاییدی پیش میرود (شکل ۱.۲۸). گرفتن شرح حال سرنخهایی برای علت یا علت بیماری ارائه میدهد، در حالی که معاینه بالینی محل ضایعه را مشخص میکند (شکل ۱.۲۹). یک ضایعه پاتولوژیک که در یک محل خاص در محور عصبی عضلانی عمل میکند یک سندرم قابل تشخیص را تشکیل میدهد که بررسی آن منجر به ایجاد اتیولوژی یا تشخیص میشود.

شکل ۱.۲۸ فرآیند تشخیص بالینی.
 شکل ۱.۲۹ سیستم عصبی مرکزی و محیطی و محل های احتمالی ضایعه پاتولوژیک.
شکل ۱.۲۹ سیستم عصبی مرکزی و محیطی و محل های احتمالی ضایعه پاتولوژیک.
اتیولوژی بیماریهای عصبی
اختلالات سیستم عصبی عضلانی چهار نوع عمده هستند (شکل ۱.۳۰) در رابطه با علت یا علت.
 شکل ۱.۳۰ چهار دسته عمده اختلالات سیستم عصبی عضلانی.
شکل ۱.۳۰ چهار دسته عمده اختلالات سیستم عصبی عضلانی.
برای هر یک از علل اصلی بیماری انواع تحقیقات مناسبی وجود دارد که منجر به اشکال خاصی از درمان میشود. این چهار علت به ترتیب اولویت بالینی رتبهبندی میشوند تا شرایطی که شایع، بالقوه تهدیدکننده زندگی هستند و با درمان سریع قابل برگشت هستند، ابتدا ایجاد یا حذف شوند. شرایطی که نادر، مزمن و غیرقابل درمان هستند را میتوان بعدا در نظر گرفت.
اختلالات بیرونی
اختلالات بیرونی منجر به فشردگی مغز، نخاع، ریشههای عصبی و اعصاب محیطی میشود (شکل ۱.۳۱) و بنابراین با جراحی قابل درمان هستند. قبل از مداخله جراحی مغز و اعصاب، بررسیهایی مانند تصویربرداری نورورادیولوژیک سیستم عصبی مرکزی باید به سرعت انجام شود. تاخیر در جراحی مغز و اعصاب رفع فشار میتواند منجر به فلج دائمی، از دست دادن حس و بی اختیاری شود.
 شکل ۱.۳۱ اختلالات بیرونی سیستم عصبی عضلانی.
شکل ۱.۳۱ اختلالات بیرونی سیستم عصبی عضلانی.
مغز، نخاع و اعصاب محیطی میتوانند در اثر بیماری ساختارهای مجاور تحت فشار قرار گیرند. مغز ممکن است در سطح بیرونی خود توسط لختههای خون (هماتوم)، آبسهها و تومورهایی که در داخل جمجمه و پوششهای مخ ایجاد میشوند، فشرده شود. از طرف دیگر، زمانی که انسداد جریان CSF منجر به افزایش فشار و انبساط بطنها (هیدروسفالی) میشود، بطنهای پر از مایع ممکن است مغز را از درون فشرده کنند.
طناب نخاعی ممکن است در اثر بیماریهای ستون فقرات، مانند آرتریت (اسپوندیلوز)، افتادگی دیسکهای بین مهره ای و تومورهای استخوانی، و همچنین توسط تومورهای مننژ (مننژیوم) فشرده شود. کانال مرکزی نخاع، معمولاً یک فضای وستیژیال کوچک است، ممکن است به یک حفره (syrinx) منبسط شود و رشتههای عصبی را در مرکز طناب (سیرنگومیلیا) فشرده کند.
اعصاب جمجمه ای که از ساقه مغز بیرون میآیند ممکن است تحت فشار قرار گیرند، زیرا از طریق جمجمه عبور میکنند و سوراخ جمجمه را ترک میکنند، توسط تومورها و شریانهای متورم (آنوریسم). ریشههای عصبی نخاعی که نخاع را در گردن و پشت ترک میکنند ممکن است توسط تومورها و افتادگی دیسکهای بین مهره ای به دام افتاده و باعث درد، ضعف و از دست دادن حسی در ناحیه توزیع آنها شود (رادیکولوپاتی). اعصاب محیطی ممکن است در مکانهای فشار آسیبپذیر در اندامها توسط دندهها و نوارهای فیبری سخت به دام بیفتند که منجر به درد، ضعف و از دست دادن حسی در توزیع آنها میشود (نوروپاتی به دام افتادن).
تحقیقات برای اختلالات بیرونی عمدتاً نوررادیولوژیک (مانند توموگرافی کامپیوتری (CT) اسکن مغز و تصویربرداری رزونانس مغناطیسی (MRI)) برای تعیین اختلالات (ضایعات) برای رفع فشار جراحی مغز و اعصاب است. ممکن است برای جلوگیری از ناتوانی دائمینیاز به جراحی فوری باشد و به همین دلیل است که اختلالات بیرونی باید اولین ملاحظات تشخیصی باشد.
اختلالات سیستمیک
اختلالات سیستمیک در درجه اول اختلالات اندامهایی غیر از سیستم عصبی هستند که عملکرد عصبی عضلانی را با متابولیسم غیر طبیعی مختل میکنند (شکل ۱.۳۲). بیمار با یک بیماری یا سندرم عصبی مراجعه میکند، اما علت در درجه اول در جای دیگری نهفته است. ممکن است مسمومیت با داروها (مثلاً الکل)، کمبود رژیم غذایی (مثلاً ویتامین B)، نارسایی سیستم قلبی تنفسی، کبد یا کلیه، یا اختلالات هورمونی (غدد درون ریز) مانند بیماری تیروئید، دیابت شیرین و ناهنجاری در تعادل کلسیم و پتاسیم باشد.تحقیقات برای بیماری سیستمیک عمدتاً آزمایشهای خونی، بیوشیمیایی و اقدامات خاص عملکرد قلبی تنفسی، کبدی، کلیوی و غدد درون ریز است. درمان بیماری سیستمیک توسط متخصص مناسب میتواند منجر به درمان اختلال عصبی شود.
 شکل ۱.۳۲ اختلالات سیستمیک سیستم عصبی عضلانی.
شکل ۱.۳۲ اختلالات سیستمیک سیستم عصبی عضلانی.
اختلالات عروقی
اختلالات عروقی (شکل ۱.۳۳) به چند روش به گردش خون در سیستم عصبی آسیب میرساند:
![]() انسداد عروق (ترومبوز)
انسداد عروق (ترومبوز)
![]() محدودیت تامین خون و اکسیژن (انفارکتوس)
محدودیت تامین خون و اکسیژن (انفارکتوس)
![]() خونریزی در بافتهای عصبی (خونریزی).
خونریزی در بافتهای عصبی (خونریزی).
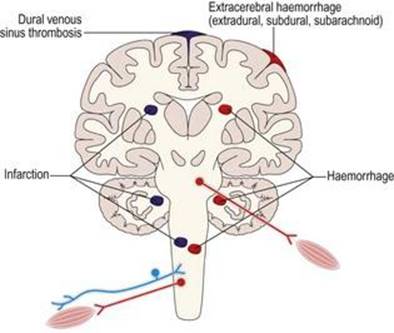 شکل ۱.۳۳ اختلالات عروقی سیستم عصبی عضلانی.
شکل ۱.۳۳ اختلالات عروقی سیستم عصبی عضلانی.
رشد سریع ضایعه عروقی را سکته مینامند. تورمهای مادرزادی شریانها (آنوریسم) یا تومورهای عروق خونی (آنژیوم) میتوانند اعصاب جمجمه و خود مغز را تحت فشار قرار دهند. بررسی اختلالات عروقی با هدف حذف اختلالات انعقادی غیرطبیعی در گردش خون، آزمایش دریچهها و عضلات قلب (اکوکاردیوگرافی، الکتروکاردیوگرافی و آنژیوگرافی قلب) و نمایش عروق گردن و مغز توسط آنژیوگرافی انجام میشود. درمان اختلالات عروقی ممکن است خونی یا قلبی باشد و ممکن است به جراحی قلب یا شریانهای گردن و جمجمه نیاز داشته باشد.
اختلالات درونی
اختلالات درونی (شکل ۱.۳۴) اختلالات اولیه خود سیستم عصبی هستند. اختلالات عصبی اولیه درونی ناشایع و اغلب مزمن و غیرقابل برگشت هستند، به طوری که میتوان یک سری تحقیقات آرامتر را انتخاب کرد. بسیاری از اختلالات عصبی تحت تأثیرات ژنتیکی قرار دارند (هرادو-دژنراسیون). خطاهای ذاتی متابولیسم منجر به ناهنجاری ذهنی و ناتوانی در کودکان میشود و معمولاً ناشی از کمبود آنزیمهای خاص است. اختلالات حمله ای شامل از دست دادن اپیزودیک هوشیاری (صرع)، خواب زیاد (نارکولپسی) و سردرد (میگرن) است. انحطاط سیستم (شکل ۱.۳۵) در جوانی و پیری رخ میدهد و منجر به مرگ زودرس (آتروفی) برخی از اجزای عصبی عضلانی، با صرفه جویی در سایر اجزا میشود. هنگامیکه انحطاط سیستم در جوانی رخ میدهد، اغلب یک علت ارثی یا ژنتیکی آشکار دارد: به عنوان مثال، دیستروفیهای عضلانی، نوروپاتیهای حسی حرکتی ارثی، پاراپارزی اسپاستیک ارثی، آتاکسیهای مخچه و بیماریهانتینگتون. هنگامیکه آنها در اواخر زندگی رخ میدهند اغلب پراکنده هستند: به عنوان مثال بیماری نورون حرکتی، بیماری پارکینسون و بیماری آلزایمر.انحطاط سیستم به طور قابل ملاحظه ای انتخابی هستند. به عنوان مثال، در بیماری نورون حرکتی فلج عضله وجود دارد اما هیچ اختلالی در حس وجود ندارد، در حالی که در بیماری آلزایمر، فراموشی شدید اما فلج وجود ندارد.
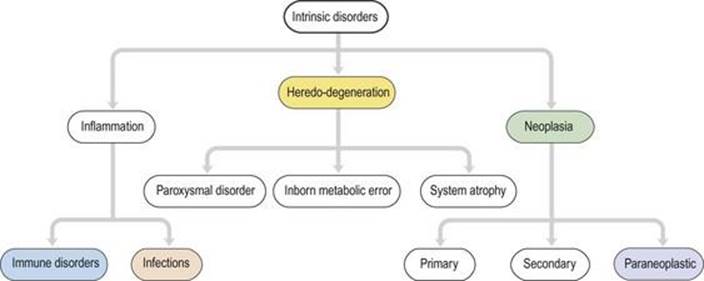 شکل ۱.۳۴ اختلالات درونی سیستم عصبی عضلانی. رنگها به شکلهای ۱.۳۵ – ۱.۳۹ مراجعه میکنند.
شکل ۱.۳۴ اختلالات درونی سیستم عصبی عضلانی. رنگها به شکلهای ۱.۳۵ – ۱.۳۹ مراجعه میکنند.
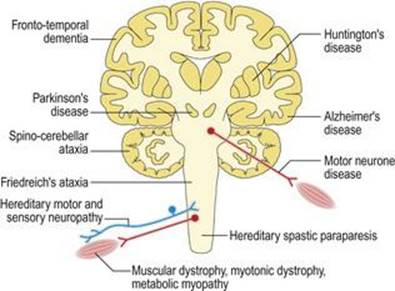 شکل ۱.۳۵ اختلالات دژنراتیو سیستم عصبی عضلانی.
شکل ۱.۳۵ اختلالات دژنراتیو سیستم عصبی عضلانی.
اصول اولیه تشخیص بالینی
![]() گرفتن شرح حال، معاینه بالینی و بررسیها منجر به تشخیص علت (علت شناسی) بیماری میشود.
گرفتن شرح حال، معاینه بالینی و بررسیها منجر به تشخیص علت (علت شناسی) بیماری میشود.
![]() محل ضایعه (ها) نشانگان بالینی آشکار شده توسط معاینه عصبی را تعیین میکند.
محل ضایعه (ها) نشانگان بالینی آشکار شده توسط معاینه عصبی را تعیین میکند.
![]() اختلالات درونی شامل انحطاط سیستم (آتروفی)، خطاهای ذاتی متابولیسم، اختلالات حمله ای، نئوپلاسمها، عفونتها و اختلالات ایمنی است.
اختلالات درونی شامل انحطاط سیستم (آتروفی)، خطاهای ذاتی متابولیسم، اختلالات حمله ای، نئوپلاسمها، عفونتها و اختلالات ایمنی است.
نئوپلازی به رشد بیش از حد و کنترل نشده بافتها اشاره دارد که یک تومور خوش خیم یا بدخیم را تشکیل میدهند. نئوپلاسمهای اولیه در خود بافتهای عصبی عضلانی ایجاد میشوند (شکل ۱.۳۶). نئوپلاسمهای ثانویه در گردش خون از سایر مکانهای اندام اولیه (مانند ریه یا سینه) پخش میشوند. به ندرت، تومورها در مکانهای دور توسط مکانیسمهای هومورال یا ایمنی به سیستم عصبی آسیب میرسانند و اختلالات ناشی از آن سندرمهای غیر متاستاتیک یا پارانئوپلاستیک نامیده میشوند (شکل ۱.۳۷).
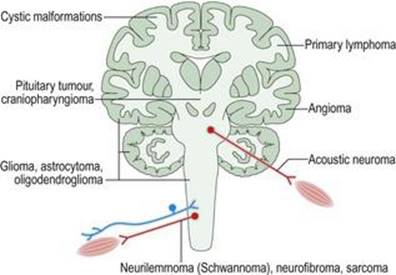 شکل ۱.۳۶ نئوپلازی سیستم عصبی عضلانی.
شکل ۱.۳۶ نئوپلازی سیستم عصبی عضلانی.
 شکل ۱.۳۷ سندرمهای پارانئوپلاستیک سیستم عصبی عضلانی.
شکل ۱.۳۷ سندرمهای پارانئوپلاستیک سیستم عصبی عضلانی.
التهاب بافت عصبی عضلانی ممکن است ناشی از عفونت توسط میکروارگانیسمها باشد (شکل ۱.۳۸) و میتواند ساختارهای مختلفی را تحت تاثیر قرار دهد: به عنوان مثال مننژها (مننژیت مننگوکوکی)، مغز (آنسفالیت ویروسی، نوروسیفلیس) یا اعصاب محیطی (جذام). متناوبا، التهاب میتواند در اختلالات ایمنی رخ دهد (شکل ۱.۳۹)، در غیاب عفونت. شایع ترین اختلال سیستم ایمنی CNS، مولتیپل اسکلروزیس است. اختلالات ایمنی ممکن است به اعصاب محیطی نیز ضربه بزند (نوروپاتی التهابی حاد یا سندرم گیلن – باره).اتصال عصبی عضلانی (میاستنی گراویس) یا ماهیچه (پلی میوزیت). اختلالات التهابی با آزمایشات میکروبیولوژیکی و سرولوژیکی خون و CSF بررسی میشود. درمان عفونت با عوامل ضد میکروبی و سرکوب پاسخهای ایمنی توسط داروهایی مانند کورتیکواستروئیدها ممکن است این بیماریهای عفونی یا ایمنی را درمان یا کنترل کند.
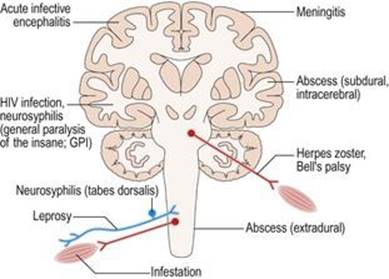 شکل ۱.۳۸ عفونتهای سیستم عصبی عضلانی.
شکل ۱.۳۸ عفونتهای سیستم عصبی عضلانی.
 شکل ۱.۳۹ اختلالات ایمنی سیستم عصبی عضلانی.
شکل ۱.۳۹ اختلالات ایمنی سیستم عصبی عضلانی.
دوره زمانی بیماری
گرفتن شرح حال میتواند در نشان دادن علت احتمالی بیماری با تعیین سرعت تکامل این اختلال، که اغلب مشخصه علل مختلف است، ارزشمند باشد (شکل ۱.۴۰). اختلالات با شروع ناگهانی (حاد) در اثر آسیب خارجی (تروما) یا یک حادثه عروقی (سکته مغزی) ایجاد میشود. هنگامیکه این بیماری در طی چند روز (تحت حاد) ایجاد میشود تا در حدود ۱ هفته به حداکثر میرسد، این به شدت نشان دهنده یک اختلال التهابی است که ممکن است عفونی (مانند مننژیت) یا ایمنی (مانند مولتیپل اسکلروزیس) باشد. بهبودی از اختلالات ایمنی هفتهها یا ماهها طول میکشد یا ناقص است. علاوه بر این، اختلالات ایمنی اغلب یک دوره عودکننده و فروکش کننده دارند، و رویدادهای حاد بر کاهش مزمن در طول ماهها و سالها قرار میگیرند. این دوره حمله ای بسیار مشخصه مولتیپل اسکلروزیس است. از این رو نام این بیماری است. در مقابل، انحطاطهای سیستمیسالها طول میکشد تا ایجاد شوند (مزمن) و شروع آن اغلب دشوار است، به خصوص که بیماران به کندی با ناتوانی انباشته خود سازگار میشوند. نئوپلاسمها یا تومورها معمولاً طی ماهها یا سالها با علائمیمانند تشنج صرع و سردرد ایجاد میشوند. به ندرت تومورهای بسیار بدخیم (گلیوما و تومورهای ثانویه) طی روزها یا هفتهها خود را نشان میدهند.
 شکل ۱.۴۰ تاریخچه (شروع) علل احتمالی را محدود میکند.
شکل ۱.۴۰ تاریخچه (شروع) علل احتمالی را محدود میکند.
محل ضایعه و سندرمهای بالینی
علت ضایعه هر چه باشد، محل آن در سیستم عصبی عضلانی منجر به یک سندرم مشخصه میشود. این از نظر بالینی با بررسی دقیق اعصاب جمجمه، سیستم حرکتی، رفلکسها، حس و هماهنگی تعریف میشود. در این کتاب، وضعیت عملکردی این سیستمها به صورت نموداری نشان داده شده است (شکل ۱.۴۱) به طوری که سندرمهای تولید شده توسط انواع خاصی از ضایعات را میتوان به شکلی تصویری نشان داد که آناتومی عصبی را با علائم بالینی پیوند میدهد. در فصلهای مناسب، شرح آناتومی و عملکردهای اساسی سیستم عصبی با شکلهایی همراه است که بر اساس شکل اولیه ۱.۴۱ است.، که سندرمهای بالینی اصلی را از نظر نقص حسی و حرکتی خلاصه میکند. به منظور درک کامل رابطه بین نوروناتومیو علائم بالینی، لازم است مسیرهای مسیرهای حسی و حرکتی اصلی، اهمیت ضایعات”نورون حرکتی تحتانی”و”نورون حرکتی فوقانی”و عملکرد کلی مخچه، عقدههای قاعده ای و قشر مخ.
 شکل ۱.۴۱ شکل اولیه برای نشان دادن سندرمهای اصلی سیستم عصبی عضلانی. در سرتاسر متن، طرحهای سادهشده سندرمهای آناتومیکی- بالینی به صورت نموداری به دنبال معاینه عصبی مرسوم اعصاب جمجمه، عملکرد حرکتی، رفلکسها، حس، هماهنگی و حالت روانی که در کتابهای درسی روشهای بالینی شرح داده شدهاند، ارائه میشوند.
شکل ۱.۴۱ شکل اولیه برای نشان دادن سندرمهای اصلی سیستم عصبی عضلانی. در سرتاسر متن، طرحهای سادهشده سندرمهای آناتومیکی- بالینی به صورت نموداری به دنبال معاینه عصبی مرسوم اعصاب جمجمه، عملکرد حرکتی، رفلکسها، حس، هماهنگی و حالت روانی که در کتابهای درسی روشهای بالینی شرح داده شدهاند، ارائه میشوند.
اطلاعات عصبی آناتومیک موجود در این فصل نشان دهنده دانش حداقل و ضروری مورد نیاز قبل از رویکرد بالینی به بیمار عصبی است. بدون این دانش، تفسیر اهمیت علائم ناشی از معاینه سیستم عصبی همانطور که در متون استاندارد در مورد”روشهای بالینی”توضیح داده شده است، غیرممکن است. تمرین مداوم معاینه سیستم عصبی از طریق تجربه، و توسعه هوش بالینی، امکان محلی سازی بسیار دقیق ضایعات در سیستم عصبی مرکزی و محیطی را فراهم میکند. محل ضایعه ممکن است کاملاً کانونی باشد – به عنوان مثال، یک تومور در نیمکره چپ مغز، یا ممکن است نشان دهنده محلی سازی در یک سیستم عصبی آناتومیک عملکردی باشد – به عنوان مثال، نورونهای حرکتی فوقانی و تحتانی در بیماری نورون حرکتی، یا اعصاب محیطی در پلی نوروپاتی حسی حرکتی. تشریح بیشتر ضایعات کانونی تعیین این است که آیا آنها در داخل سیستم عصبی قرار دارند (ضایعات درونی) یا اینکه آیا در خارج قرار دارند و سیستم عصبی را فشرده میکنند (ضایعات بیرونی). این یک تمایز مهم است زیرا ضایعات بیرونی نشان دهنده اختلالاتی هستند که به طور بالقوه توسط جراحی مغز و اعصاب قابل اصلاح هستند.
اصول خاصی وجود دارد که میتوان از سازماندهی ساختارهای عصبی تشریح شده استخراج کرد که ارزش توضیحی بالایی برای پزشک در تعیین محل ضایعه دارند.
مسیرهای حسی اصلی
حس در تنه و اندامها از گیرندههای حسی در محیط توسط اعصاب محیطی و ریشههای عصبی به گانگلیونهای ریشه پشتی و سپس به نخاع هدایت میشود. در داخل نخاع یک واگرایی از مسیرهای حسی وجود دارد که دارای روشهای حسی مختلف هستند (شکل ۱.۲۴). مسیرهای حسی درد و دما در طناب نخاعی جدا میشوند و در مجرای اسپینوتالاموس طرف مقابل بالا میروند تا به تالاموس و از آنجا به قشر حسی طرف مقابل نیمکره مغز برسند. در مقابل، مسیرهای لمسی و حس عمقی در ستونهای پشتی (خلفی) نخاع، قبل از اینکه در ساقه تحتانی مغز جدا شوند و از طریق تالاموس به قشر حسی طرف مقابل عبور کنند، بالا میروند.
![]() الگوهای از دست دادن حس در بیماری
الگوهای از دست دادن حس در بیماری
![]() ضایعات یک طرفه نخاع یا ساقه تحتانی مغز منجر به از دست دادن حس جدا میشود.
ضایعات یک طرفه نخاع یا ساقه تحتانی مغز منجر به از دست دادن حس جدا میشود.
![]() ضایعات ساقه فوقانی مغز یا نیمکره مغز منجر به از دست دادن تمام حس در سمت مقابل بدن میشود.
ضایعات ساقه فوقانی مغز یا نیمکره مغز منجر به از دست دادن تمام حس در سمت مقابل بدن میشود.
این آرایش متفاوت به ضایعات نخاع و ساقه مغز اجازه میدهد تا ترجیحاً به یک مسیر آسیب برساند و مسیر دیگر را حفظ کند. اصطلاح از دست دادن حسی تفکیک شده به یافتههای بالینی از دست دادن انتخابی روشهای لمس و حس عمقی، با حفظ مدالیتههای درد و دما و یا بالعکس اشاره دارد. این از دست دادن انتخابی روشهای حسی ناشی از درگیری انتخابی توسط ضایعات مسیرهای عملکردی خاص برای لمس / حس عمقی یا درد / دما است. ضایعه ستونهای پشتی طناب نخاعی منجر به از دست دادن حس حس عمقی یا لمس همان طرف در زیر سطح ضایعه میشود. در مقابل، ضایعه دستگاه اسپینوتالاموس منجر به از دست دادن درد/دمای پایین تر از سطح ضایعه میشود.
به عنوان مثال، ضایعه یک طرفه نخاع قفسه سینه منجر به از دست دادن حس لامسه و حس عمقی همان طرف و از دست دادن احساس درد و دمای طرف مقابل در تنه و اندام تحتانی زیر سطح ضایعه میشود. علاوه بر این، همانطور که در زیر توضیح داده شد، یک”ضعف هرمی”همان طرف در اندام تحتانی وجود دارد. در مجموع، اینها به عنوان همیکورد یا سندرم براون سکوارد شناخته میشوند ( شکل ۸.21E را ببینید).
ضایعه انتخابی در ساقه مغز لمنیسکوس داخلی منجر به از دست دادن حس لامسه میشود، در حالی که ضایعه دستگاه تریژمینوتالاموس منجر به از دست دادن احساس درد و دما در صورت میشود. بنابراین، یافته بالینی از دست دادن حسی تفکیک شده به وجود یک ضایعه کانونی درونی در نخاع یا ساقه مغز دلالت دارد.
از آنجایی که تمام مسیرهای حسی صعودی در طناب نخاعی یا ساقه تحتانی مغز مورد بررسی قرار میگیرند، ضایعات ساقه فوقانی مغز یا نیمکره مغزی منجر به از دست دادن تمام حس در طرف مقابل (مقابل) بدن میشود. (همچنین به ضایعات طناب نخاعی در شکل ۸.21A-E، ضایعات ساقه مغز در شکل ۹.۱۴ و ضایعات یک طرفه نیمکره مغزی در شکل ۱۳.۱۹ مراجعه کنید.)
مسیرهای اصلی موتور
نورونهای حرکتی که در ساقه مغز ایجاد میشوند و اعصاب جمجمهای را تشکیل میدهند، همراه با آنهایی که شاخهای شکمینخاع را در ریشههای حرکتی اعصاب نخاعی ترک میکنند، نورونهای حرکتی تحتانی نامیده میشوند و فیبرهای عضلانی خاصی را عصببندی میکنند. بنابراین، آسیب به نورونهای حرکتی پایین منجر به ضعف (پارزی) یا فلج و تحلیل رفتن عضلات فردی میشود. ماهیچهها رفلکسهای عمیق تاندون را نشان میدهند (هیپررفلکسی) و تون خود را از دست میدهند (هیپوتونی). انقباضات خودبخودی فیبرهای عضلانی مرتبط با یک عصب حرکتی واحد (واحد حرکتی) زمانی اتفاق میافتد که فیبرهای عضلانی عصبکشی میکنند و به صورت فاسیکولاسیون دیده میشوند.، یعنی حرکات موج مانند عضلات زیر پوست. از آنجایی که تمام نورونهای حرکتی تحتانی عضلات را در همان سمت بدن قرار میدهند که محل قرارگیری بدن سلول عصبی است، اثرات ضایعات نورون حرکتی تحتانی در همان طرف ضایعه دیده میشود.
مسیرهای حرکتی نزولی که فعالیت نورونهای حرکتی تحتانی را کنترل میکنند، خود به عنوان نورونهای حرکتی فوقانی شناخته میشوند. آنها از قشر مغز و ساقه مغز ناشی میشوند. مسیرهای قشر نخاعی (همی) و کورتیکوبولبار به ویژه مهم هستند (شکل ۱.۲۵). این مسیرهای حرکتی نزولی به شدت به صورت سوماتوتوپی سازماندهی شده اند اما به حرکات هماهنگ اندامها نیز مربوط میشوند. آسیب به مسیر قشر نخاعی (ضایعه نورون حرکتی فوقانی) منجر به از دست دادن حرکات فردی انگشتان و از بین رفتن حرکات کشش و ابداکشن اندام فوقانی و خم شدن اندام تحتانی میشود. این ضعف مشخصه حرکات از نظر بالینی به عنوان ضعف هرمینامیده میشود.
![]() ضایعات نورون حرکتی تحتانی و فوقانی
ضایعات نورون حرکتی تحتانی و فوقانی
آسیب به نورونهای حرکتی تحتانی با تعدادی از علائم و نشانههای حرکتی همراه است که آن را از ضایعات نورون حرکتی فوقانی متمایز میکند. تمایز بین سندرم نورون حرکتی تحتانی و فوقانی در معاینه و تشخیص عصبی بسیار مهم است. علائم بالینی آسیب به عملکرد نورون فوقانی اغلب به عنوان علائم هرمیشناخته میشود.
سندرم نورون حرکتی تحتانی
![]() ضعف (پارزیس) یا فلج (پلژی) تک تک عضلات
ضعف (پارزیس) یا فلج (پلژی) تک تک عضلات
![]() تحلیل رفتن عضلات
تحلیل رفتن عضلات
![]() انقباضات خود به خودی قابل مشاهده واحدهای حرکتی (فاسیکولاسیون)
انقباضات خود به خودی قابل مشاهده واحدهای حرکتی (فاسیکولاسیون)
![]() کاهش مقاومت در برابر کشش غیرفعال (هیپوتونی)
کاهش مقاومت در برابر کشش غیرفعال (هیپوتونی)
![]() کاهش یا از دست دادن رفلکسهای عمیق تاندون (هیپورفلکسی یا آرفلکسی).
کاهش یا از دست دادن رفلکسهای عمیق تاندون (هیپورفلکسی یا آرفلکسی).
سندرم نورون حرکتی فوقانی
![]() ضعف یا فلج حرکات خاص (کشش اندام فوقانی و خم شدن اندام تحتانی، به نام”ضعف هرمی”)
ضعف یا فلج حرکات خاص (کشش اندام فوقانی و خم شدن اندام تحتانی، به نام”ضعف هرمی”)
![]() بدون هدر رفتن عضلات
بدون هدر رفتن عضلات
![]() افزایش مقاومت در برابر کشش غیرفعال عضلات (اسپاستیسیته)؛ مقاومت اولیه در برابر کشش عضلانی و به دنبال آن آرام سازی (پاسخ با چاقو گیره)
افزایش مقاومت در برابر کشش غیرفعال عضلات (اسپاستیسیته)؛ مقاومت اولیه در برابر کشش عضلانی و به دنبال آن آرام سازی (پاسخ با چاقو گیره)
![]() بیش فعالی رفلکسهای تاندون عمیق (هیپررفلکسی)
بیش فعالی رفلکسهای تاندون عمیق (هیپررفلکسی)
![]() ظهور پاسخ اکستانسور کف پا (رفلکس بابینسکی مثبت) که منجر به خمیدگی پشتی شست پا در تحریک کف پا میشود.
ظهور پاسخ اکستانسور کف پا (رفلکس بابینسکی مثبت) که منجر به خمیدگی پشتی شست پا در تحریک کف پا میشود.
![]() از دست دادن رفلکسهای شکمی.
از دست دادن رفلکسهای شکمی.
همچنین ضعف هرمیاز نظر بالینی با رفلکسهای کشش بیش فعال تاندون (هیپررفلکسی) و با افزایش تون عضلانی (هیپرتونی)، یعنی مقاومت در برابر حرکت غیرفعال اندام، که اسپاستیسیتی نامیده میشود، همراه است. افزایش تون در کشش اولیه عضلات اندام اتفاق میافتد و سپس با شل شدن تون (پاسخ چاقوی گیره) دنبال میشود. اسپاستیسیتی در عضلات خم کننده اندام فوقانی و عضلات بازکننده اندام تحتانی ظاهر میشود. اینها همچنین گروههای عضلانی قویتر در اندامهای مربوطه هستند، و ترکیب اسپاستیسیته و قدرت بیشتر به ایجاد یک وضعیت غیرطبیعی کمک میکند که در آن دستها در حالت خمشی نسبتاً ثابت و پاها در حالت کشش قرار دارند (شکل ۱۳.۱۹ را ببینید). اندام تحتانی نیز رفلکس بابینسکی مثبت (دورسی فلکشن انگشت بزرگ در تحریک کف پا) را نشان میدهد که به عنوان پاتگنومیک آسیب دستگاه قشر نخاعی در نظر گرفته میشود.
این واقعیت که مسیرهای حرکتی نزولی در مسیرهای کورتیکوبولبار و قشر نخاعی در ساقه تحتانی مغز مورد بحث قرار میگیرند به این معنی است که یک ضایعه یک طرفه در نیمکره مغزی (نگاه کنید به شکل ۱۳.۱۹) یا ساقه مغز (نگاه کنید به شکل ۹.۱۴) منجر به فلج سمت چپ میشود..
استفاده از اصطلاح”همی”برای توصیف اثرات ضایعات نورون حرکتی فوقانی نشان میدهد که این ویژگیهای بالینی صرفاً از آسیب به دستگاه هرمییا قشر نخاعی ناشی میشود. با این حال، مجاری فردی به ندرت به صورت مجزا آسیب میبینند و بنابراین، اغلب دشوار است که نقایص بالینی را به درگیری مسیرهای خاص نسبت دهیم. آسیب به مجرای هرمیاحتمالاً دلیل از دست دادن حرکات گسسته و ظاهر رفلکس بابینسکی است.هایپررفلکسی و اسپاستیسیته به دلیل درگیری اضافی مسیرهای دیگر است.
مخچه
طرح حرکت مورد نظر از قسمتهای حرکتی قشر مخ از طریق ساقه مغز به مخچه منتقل میشود (شکل ۱.۲۷). هنگامیکه حرکت خاص انجام میشود، تکانههای آوران از گیرندههای حسی اندامها، اطلاعات مربوط به حرکت واقعی را منتقل میکنند، از طریق اعصاب محیطی به نخاع جریان مییابند و از طریق ساقه مغز به مخچه بالا میروند. بنابراین مخچه در موقعیت منحصر به فردی برای مقایسه حرکات مورد نظر با حرکات واقعی اندامها در فضا قرار دارد. هنگامیکه بین اینها اختلاف وجود داشته باشد، مخچه قادر به اصلاح حرکات انحرافی است. این امر با مسیرهای صعودی که از طریق تالاموس به قشر حرکتی و از آنجا از طریق فیبرهای نزولی که به ساقه مغز و نخاع میرسند، به دست میآید. همچنین اتصالات مخچه ای مستقیم به هستههای دهلیزی و شبکه ای ساقه مغز وجود دارد.
ضایعات مسیرهای مخچه منجر به سندرم مخچه میشود که شامل ناهماهنگی حرکات چشم (نیستاگموس)، گفتار (دیسرتری)، اندام فوقانی (لرزش قصد) و راه رفتن (آتاکسی) در غیاب ضعف یا از دست دادن حس است ( شکل را ببینید). ۹.۱۴). علائم و نشانهها در همان سمت ضایعه (همان طرف) در مخچه رخ میدهد.
ضایعه ای که مسیرهای مخچه را قطع میکند ممکن است در خود مخچه، ساقه مغز یا مسیرهای خار مخچه صعودی در نخاع قرار داشته باشد. ضایعات یک طرفه مخچه منجر به از دست دادن هماهنگی همان طرف میشود. به طور مشابه، ضایعه یک طرفه ساقه مغز به ناچار اتصالات مخچه به نیمکره مغز و نخاع را از بین میبرد و منجر به ناهماهنگی همان طرف و همانطور که در بالا توضیح داده شد، ضعف هرمیطرف مقابل اندامها میشود.
اختلالات مخچه
ضایعات مخچه باعث:
![]() نیستاگموس
نیستاگموس
![]() دیزآرتری (اسکن گفتار)
دیزآرتری (اسکن گفتار)
![]() لرزش قصد
لرزش قصد
![]() آتاکسی
آتاکسی
علائم و نشانهها در همان طرف ضایعه ظاهر میشوند.
گاهی اوقات به اشتباه تصور میشود که عدم هماهنگی اندامها مترادف با اختلال مخچه است. این مورد نیست، در حالی که این درست است که ضایعات مخچه منجر به ناهماهنگی میشود. برای مثال، بیمار با پای کوتاه و مفصل ران آرتروز، راه رفتن نامتناسب خواهد داشت. همچنین ضعف اندامها در اثر بیماری سیستم عصبی مرکزی یا محیطی باعث ناهماهنگی میشود. آسیب به اعصاب حسی محیطی یا ستونهای پشتی نخاع، مغز را از اطلاعات حس عمقی از اندامها محروم میکند، در نتیجه باعث عدم هماهنگی بازوها و راه رفتن آتاکسیک میشود. این به عنوان”آتاکسی آنسوری“شناخته میشود. هنگامیکه بیماران مبتلا به آتاکسی حسی چشمان خود را میبندند به راحتی تعادل خود را از دست میدهند و این به عنوان علامت رومبرگ شناخته میشود.با ضایعات مسیرهای مخچه این اتفاق نمیافتد.
به دلیل این مشکلات تفسیر، انجام وظایف هماهنگی در پایان معاینه عصبی به منظور ارزیابی سهم ناهنجاریهای ارتوپدی، ضعف عصبی و از دست دادن حسی در درجه ناهماهنگی، مرسوم است. اگر بتوان این نقصهای قبلی را در معاینه حذف کرد، ناهماهنگی را میتوان به طور قابل اعتمادی به ضایعات خود مسیرهای مخچه متهم کرد. این گاهی اوقات میتواند یک تمرین دشوار باشد. به عنوان مثال، در بیماریهایی مانند مولتیپل اسکلروزیس، ضایعات متعددی در مخچه، ساقه مغز و نخاع وجود دارد که هر کدام در ماهیت و درجه ناتوانی عصبی نقش دارند.
عقدههای قاعده ای
عقدههای قاعده ای، که در عمق نیمکره مغز قرار دارند، اطلاعات حسی و حرکتی را از تمام قسمتهای قشر مغز دریافت میکنند (شکل ۱.۲۶).) و همچنین از ساقه مغز و نخاع. توصیف مختصر عملکرد آنها دشوار است، اما ممکن است به عنوان ساختارهایی در نظر گرفته شوند که حرکات مفید و هدفمند را تسهیل میکنند و حرکات ناخواسته را مهار میکنند. آنها همچنین در کنترل وضعیت و تون عضلانی مهم هستند. این عملکردها با علائم اختلال عملکرد گانگلیون پایه، که طیف وسیعی از تظاهرات را پوشش میدهد، نشان داده میشوند. ضایعات عقدههای قاعده ای منجر به از دست دادن حس، قدرت یا هماهنگی نمیشود. در عوض، از دست دادن کنترل حرکات و وضعیت بدنی ارادی و تغییر در تون عضلانی وجود دارد. ضایعات یک طرفه عقدههای قاعده ای منجر به اختلال حرکتی طرف مقابل میشود.
یکی از انتهای طیف بیماریهای عقدههای قاعده ای را بیماری پارکینسون نشان میدهد. این در درجه اول با کندی در شروع و اجرای حرکت مشخص میشود (آکینزی). همچنین یک لرزش آرام و ریتمیک مشخصه در حالت استراحت وجود دارد . همچنین تون عضلانی (هیپرتونی، سفتی) اندامها افزایش مییابد. سفتی مشخصه اختلالات عقدههای قاعده ای است و به صورت مقاومت در برابر حرکت غیرفعال در تمام طول اندام ظاهر میشود. با اسپاستیسیته متفاوت است، که در آن مقاومت اولیه در برابر حرکت غیرفعال اندام و به دنبال آن آرامش وجود دارد (به بالا مراجعه کنید). در برخی از اشکال بیماری گانگلیون پایه، تون عضلانی کاهش مییابد (هیپوتونی).
انتهای دیگر طیف بیماریهای عقدههای قاعده ای را بیماریهانتینگتون و دیسکینزی ناشی از لوودوپا نشان میدهد. در این شرایط حرکات غیر ارادی غیر طبیعی (دیسکینزیا) ظاهر میشود که به شکل قطعات نامناسب و شبه هدفمند حرکت طبیعی (کریا) است. دیستونی با وضعیتهای غیر طبیعی نسبتا ثابت مشخص میشود و ممکن است با پیچش آهسته و مارپیچ اندامها (اتتوز) همراه باشد. در سندرم تورت حرکات تکراری کلیشه ای کوتاهی به نام تیک رخ میدهد. میوکلونوس با تکانهای عضلانی ناگهانی و شوکمانند مشخص میشود، اگرچه پاتوفیزیولوژی آن اغلب با عقدههای پایه ارتباطی ندارد.
![]() اختلالات عقدههای قاعده ای
اختلالات عقدههای قاعده ای
ضایعات گانگلیون پایه باعث:
![]() شروع و اجرای آهسته حرکت (آکینزی)
شروع و اجرای آهسته حرکت (آکینزی)
![]() افزایش تون عضلانی (سفتی)
افزایش تون عضلانی (سفتی)
![]() حرکات غیر ارادی غیر طبیعی (دیسکینزی).
حرکات غیر ارادی غیر طبیعی (دیسکینزی).
علائم و نشانهها در مقابل ضایعه رخ میدهد.
عملکردهای عصب روانشناختی
کارکردهای عصب روانشناختی زبان، ادراک، تحلیل فضایی، حرکات ماهر آموخته شده، حافظه و حل مسئله (یا کارکردهای اجرایی) در نیمکرههای مغزی سازماندهی میشوند (شکل ۱.۴۲). بر این اساس، ضایعات ساقه مغز، مخچه و نخاع با نقص روانی همراه نیستند. سازماندهی عملکردهای عصب روانشناختی در نیمکره مغز مانند سیستم حرکتی و حسی بسیار محلی است.
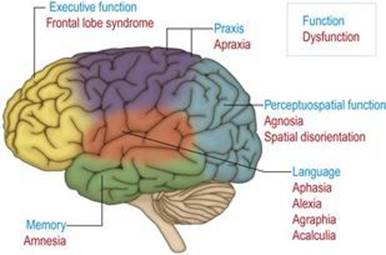 شکل ۱.۴۲ محلی سازی عملکردهای عصبی روانشناختی در نیمکره مغزی و سندرمهای مرتبط با اختلال عملکرد.
شکل ۱.۴۲ محلی سازی عملکردهای عصبی روانشناختی در نیمکره مغزی و سندرمهای مرتبط با اختلال عملکرد.
کارکردهای زبان (گفتار، خواندن، نوشتن و محاسبه) در نواحی لوبهای پیشانی، جداری و تمپورال مجاور شکاف جانبی چپ، به اصطلاح”ناحیه زبان“سازماندهی میشوند. در حالی که فرآیندهای بصری اولیه در لوبهای پس سری سازماندهی میشوند، ادراک یا تشخیص اشیاء و چهرههای انسان به صورت پیشبینی به لوبهای زمانی مغز سازماندهی میشود. توانایی فضایی جهت حرکت اندامها و بدن در فضا (عملکرد دیداری فضایی) از طریق برآمدگی به لوبهای جداری سازماندهی میشود. نواحی پیش حرکتی لوبهای فرونتال، از جمله ناحیه حرکتی تکمیلی که در قسمت داخلی نیمکره قرار دارد، بر اجرای حرکات آموخته شده و ماهرانه سر، گردن و اندامها حاکم است (پراکسیس). ساختارها در جنبههای داخلی لوبهای تمپورال، بخشی از سیستم لیمبیک، مسئول یادگیری اطلاعات جدید و یادآوری از تجربه (حافظه) هستند. سازماندهی رفتار شامل حل مسئله و دستیابی به رفتار هدفمند (عملکرد اجرایی) در نواحی پیش پیشانی لوبهای فرونتال سازماندهی میشود.
از این همبستگیهای عصبی-روانشناختی و تشریحی استنباط میشود که ضایعات ناحیه زبان منجر به از دست دادن گفتار (آفازی)، خواندن (الکسی)، نوشتن (آگرافیا) و محاسبه (acalculia) میشود، در حالی که ضایعات قشر گیجگاهی منجر به از دست دادن میشود. ادراک (آگنوزیا) و جهت گیری فضایی (بی جهتی فضایی). از دست دادن دانش حرکات ماهر آموخته شده (آپراکسی) به دنبال ضایعات قشر پیش حرکتی است. اختلالات دو طرفه لوبهای تمپورال داخلی و سیستم لیمبیک منجر به از دست دادن عملکرد حافظه میشود (فراموشی). آسیب به قشر جلوی مغز منجر به اختلال رفتاری مشخص همراه با از دست دادن پیشاندیشی، برنامهریزی و عاطفه مناسب میشود، که در تغییر شخصیت و رفتار آشکار میشود (لوب فرونتال یا سندرم نارسا اجرایی).
بررسی بیماری عصبی عضلانی
تعریف بالینی یک سندرم خاص امکان انتخاب تحقیقات مناسب برای تایید تشخیص را فراهم میکند. تمرکز اصلی تحقیقات شامل موارد زیر است:
![]() تجزیه و تحلیل CSF
تجزیه و تحلیل CSF
![]() رادیولوژی عصبی
رادیولوژی عصبی
![]() فیزیولوژی عصبی
فیزیولوژی عصبی
![]() نوروپاتولوژی (بیوپسی).
نوروپاتولوژی (بیوپسی).
پونکسیون کمری اندازه گیری فشار CSF و جمع آوری CSF را برای آنالیزهای باکتریولوژیکی، بیوشیمیایی، سرولوژیکی و سیتولوژیک ممکن میسازد. اینها ممکن است وجود خون (خونریزی زیر عنکبوتیه)، عفونت، بیماری ایمنی مانند مولتیپل اسکلروزیس یا وجود سلولهای تومور را نشان دهند.
نورورادیولوژی شامل تعدادی تکنیک است که میتواند برای به دست آوردن تصاویر ساختاری و عملکردی از سیستم عصبی مرکزی و ساختارهای اطراف استفاده شود. تصویربرداری معمولی با اشعه ایکس بر روی جمجمه و ستون مهرهها اعمال میشود، در حالی که تصاویر ساختاری مغز و نخاع با استفاده از توموگرافی کامپیوتری (CT؛ شکل ۱.۴۳) و تصویربرداری تشدید مغناطیسی (MRI؛ شکل. ۱.۴۴) به دست میآید. تصاویر عملکردی از جریان خون منطقهای مغز، متابولیسم مغزی و اتصال لیگاندهایی مانند داروها به مغز را میتوان با استفاده از توموگرافی کامپیوتری با گسیل تک فوتون بهدست آورد (SPECT; شکل ۱.۴۵) و توموگرافی گسیل پوزیترون (PET). در رادیولوژی متضاد، یک محیط مات به شریانها یا وریدها (آنژیوگرافی) تزریق میشود تا رگهای خونی را مشخص کند (شکل ۱.۴۶).
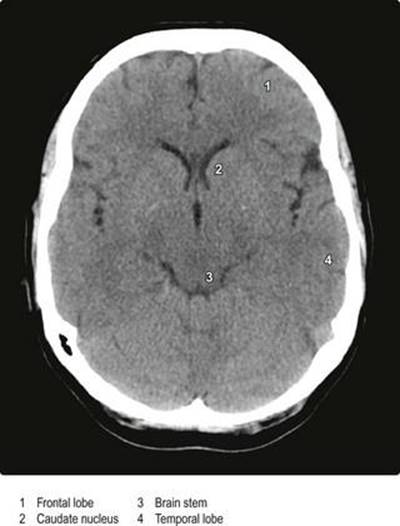 شکل ۱.۴۳ اسکن محوری کامپیوتری (CT) سر.
شکل ۱.۴۳ اسکن محوری کامپیوتری (CT) سر.
(با اجازه پروفسور PD Griffiths، واحد دانشگاهی رادیولوژی، دانشگاه شفیلد، شفیلد، انگلستان).
 شکل ۱.۴۴ تصویر تشدید مغناطیسی ساژیتال (MRI) سر.
شکل ۱.۴۴ تصویر تشدید مغناطیسی ساژیتال (MRI) سر.
(با اجازه پروفسور جکسون، مرکز تصویربرداری مولکولی ولفسون، دانشگاه منچستر، منچستر، انگلستان.)
 شکل ۱.۴۵ اسکن توموگرافی کامپیوتری با گسیل تک فوتون محوری (SPECT) از سر در یک سوژه عادی. لیگاند نشاندار شده ۱۲۳ I-FP-CIT برای برچسب زدن ناقل دوپامین (DAT) در جسم مخطط استفاده شده است.
شکل ۱.۴۵ اسکن توموگرافی کامپیوتری با گسیل تک فوتون محوری (SPECT) از سر در یک سوژه عادی. لیگاند نشاندار شده ۱۲۳ I-FP-CIT برای برچسب زدن ناقل دوپامین (DAT) در جسم مخطط استفاده شده است.
(از پروفسور دی جی بروکس، دانشکده پزشکی، امپریال کالج، لندن، انگلستان.)
 شکل ۱.۴۶ نمای جانبی آنژیوگرافی کاروتید. این تصویر با تزریق یک ماده حاجب حاوی ید به صورت انتخابی به شریان کاروتید داخلی چپ به دست آمد. این کار از طریق یک کاتتر که به شریان فمورال وارد شده و سپس تحت کنترل فلوروسکوپی به داخل قوس آئورت و شریان کاروتید هدایت میشود، انجام میشود.
شکل ۱.۴۶ نمای جانبی آنژیوگرافی کاروتید. این تصویر با تزریق یک ماده حاجب حاوی ید به صورت انتخابی به شریان کاروتید داخلی چپ به دست آمد. این کار از طریق یک کاتتر که به شریان فمورال وارد شده و سپس تحت کنترل فلوروسکوپی به داخل قوس آئورت و شریان کاروتید هدایت میشود، انجام میشود.
(با اجازه پروفسور PD Griffiths، واحد دانشگاهی رادیولوژی، دانشگاه شفیلد، شفیلد، انگلستان.)
فیزیولوژی عصبی به فعالیت الکتریکی CNS توسط الکتروانسفالوگرافی (EEG) و تشخیص و اندازه گیری پاسخهای برانگیخته به محرکهای بینایی، شنوایی و حسی تنی میپردازد. تحریک مغناطیسی مرکزی مغز، اندازه گیری زمان هدایت حرکتی به نخاع و عضلات اندام را امکان پذیر میکند. در سیستم عصبی محیطی، اندازهگیریهای سرعت هدایت عصبی حرکتی و حسی و پتانسیلهای عمل حسی برانگیخته با اندازهگیری پاسخهای عضلانی فردی به انقباض ارادی و برانگیخته الکتریکی (الکترومیوگرافی) ترکیب میشوند.
بیوپسی عصب، ماهیچه و بافت مغز، فرآیند پاتوفیزیولوژیک (مانند دژنراسیون آکسون، دمیلیناسیون، دژنراسیون عضلانی) و علت شناسی (مانند التهاب، نئوپلازی) را روشن میکند.
در ادامه بخوانید:
در ادامه شرکت کنید:
در ادامه شرکت کنید:
در ادامه شرکت کنید:
در ادامه شرکت کنید:
Textbook of Clinical Neuroanatomy, 2 ed
Chapter 1. Development of the Nervous System
Chapter 2. Organization and Functions of the Nervous System
Chapter 3. Peripheral Nerves and Ganglia
Chapter 4. Receptors and Effectors
Chapter 5. Dermatomes and Muscular Activity
Chapter 6. Central Nervous System: an Overview
Chapter 7. Spinal Cord
Chapter 8. Brainstem
Chapter 9. Nuclei, Functional Components and Distribution of Cranial Nerves
Chapter 10. Cerebellum and Fourth Ventricle
Chapter 11. Diencephalon and Third Ventricle
Chapter 12. Cerebrum
Chapter 13. Basal Nuclei (Basal Ganglia)
Chapter 14. White Matter of the Cerebrum and Lateral Ventricles
Chapter 15. Blood Supply of the Brain
Chapter 16. Meninges And Cerebrospinal Fluid
Chapter 17. Somatic Motor and Sensory Pathways
Chapter 18. Special Senses and their Neural Pathways
Chapter 19. Reticular Formation and Limbic System
Chapter 20. Autonomic Nervous System