اصول علم اعصاب اریک کندل؛ مدولاسیون انتقال سیناپسی و تحریک پذیری عصبی: پیام رسان های دوم

دعای مطالعه [ نمایش ]
بِسْمِ الله الرَّحْمنِ الرَّحیمِ
اَللّهُمَّ اَخْرِجْنى مِنْ ظُلُماتِ الْوَهْمِ
خدايا مرا بيرون آور از تاريكىهاى وهم،
وَ اَكْرِمْنى بِنُورِ الْفَهْمِ
و به نور فهم گرامى ام بدار،
اَللّهُمَّ افْتَحْ عَلَيْنا اَبْوابَ رَحْمَتِكَ
خدايا درهاى رحمتت را به روى ما بگشا،
وَانْشُرْ عَلَيْنا خَزائِنَ عُلُومِكَ بِرَحْمَتِكَ يا اَرْحَمَ الرّاحِمينَ
و خزانههاى علومت را بر ما باز كن به امید رحمتت اى مهربانترين مهربانان.
» کتاب مبانی علوم اعصاب اریک کندل
» » فصل چهاردهم: مدولاسیون انتقال سیناپسی و تحریک پذیری عصبی: پیام رسانهای دوم؛ قسمت آخر
در حال ویرایش
Principles of Neural Science; Eric R. Kandel
»» Modulation of Synaptic Transmission and Neuronal Excitability: Second Messengers
The Physiological Actions of Metabotropic Receptors Differ From Those of Ionotropic Receptors
اعمال فیزیولوژیکی گیرندههای متابوتروپیک با گیرندههای یونوتروپیک متفاوت است.
Second-Messenger Cascades Can Increase or Decrease the Opening of Many Types of Ion Channels
آبشارهای پیام رسان دوم میتوانند باز شدن بسیاری از انواع کانالهای یونی را افزایش یا کاهش دهند.
G Proteins Can Modulate Ion Channels Directly
پروتئینهای G میتوانند کانالهای یونی را مستقیما تعدیل کنند
Cyclic AMP-Dependent Protein Phosphorylation Can Close Potassium Channels
فسفوریلاسیون چرخه ای پروتئین وابسته به AMP میتواند کانالهای پتاسیم را ببندد
Second Messengers Can Endow Synaptic Transmission with Long-Lasting Consequences
پیامرسان دوم میتواند پیامدهای طولانیمدتی برای انتقال سیناپسی به همراه داشته باشد
Modulators Can Influence Circuit Function by Altering Intrinsic Excitability or Synaptic Strength
تعدیل کنندهها میتوانند با تغییر تحریک پذیری ذاتی یا قدرت سیناپسی بر عملکرد مدار تأثیر بگذارند.
Multiple Neuromodulators Can Converge onto the Same Neuron and Ion Channels
تعدیل کنندههای عصبی چندگانه میتوانند به کانالهای عصبی و یونی مشابهی همگرا شوند
Why So Many Modulators?
چرا تعدیل کنندههای زیادی وجود دارد؟
Highlights
نکات برجسته
The Physiological Actions of Metabotropic Receptors Differ From Those of Ionotropic Receptors
اعمال فیزیولوژیکی گیرندههای متابوتروپیک با گیرندههای یونوتروپیک متفاوت است
Second-Messenger Cascades Can Increase or Decrease the Opening of Many Types of Ion Channels
آبشارهای پیام رسان دوم میتوانند باز شدن بسیاری از انواع کانالهای یونی را افزایش یا کاهش دهند.
The functional differences between metabotropic and ionotropic receptors reflect the differences in their properties. For example, metabotropic receptor actions are much slower than ionotropic ones (Table 14-1). The physiological actions of the two classes of receptors also differ.
تفاوتهای عملکردی بین گیرندههای متابوتروپیک و یونوتروپیک نشان دهنده تفاوت در خواص آنهاست. به عنوان مثال، عملکرد گیرنده متابوتروپیک بسیار کندتر از گیرندههای یونوتروپیک است (جدول 14-1). اعمال فیزیولوژیکی این دو دسته از گیرندهها نیز متفاوت است.
Ionotropic receptors are channels that function as simple on-off switches; their main job is either to excite a neuron to bring it closer to the threshold for firing or inhibit the neuron to decrease its likelihood to fire. Because these channels are normally confined to the postsynaptic region of the membrane, the action of ionotropic receptors is local. Metabotropic receptors, on the other hand, because they activate diffusible second messengers, can act on channels some distance from the receptor. Moreover, metabotropic receptors regulate a variety of channel types, including resting channels, ligand-gated channels, and voltage-gated channels that generate action potentials, underlie pacemaker potentials, and provide Ca2+ influx for neurotransmitter release.
گیرندههای یونوتروپیک کانالهایی هستند که به عنوان کلیدهای روشن و خاموش عمل میکنند. کار اصلی آنها یا تحریک یک نورون برای نزدیکتر کردن نورون به آستانه شلیک است یا مهار نورون برای کاهش احتمال شلیک. از آنجایی که این کانالها معمولاً به ناحیه پسسیناپسی غشاء محدود میشوند، عملکرد گیرندههای یونوتروپیک موضعی است. از طرف دیگر، گیرندههای متابوتروپیک، چون پیامرسانهای دوم قابل انتشار را فعال میکنند، میتوانند در کانالهایی در فاصلهای از گیرنده عمل کنند. علاوه بر این، گیرندههای متابوتروپیک انواع مختلفی از کانالها را تنظیم میکنند، از جمله کانالهای استراحت، کانالهای دردار با لیگاند، و کانالهای دردار ولتاژی که پتانسیلهای عمل تولید میکنند، زیربنای پتانسیلهای ضربانساز هستند و ورود +Ca2 را برای آزادسازی انتقالدهنده عصبی فراهم میکنند.
Finally, whereas transmitter binding leads to an increase in the opening of ionotropic receptor-channels, the activation of metabotropic receptors can lead to an increase or decrease in channel opening. For example, MAP kinase phosphorylation of an inactivating (A-type) K+ channel in the dendrites of hippocampal pyramidal neurons decreases channel opening and, thus, K+ current magnitude, thereby enhancing dendritic action potential firing.
در نهایت، در حالی که اتصال ترانسمیتر منجر به افزایش باز شدن کانالهای گیرنده یونوتروپیک میشود، فعال شدن گیرندههای متابوتروپیک میتواند منجر به افزایش یا کاهش باز شدن کانال شود. به عنوان مثال، فسفوریلاسیون MAP کیناز یک کانال +K غیرفعال کننده (نوع A) در دندریتهای نورونهای هرمی هیپوکامپ باعث کاهش باز شدن کانال و در نتیجه بزرگی جریان +K میشود و در نتیجه شلیک پتانسیل عمل دندریتی را افزایش میدهد.
The binding of transmitter to metabotropic receptors can greatly influence the electrophysiological properties of a neuron (Figure 14-7). Metabotropic receptors in a presynaptic terminal can alter transmitter release by regulating either Ca2+ influx or the efficacy of the synaptic release process itself (Figure 14-7A). Metabotropic receptors in the postsynaptic cell can influence the strength of a synapse by modulating the ionotropic receptors that mediate the postsynaptic potential (Figure 14-7B). By acting on resting and voltage-gated channels in the postsynaptic neuron’s cell body, dendrites, and axon, metabotropic receptor actions can also alter the resting potential, membrane resistance, length and time constants, threshold potential, action potential duration, and repetitive firing characteristics. Such modulation of the intrinsic excitability of neurons can play an important role in regulating information flow through neuronal circuits to alter behavior.
اتصال ترانسمیتر به گیرندههای متابوتروپیک میتواند تا حد زیادی بر خواص الکتروفیزیولوژیکی یک نورون تأثیر بگذارد (شکل 14-7). گیرندههای متابوتروپیک در یک پایانه پیشسیناپسی میتوانند با تنظیم ورود +Ca2 یا کارایی خود فرآیند آزادسازی سیناپسی، رهاسازی ترانسمیتر را تغییر دهند (شکل 14-7A). گیرندههای متابوتروپیک در سلول پسسیناپسی میتوانند با تعدیل گیرندههای یونوتروپیک که واسطه پتانسیل پسسیناپسی هستند بر قدرت سیناپس تأثیر بگذارند (شکل 14-7B). عملکرد گیرنده متابوتروپیک با اثر بر روی کانالهای در حال استراحت و دریچه ولتاژ در بدنه سلولی نورون پسسیناپسی، دندریتها و آکسون میتواند پتانسیل استراحت، مقاومت غشا، ثابتهای طول و زمان، پتانسیل آستانه، مدت زمان پتانسیل عمل و ویژگیهای شلیک مکرر را تغییر دهد. . چنین مدولاسیونی از تحریک پذیری ذاتی نورونها میتواند نقش مهمیدر تنظیم جریان اطلاعات از طریق مدارهای عصبی برای تغییر رفتار ایفا کند.
Table 14-1 Comparison of Synaptic Excitation Produced by the Opening and Closing of Ion Channels
جدول 14-1 مقایسه تحریک سیناپسی تولید شده توسط باز و بسته شدن کانالهای یونی



Figure 14-7 The modulatory actions of second messengers can regulate fast synaptic transmission by acting at two synaptic sites.
شکل 14-7 اعمال تعدیل کننده پیام رسانهای دوم میتواند انتقال سریع سیناپسی را با عمل در دو مکان سیناپسی تنظیم کند.
A. In the presynaptic terminal, second messengers can regulate the efficacy of transmitter release and thus the size of the fast postsynaptic potential mediated by ionotropic receptors. This can occur by altering presynaptic Ca2+ influx, either directly by modulating presynaptic voltage-gated Ca2+ channels or indirectly by modulating presynaptic K+ channels, which alters Ca2+ influx by controlling action potential duration as illustrated (and thereby the length of time Ca2+ channels remain open). Some modulatory transmitters act to directly modulate the efficacy of the release machinery.
الف. در پایانه پیشسیناپسی، پیام رسانهای دوم میتوانند اثربخشی انتشار ترانسمیتر و در نتیجه اندازه پتانسیل سریع پسسیناپسی را با واسطه گیرندههای یونوتروپیک تنظیم کنند. این میتواند با تغییر ورود +Ca2 پیشسیناپسی، یا مستقیماً با تعدیل کانالهای +Ca2 دارای ولتاژ پیشسیناپسی یا به طور غیرمستقیم با تعدیل کانالهای +K پیشسیناپسی، که ورود +Ca2 را با کنترل مدت زمان پتانسیل عمل همانطور که نشان داده شده است، تغییر میدهد (و در نتیجه مدت زمانی که کانالهای +Ca2 باز میمانند). برخی از ترانسمیترهای تعدیلی برای تعدیل مستقیم کارایی سازوکار آزادسازی عمل میکنند.
B. In the postsynaptic terminal, second messengers can alter directly the amplitude of postsynaptic potentials by modulating ionotropic receptors.
ب. در پایانه پسسیناپسی، پیام رسانهای دوم میتوانند مستقیماً دامنه پتانسیلهای پسسیناپسی را با تعدیل گیرندههای یونوتروپیک تغییر دهند.
The distinction between direct and indirect regulation of ion channels is nicely illustrated by cholinergic synaptic transmission in autonomic ganglia of the peripheral nervous system. Stimulation of the presynaptic nerve releases ACh from the nerve terminals, directly opening nicotinic ACh receptor-channels in the postsynaptic neuron, thereby producing a fast excitatory postsynaptic potential (EPSP). The fast EPSP is followed by a slow EPSP that takes approximately 100 ms to develop but then lasts for several seconds. The slow EPSP is produced by an action of ACh on metabotropic muscarinic receptors that leads to the closing of a delayed-rectifier K+ channel called the muscarine-sensitive (or M-type) K+ channel (Figure 14-8A). These voltage-gated channels, which are formed by members of the KCNQ gene family, are partially activated when the cell is at rest; as a result, the current they carry helps determine the cell resting potential and membrane resistance.
تمایز بین تنظیم مستقیم و غیرمستقیم کانالهای یونی به خوبی با انتقال سیناپسی کولینرژیک در گانگلیونهای اتونوم سیستم عصبی محیطی نشان داده شده است. تحریک عصب پیشسیناپسی، ACh را از پایانههای عصبی آزاد میکند و به طور مستقیم کانالهای گیرنده ACh نیکوتینی را در نورون پسسیناپسی باز میکند و در نتیجه یک پتانسیل پسسیناپسی تحریکی سریع (EPSP) ایجاد میکند. EPSP سریع توسط یک EPSP آهسته دنبال میشود که تقریباً 100 میلی ثانیه طول میکشد تا ایجاد شود اما سپس چند ثانیه طول میکشد. EPSP آهسته توسط یک عمل ACh بر روی گیرندههای موسکارینی متابوتروپیک تولید میشود که منجر به بسته شدن یک کانال +K یکسو کننده تاخیری به نام کانال +K حساس به موسکارین (یا نوع M) میشود (شکل 14-8A). این کانالهای دریچهدار وابسته به ولتاژ، که توسط اعضای خانواده ژن KCNQ تشکیل میشوند، زمانی که سلول در حالت استراحت است تا حدی فعال میشوند. در نتیجه، جریانی که آنها حمل میکنند به تعیین پتانسیل استراحت سلول و مقاومت غشایی کمک میکند.
The M-type K+ channel differs from other delayed- rectifier K+ channels by its much slower activation. It requires several hundred milliseconds to fully activate on depolarization. Because M-type channels are partially open at the resting potential, their closure in response to muscarinic stimulation causes a decrease in resting K+ conductance, thus depolarizing the cell (Figure 14-8B). How far will the membrane depolarize? This can be calculated using the equivalent circuit form of the Goldman equation (Chapter 9) by decreasing the gk term from its initial value. As the change in gk due to closure of M-type K+ channels is relatively modest, the depolarization at the peak of the slow EPSP is small, only a few millivolts. Nonetheless, M-type K+ channel closure by ACh can lead to a striking increase in action potential firing in response to a depolarizing input.
کانال +K نوع M به دلیل فعال شدن بسیار کندتر با سایر کانالهای +K یکسو کننده تاخیری متفاوت است. برای فعال شدن کامل در دپلاریزاسیون به چند صد میلی ثانیه نیاز دارد. از آنجایی که کانالهای نوع M در پتانسیل استراحت تا حدی باز هستند، بسته شدن آنها در پاسخ به تحریک موسکارینی باعث کاهش رسانایی +K در حالت استراحت میشود و در نتیجه سلول را دپولاریزه میکند (شکل 14-8B). غشا تا چه اندازه دپلاریزه میشود؟ این را میتوان با استفاده از شکل مدار معادل معادله گلدمن (فصل 9) با کاهش عبارت gK از مقدار اولیه آن محاسبه کرد. از آنجایی که تغییر در gk به دلیل بسته شدن کانالهای +K نوع M نسبتاً کم است، دپلاریزاسیون در اوج EPSP آهسته کوچک است، فقط چند میلی ولت. با این وجود، بسته شدن کانال +K نوع M توسط ACh میتواند منجر به افزایش چشمگیر شلیک پتانسیل عمل در پاسخ به ورودی دپلاریزاسیون شود.
What are the special properties of M-type K+ channel closure that dramatically enhance excitability? First, the depolarization resulting from the reduction in resting & drives the membrane closer to threshold. Second, the increase in membrane resistance decreases the amount of excitatory current necessary to depolarize the cell to a given voltage. Third, the reduction in the delayed K+ current enables the cell to produce a more sustained firing of action potentials in response to a prolonged depolarizing stimulus.
ویژگیهای خاص بسته شدن کانال +K نوع M که به طور چشمگیری تحریک پذیری را افزایش میدهد چیست؟ اول، دپلاریزاسیون ناشی از کاهش استراحت و غشاء را به آستانه نزدیکتر میکند. دوم، افزایش مقاومت غشاء، مقدار جریان تحریکی لازم برای دپلاریزه کردن سلول به یک ولتاژ معین را کاهش میدهد. سوم، کاهش در جریان +K تاخیری، سلول را قادر میسازد تا در پاسخ به یک محرک دپلاریزاسیون طولانیمدت، پتانسیلهای کنش پایدارتری تولید کند.
In the absence of ACh, a ganglionic neuron normally fires only one or two action potentials and then stops firing in response to prolonged excitatory stimulation that is just above threshold. This process, termed spike-frequency adaptation, results in part from the increase in M-type K+ current in response to the prolonged depolarization, which helps repolarize the membrane below threshold. As a result, if the same prolonged stimulus is applied during a slow EPSP (when the M-type K+ channels are closed), the neuron remains depolarized above threshold during the entire stimulus and thus fires a prolonged burst of impulses (Figure 14-8C). As this modulation by ACh illustrates, the M-type K+ channels do more than help set the resting potential-they also control excitability.
در غیاب ACh، یک نورون گانگلیونی به طور معمول تنها یک یا دو پتانسیل عمل را شلیک میکند و سپس در پاسخ به تحریک تحریکی طولانی مدت که درست بالاتر از آستانه است، شلیک را متوقف میکند. این فرآیند که سازگاری با فرکانس سنبله نامیده میشود، تا حدی ناشی از افزایش جریان +K نوع M در پاسخ به دپلاریزاسیون طولانیمدت است که به قطبش مجدد غشاء زیر آستانه کمک میکند. در نتیجه، اگر همان محرک طولانیمدت در طول یک EPSP آهسته (زمانی که کانالهای +K نوع M بسته میشوند) اعمال شود، نورون در طول کل محرک در بالای آستانه دپلاریزه باقی میماند و بنابراین یک انفجار طولانیمدت از تکانهها را شلیک میکند (شکل 14-8C). همانطور که این مدولاسیون توسط ACh نشان میدهد، کانالهای +K نوع M بیشتر از کمک به تنظیم پتانسیل استراحت انجام میدهند – آنها همچنین تحریک پذیری را کنترل میکنند.
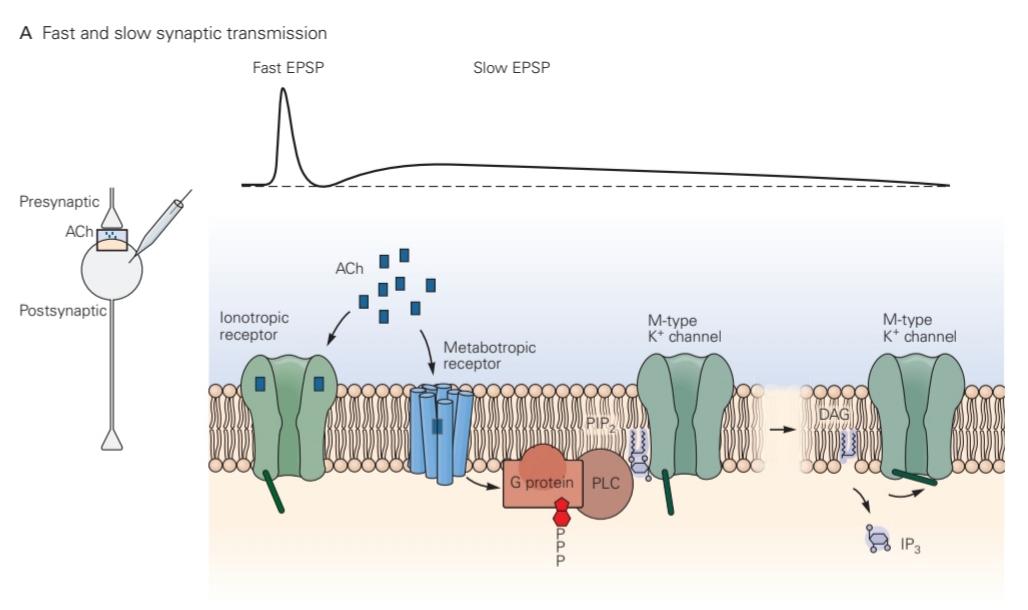

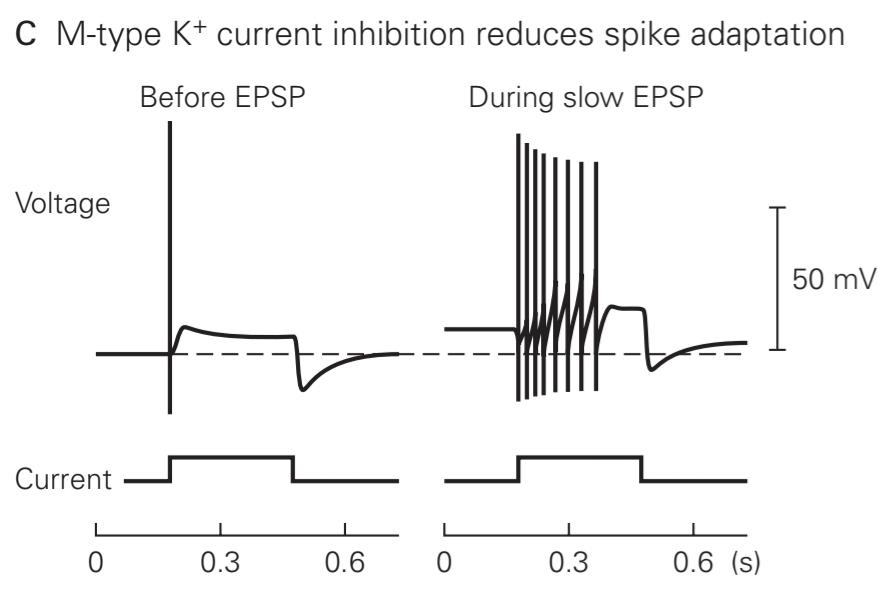
Figure 14-8 Fast ionotropic and slow metabotropic synaptic actions at autonomic ganglia.
شکل 14-8 اقدامات سیناپسی یونوتروپیک و آهسته متابوتروپیک در گانگلیونهای اتونومیک.
A. The release of ACh onto a postsynaptic neuron in autonomic ganglia produces a fast EPSP followed by a slow EPSP. The fast EPSP is produced by activation of ionotropic nicotinic ACh receptors, the slow EPSP by activation of metabotropic muscarinic ACh receptors. The metabotropic receptor stimulates PLC to hydrolyze PIP2, yielding IP3 and DAG. The decrease in PIP2 causes the closure of M-type delayed-rectifier K+ channels.
الف. آزادسازی ACh بر روی یک نورون پسسیناپسی در گانگلیونهای اتونومیک EPSP سریع و به دنبال آن EPSP آهسته تولید میکند. EPSP سریع با فعال شدن گیرندههای ACh نیکوتین یونوتروپیک، EPSP آهسته با فعال شدن گیرندههای موسکارینی متابوتروپیک ACh تولید میشود. گیرنده متابوتروپیک PLC را تحریک میکند تا PIP2 را هیدرولیز کند و IP3 و DAG را تولید کند. کاهش PIP2 باعث بسته شدن کانالهای +K یکسو کننده تاخیری نوع M میشود.
B. Voltage-clamp recordings from an autonomic ganglion neuron indicate that ACh decreases the magnitude of the cur rent carried by the voltage-gated M-type K+ channels. In this experiment, the cell is initially clamped at a holding potential (Vh) near the resting potential in the absence of ACh (typically -60 mV). At this potential, the M-type K+ channels are partially open, leading to a steady outward K+ current. The voltage is then stepped for 1 second to a more positive test potential (Vt, typically -40 mV), which normally causes a slow increase in outward K+ current (Ik) as the M-type K+ channels respond to the more positive voltage by increasing their opening (control). Application of muscarine, a plant alkaloid that selectively stimulates the muscarinic ACh receptor, causes a fraction of the M-type K+ channels to close. This decreases the outward K+ current at the holding potential (note the shift in baseline current, A), by closing the M-type K+ channels that are open at rest, and decreases the magnitude of the slowly activating K+ current in response to the step depolarization. (Adapted from Adams et al. 1986.)
B. ثبتهای کلمپ ولتاژ از یک نورون گانگلیونی خودمختار نشان میدهد که ACh مقدار جریان جریانی را که توسط کانالهای +K نوع M دریچهدار وابسته به ولتاژ منتقل میشود، کاهش میدهد. در این آزمایش، سلول ابتدا در یک پتانسیل نگهدارنده (Vh) نزدیک پتانسیل استراحت در غیاب ACH (معمولا 60- میلی ولت) گیره میشود. در این پتانسیل، کانالهای +K نوع M تا حدی باز هستند که منجر به جریان +K به بیرون ثابت میشود. سپس ولتاژ به مدت 1 ثانیه به یک پتانسیل تست مثبت تر (Vt، معمولاً 40- میلی ولت) افزایش مییابد، که معمولاً باعث افزایش آهسته در جریان +K به بیرون (Ik) میشود زیرا کانالهای +K نوع M به ولتاژ مثبت تر پاسخ میدهند. افزایش باز شدن آنها (کنترل). استفاده از موسکارین، یک آلکالوئید گیاهی که به طور انتخابی گیرنده موسکارینی ACh را تحریک میکند، باعث بسته شدن کسری از کانالهای +K نوع M میشود. این امر با بستن کانالهای +K نوع M که در حالت سکون باز هستند، جریان خروجی +K را در پتانسیل نگهدارنده کاهش میدهد (به تغییر جریان پایه، A توجه کنید)، و مقدار جریان +K را که به آرامی فعال میشود در پاسخ به مرحله دپلاریزاسیون کاهش میدهد. (برگرفته از آدامز و همکاران 1986.)
C. In the absence of muscarinic ACh receptor stimulation, the neuron fires only a single action potential in response to a prolonged depolarizing current stimulus, a process termed spike- frequency adaptation (left). This is because the slow activation of the M-type K+ channel during the depolarization generates an outward current that repolarizes the membrane below threshold. When the same current stimulus is applied during a slow EPSP, when a large fraction of M-type channels are now unable to open, the neuron fires a more sustained train of action potentials (right). (Adapted from Adams et al. 1986.)
ج. در غیاب تحریک گیرنده موسکارینی ACh، نورون تنها یک پتانسیل عمل واحد را در پاسخ به یک محرک جریان دپلاریزاسیون طولانی مدت، فرآیندی که انطباق با فرکانس سنبله (سمت چپ) نامیده میشود، شلیک میکند. این به این دلیل است که فعال شدن آهسته کانال +K نوع M در طول دپلاریزاسیون، جریانی رو به بیرون تولید میکند که غشاء را در زیر آستانه مجدداً قطبی میکند. هنگامیکه همان محرک فعلی در طول یک EPSP آهسته اعمال میشود، زمانی که بخش بزرگی از کانالهای نوع M اکنون قادر به باز شدن نیستند، نورون یک قطار پایدارتر از پتانسیلهای عمل را شلیک میکند (راست). (برگرفته از آدامز و همکاران 1986.)
Although it has been known for some time that muscarinic receptor actions in autonomic ganglia result in the activation of PLC and the production of DAG and IP3, the precise mechanism by which this signaling cascade produces M-type channel closure remained mysterious. However, it is now clear that M-channel closure upon muscarinic receptor activation is not due to the production of a second messenger. Rather, the M-channels, as well as a number of other types of channels (eg, see Figure 14-10), bind membranous PIP2, as a cofactor for their proper functioning. Thus, muscarinic receptor activation closes M-type channels by activating PLC, and thereby decreasing the levels of PIP2 in the membrane due to hydrolysis by PLC. We shall next discuss the mechanisms by which other signaling cascades are capable of modulating other types of ion channels. We start by describing the simplest mechanism, the direct gating of ion channels by G proteins, and then consider a more complex mechanism dependent on protein phosphorylation by PKA.
اگرچه مدتی است که مشخص شده است که اعمال گیرنده موسکارینی در گانگلیونهای اتونوم منجر به فعال شدن PLC و تولید DAG و IP3 میشود، مکانیسم دقیقی که توسط این آبشار سیگنالینگ باعث بسته شدن کانال نوع M میشود، مرموز باقی مانده است. با این حال، اکنون واضح است که بسته شدن کانال M پس از فعال شدن گیرنده موسکارینی به دلیل تولید یک پیام رسان دوم نیست. در عوض، کانالهای M، و همچنین تعدادی از انواع دیگر کانالها (به عنوان مثال، به شکل 14-10 مراجعه کنید)، PIP2 غشایی را به عنوان یک کوفاکتور برای عملکرد مناسب خود متصل میکنند. بنابراین، فعالسازی گیرنده موسکارینی، کانالهای نوع M را با فعال کردن PLC میبندد و در نتیجه سطح PIP2 در غشاء را به دلیل هیدرولیز توسط PLC کاهش میدهد. ما در ادامه مکانیسمهایی را مورد بحث قرار خواهیم داد که توسط آنهای دیگر آبشارهای سیگنالینگ قادر به تعدیل انواع دیگر کانالهای یونی هستند. ما با توصیف سادهترین مکانیسم، دروازهبندی مستقیم کانالهای یونی توسط پروتئینهای G، شروع میکنیم و سپس مکانیسم پیچیدهتری را که به فسفوریلاسیون پروتئین توسط PKA وابسته است، در نظر میگیریم.
G Proteins Can Modulate Ion Channels Directly
پروتئینهای G میتوانند کانالهای یونی را مستقیما تعدیل کنند
The simplest mechanism for the indirect gating of a channel occurs when transmitter binding to a metabotropic receptor releases a G protein subunit that directly interacts with the channel to modify its opening. This mechanism is used to gate two kinds of ion channels: the G protein-gated inward-rectifier K+ channels (GIRK1-4; encoded by the KCNJ1-4 genes) and a voltage-dependent Ca2+ channel. With both kinds of channels, it is the G protein’s By complex that binds to and regulates channel opening (Figure 14-9A).
سادهترین مکانیسم برای راهاندازی غیرمستقیم یک کانال زمانی اتفاق میافتد که ترانسمیتر متصل به یک گیرنده متابوتروپیک، یک زیرواحد پروتئین G آزاد میکند که مستقیماً با کانال تعامل میکند تا دهانه آن را تغییر دهد. این مکانیسم برای راهاندازی دو نوع کانال یونی مورد استفاده قرار میگیرد: کانالهای +K یکسوکننده داخلی با دروازه پروتئین G (GIRK1-4؛ کدگذاری شده توسط ژنهای KCNJ1-4) و یک کانال +Ca2 وابسته به ولتاژ. با هر دو نوع کانال، کمپلکس By پروتئین G است که به باز شدن کانال متصل شده و آن را تنظیم میکند (شکل 14-9A).
The GIRK channel, like other inward-rectifier channels, passes current more readily in the inward than the outward direction, although in physiological situations, K+ current is always outward. Inward- rectifier channels resemble a truncated voltage-gated K+ channel in having two transmembrane regions connected by a P-region loop that forms the selectivity filter in the channel (see Figure 8-11).
کانال GIRK، مانند سایر کانالهای یکسو کننده داخلی، جریان را با سهولت بیشتری در جهت درونی از جهت بیرونی عبور میدهد، اگرچه در موقعیتهای فیزیولوژیکی، جریان +K همیشه به سمت خارج است. کانالهای یکسوکننده به داخل شبیه یک کانال +K دردار ولتاژ کوتاه هستند که دارای دو ناحیه گذر غشایی هستند که توسط یک حلقه P-region به هم متصل شدهاند که فیلتر انتخابپذیری را در کانال تشکیل میدهد (شکل 8-11 را ببینید).
In the 1920s, Otto Loewi described how the release of ACh in response to stimulation of the vagus nerve slows the heart rate (Figure 14-9B). We now know that ACh activates muscarinic receptors to stimulate G protein activity, which directly opens the GIRK channel. For many years, this transmitter action was puzzling because it has properties of both ionotropic and metabotropic receptor actions. The time course of activation of the K current following release of ACh is slower (50- to 100-ms rise time) than that of ionotropic receptors (rise time <1 ms). However, the rate of GIRK channel activation is much faster than that of second-messenger-mediated actions that depend on protein phosphorylation (which can take many seconds to turn on). Although biochemical and electrophysiological studies clearly demonstrated that a G protein was required for this action, patch-clamp experiments showed that the G protein did not trigger production of a diffusible second messenger (Figure 14-9C). These findings were reconciled when it was found that the GIRK channel was activated directly by the G protein’s βγ-subunit complex, which becomes available to interact with the GIRK channel when it dissociates from the G protein α-subunit upon activation of the muscarinic receptors.
در دهه 1920، اتو لووی توضیح داد که چگونه انتشار ACh در پاسخ به تحریک عصب واگ، ضربان قلب را کاهش میدهد (شکل 14-9B). اکنون میدانیم که ACh گیرندههای موسکارینی را برای تحریک فعالیت پروتئین G فعال میکند که مستقیماً کانال GIRK را باز میکند. برای سالهای متمادی، این عمل ترانسمیتر گیجکننده بود، زیرا دارای ویژگیهای عملکرد گیرنده یونوتروپیک و متابوتروپیک بود. دوره زمانی فعالسازی جریان پتاسیم به دنبال انتشار ACh کندتر است (زمان افزایش 50 تا 100 میلیثانیه) نسبت به گیرندههای یونوتروپیک (زمان افزایش کمتر از 1 میلیثانیه). با این حال، سرعت فعالسازی کانال GIRK بسیار سریعتر از فعالیتهای پیامرسان دوم است که به فسفوریلاسیون پروتئین بستگی دارد (که ممکن است چند ثانیه طول بکشد تا روشن شود). اگرچه مطالعات بیوشیمیایی و الکتروفیزیولوژیکی به وضوح نشان داد که یک پروتئین G برای این عمل مورد نیاز است، آزمایشات پچ گیره نشان داد که پروتئین G باعث تولید یک پیام رسان دوم قابل انتشار نمیشود (شکل 14-9C). این یافتهها زمانی که مشخص شد کانال GIRK مستقیماً توسط کمپلکس زیرواحد βγ پروتیین G فعال میشود، تطبیق داده شد، که برای تعامل با کانال GIRK زمانی که از زیرواحد α پروتئین G پس از فعال شدن گیرندههای موسکارینی جدا میشود، در دسترس قرار میگیرد.
The mechanism by which the By-subunits activate the GIRK channel was recently elucidated at the atomic resolution through the solving of the X-ray crystal structure of the GIRK channel in a complex with the By-subunits. Each of the four GIRK channel subunits binds a single By-subunit complex, which interacts with the cytoplasmic surface of the channel, leading to a conformational change that promotes channel opening (Figure 14-10).
مکانیسمی که توسط آن زیرواحدهای فرعی کانال GIRK را فعال میکنند اخیراً در وضوح اتمیاز طریق حل ساختار کریستالی اشعه ایکس کانال GIRK در مجموعهای با زیرواحدهای فرعی روشن شده است. هر یک از چهار زیر واحد کانال GIRK به یک کمپلکس زیر واحدی متصل میشود که با سطح سیتوپلاسمیکانال در تعامل است و منجر به تغییر ساختاری میشود که باعث باز شدن کانال میشود (شکل 14-10).
Activation of GIRK channels hyperpolarizes the membrane in the direction of Ek (-80 mV). In certain classes of spontaneously active neurons, the outward K+ current through these channels acts predominantly to decrease the neuron’s intrinsic firing rate, opposing the slow depolarization caused by excitatory pacemaker currents carried by the hyperpolarization-activated, cyclic nucleotide-regulated channels, which are encoded by the HCN gene family (Chapter 10). Because GIRK channels are activated by neurotransmitters, they provide a means for synaptic modulation of the firing rate of excitable cells. These channels are regulated in a wide variety of neurons by a large number of transmitters and neuropeptides that act on different G protein-coupled receptors to activate either Gi, or Go, thereby releasing the By-subunits.
فعال شدن کانالهای GIRK غشاء را در جهت Ek (80- میلی ولت) هیپرپلاریزه میکند. در کلاسهای خاصی از نورونهای فعال خود به خود، جریان +K بیرونی از طریق این کانالها عمدتاً برای کاهش سرعت شلیک ذاتی نورون عمل میکند و با دپلاریزاسیون آهسته ناشی از جریانهای ضربانساز تحریکی که توسط کانالهای تنظیمشده با نوکلئوتید حلقوی فعال شده با هیپرپلاریزاسیون، مخالفت میکند، عمل میکند. کدگذاری شده توسط خانواده ژن HCN (فصل 10). از آنجایی که کانالهای GIRK توسط انتقالدهندههای عصبی فعال میشوند، وسیلهای برای مدولاسیون سیناپسی سرعت شلیک سلولهای تحریکپذیر فراهم میکنند. این کانالها در طیف گستردهای از نورونها توسط تعداد زیادی ترانسمیتر و نوروپپتید تنظیم میشوند که روی گیرندههای مختلف جفت شده با پروتئین G عمل میکنند تا Gi یا Go را فعال کنند و در نتیجه زیرواحدهای فرعی را آزاد کنند.
Several G protein-coupled receptors also act to inhibit the opening of certain voltage-gated Ca2+ channels, again as a result of the direct binding of the βγ complex of Gi or Go to the channel. Because Ca2+ influx through voltage-gated Ca2+ channels normally has a depolarizing effect, the dual action of G protein βγ-subunits-Ca2+ channel inhibition and K+ channel activation-strongly inhibits neuronal firing. As we will see in Chapter 15, inhibition of voltage-gated Ca2+ channels in presynaptic terminals can suppress the release of neurotransmitter.
چندین گیرنده جفت شده با پروتئین G نیز برای مهار باز شدن برخی کانالهای +Ca2 دارای ولتاژ، باز هم در نتیجه اتصال مستقیم کمپلکس βγ از Gi یا Go به کانال عمل میکنند. از آنجایی که ورود +Ca2 از طریق کانالهای +Ca2 دارای ولتاژ معمولاً یک اثر دپلاریزاسیون دارد، عمل دوگانه زیرواحدهای پروتئین G- مهار کانال +Ca2 و فعالسازی کانال +K به شدت شلیک عصبی را مهار میکند. همانطور که در فصل 15 خواهیم دید، مهار کانالهای +Ca2 دارای ولتاژ در پایانههای پیشسیناپسی میتواند آزادسازی انتقال دهنده عصبی را سرکوب کند.

Figure 14-9 Some G proteins can open ion channels directly without employing second messengers.
شکل 14-9 برخی از پروتئینهای G میتوانند کانالهای یونی را مستقیماً بدون استفاده از پیام رسانهای دوم باز کنند.
A. An inward-rectifying K+ channel (GIRK) is opened directly by a G protein. Binding of ACh to a muscarinic receptor causes the Gi protein and αiβγ complex to dissociate; the free βγ-subunits bind to a cytoplasmic domain of the channel, causing the channel to open.
الف. یک کانال +K تصحیح کننده (GIRK) مستقیماً توسط یک پروتئین G باز میشود. اتصال ACh به یک گیرنده موسکارینی باعث جدا شدن پروتئین Gi و کمپلکس αiβγ میشود. زیرواحدهای βγ آزاد به دامنه سیتوپلاسمی کانال متصل میشوند و باعث باز شدن کانال می شوند.
B. Stimulation of the parasympathetic vagus nerve releases ACh, which acts at muscarinic receptors to open GIRK channels in cardiac muscle cell membranes. The current through the GIRK channel hyperpolarizes the cells, thus slowing the heart rate. (Adapted from Toda and West 1967.)
ب. تحریک عصب واگ پاراسمپاتیک باعث آزاد شدن ACh میشود که در گیرندههای موسکارینی برای باز کردن کانالهای GIRK در غشای سلولی عضله قلب عمل میکند. جریان عبوری از کانال GIRK سلولها را هیپرپلاریزه میکند و در نتیجه ضربان قلب را کاهش میدهد. (برگرفته از تودا و وست 1967.)
C. Three single-channel records show that opening of GIRK channels does not involve a freely diffusible second messenger. In this experiment, the pipette contained a high concentration of K+, which makes Ek less negative. As a result, when GIRK channels open, they generate brief pulses of inward (downward) current. In the absence of ACh, channels open briefly and infrequently (top record). Application of ACh in the bath (outside the pipette) does not increase channel opening in the patch of membrane under the pipette (middle record). This is because the free βγ-subunits, released by the binding of ACh to its receptor, remain tethered to the membrane near the receptor and can only activate nearby channels. The subunits are not free to diffuse to the channels under the patch pipette. The ACh must be in the pipette to activate the channel (bottom record). (Reproduced, with permission, from Soejima and Noma 1984. Copyright © 1984 Springer Nature.)
ج. سه رکورد تک کاناله نشان میدهد که باز کردن کانالهای GIRK شامل پیام رسان دوم آزادانه قابل انتشار نیست. در این آزمایش، پیپت حاوی غلظت بالایی از +K بود که باعث میشود Ek کمتر منفی شود. در نتیجه، زمانی که کانالهای GIRK باز میشوند، پالسهای مختصری از جریان داخل (پایین) تولید میکنند. در غیاب ACh، کانالها به طور مختصر و به ندرت باز میشوند ( رکورد بالا). استفاده از ACh در حمام (خارج از پیپت) باعث افزایش باز شدن کانال در وصله غشا زیر پیپت نمیشود (رکورد میانی). این به این دلیل است که زیرواحدهای آزاد آزاد که با اتصال ACh به گیرنده آن آزاد میشوند، به غشای نزدیک گیرنده متصل میمانند و فقط میتوانند کانالهای مجاور را فعال کنند. زیرواحدها برای انتشار به کانالهای زیر پچ پیپت آزاد نیستند. ACh باید در پیپت باشد تا کانال فعال شود (رکورد پایین). (بازتولید، با اجازه، از Soejima و Noma 1984. حق چاپ © 1984 Springer Nature.)

Figure 14-10 G protein βγ-subunits can directly bind and activate GIRK channels. A high-resolution structure of a GIRK channel (green) interacting with the G protein ẞ-subunit (Gẞ, cyan) and y-subunit (Gy, purple). A geranylgeranyl lipid molecule (gg) is attached to the C-terminus of Gy. The structure illustrates that Na+ ions and the phospholipid PIP, also bind to the channel, thereby enhancing channel opening. The pink spheres inside the channel represent K+ ions. (Adapted with permission from Whorton and MacKinnon 2013. Copyright © 2013 Springer Nature.)
شکل 14-10 زیرواحدهای βγ پروتئین G میتوانند مستقیماً کانالهای GIRK را متصل و فعال کنند. ساختاری با وضوح بالا از یک کانال GIRK (سبز) که با زیرواحد β پروتئین G (Gβ، فیروزهای) و زیرواحد γ (Gγ، بنفش) در تعامل است. یک مولکول لیپیدی geranylgeranyl (gg) به C-ترمینال Gγ متصل است. این ساختار نشان میدهد که یونهای +Na و فسفولیپید PIP نیز به کانال متصل میشوند و در نتیجه باز شدن کانال را افزایش میدهند. کرههای صورتی داخل کانال نشان دهنده یونهای +K هستند. (اقتباس شده با مجوز Whorton و MacKinnon 2013. حق چاپ © 2013 Springer Nature.)
Cyclic AMP-Dependent Protein Phosphorylation Can Close Potassium Channels
فسفوریلاسیون چرخه ای پروتئین وابسته به AMP میتواند کانالهای پتاسیم را ببندد
In the marine mollusk Aplysia, a group of mechanoreceptor sensory neurons initiates defensive withdrawal reflexes in response to tactile stimuli through fast excitatory synapses with motor neurons. Certain interneurons form serotonergic synapses with these sensory neurons, and the serotonin released by the interneurons sensitizes the withdrawal reflex, enhancing the animal’s response to a stimulus and thus producing a simple form of learning (Chapter 53).
در نرم تنان دریایی Aplysia، گروهی از نورونهای حسی گیرنده مکانیکی رفلکسهای عقبنشینی دفاعی را در پاسخ به محرکهای لمسی از طریق سیناپسهای تحریککننده سریع با نورونهای حرکتی آغاز میکنند. برخی از نورونهای داخلی با این نورونهای حسی سیناپسهای سروتونرژیک تشکیل میدهند و سروتونین آزاد شده توسط این نورونها رفلکس محرومیت را حساس میکند و پاسخ حیوان به یک محرک را افزایش میدهد و بنابراین شکل سادهای از یادگیری را تولید میکند (فصل 53).
The modulatory action of serotonin depends on its binding to a G protein-coupled receptor that activates a G, protein, which elevates cAMP and thus activates PKA. This leads to the direct phosphorylation and subsequent closure of the serotonin-sensitive (or S-type) K+ channel that acts as a resting channel (Figure 14-11). Like the closing of the M-type K channel by ACh, closure of the S-type K+ channel decreases K+ efflux from the cell, thereby depolarizing the cell and decreasing its resting membrane conductance. Conversely, the opening of the same S-type K+ channels can be enhanced by the neuropeptide FMR- Famide, acting through 12-lipoxygenase metabolites of arachidonic acid. This enhanced channel opening leads to a slow hyperpolarizing inhibitory postsynaptic potential (IPSP) associated with an increase in resting membrane conductance.
عمل تعدیلی سروتونین به اتصال آن به گیرنده جفت شده با پروتئین G بستگی دارد که پروتئین G را فعال میکند که cAMP را بالا میبرد و بنابراین PKA را فعال میکند. این منجر به فسفوریلاسیون مستقیم و بسته شدن کانال +K حساس به سروتونین (یا نوع S) میشود که به عنوان یک کانال استراحت عمل میکند (شکل 14-11). مانند بسته شدن کانال +K نوع M توسط ACh، بسته شدن کانال +K نوع S باعث کاهش جریان +K از سلول میشود و در نتیجه سلول را دپولاریزه میکند و رسانایی غشای در حال استراحت آن را کاهش میدهد. برعکس، باز شدن همان کانالهای +K نوع S را میتوان با نوروپپتید FMR-Famide افزایش داد که از طریق متابولیتهای 12-لیپوکسیژناز اسید آراشیدونیک عمل میکند. این باز شدن کانال افزایش یافته منجر به پتانسیل پسسیناپسی مهاری هیپرپلاریزه آهسته (IPSP) مرتبط با افزایش رسانایی غشاء در حالت استراحت میشود.
Thus, a single channel can be regulated by distinct second-messenger pathways that produce opposite effects on neuronal excitability. Likewise, a resting K+ channel with two pore-forming domains in each subunit (the TREK-1 channel) in mammalian neurons is dually regulated by PKA and arachidonic acid in a manner very similar to the dual regulation of the S-type channel in Aplysia.
بنابراین، یک کانال واحد را میتوان توسط مسیرهای پیام رسان دوم مجزا تنظیم کرد که اثرات متضادی بر تحریک پذیری عصبی ایجاد میکند. به همین ترتیب، یک کانال +K در حال استراحت با دو حوزه تشکیل دهنده منافذ در هر زیر واحد (کانال TREK-1) در نورونهای پستانداران به طور دوگانه توسط PKA و اسید آراشیدونیک به شیوه ای بسیار شبیه به تنظیم دوگانه کانال نوع S در Aplysia تنظیم میشود.
Second Messengers Can Endow Synaptic Transmission with Long-Lasting Consequences
پیامرسان دوم میتواند پیامدهای طولانیمدتی برای انتقال سیناپسی به همراه داشته باشد
So far, we have described how synaptic second messengers alter the biochemistry of neurons for periods lasting seconds to minutes. Second messengers can also produce long-term changes lasting days to weeks as a result of alterations in a cell’s expression of specific genes (Figure 14-12). Such changes in gene expression result from the ability of second-messenger cascades to control the activity of transcription factors, regulatory proteins that control mRNA synthesis.
تاکنون توضیح دادهایم که چگونه پیامرسانهای سیناپسی بیوشیمینورونها را برای دورههایی از ثانیه تا چند دقیقه تغییر میدهند. پیامرسانهای دوم همچنین میتوانند تغییرات طولانیمدتی را ایجاد کنند که روزها تا هفتهها طول میکشد در نتیجه تغییر در بیان ژنهای خاص سلول (شکل 14-12). چنین تغییراتی در بیان ژن ناشی از توانایی آبشارهای پیام رسان دوم برای کنترل فعالیت فاکتورهای رونویسی، پروتئینهای تنظیم کننده ای است که سنتز mRNA را کنترل میکنند.
Some transcription factors can be directly regulated by phosphorylation. For example, the cAMP response element-binding protein (CREB) is activated when phosphorylated by PKA, calcium/calmodulin- dependent protein kinases, PKC, or MAP kinases. Once activated, CREB enhances transcription by binding to specific DNA sequences, the cAMP response elements or CRE, and recruiting a component of the transcription machinery, the CREB-binding protein (CBP). CBP activates transcription by recruiting RNA polymerase II and by functioning as a histone acetylase, adding acetyl groups to certain histone lysine residues. The acetylation weakens the binding between histones and DNA, thus opening up the chromatin structure and enabling specific genes to be transcribed. The changes in transcription and chromatin structure are important for regulating neuronal development, as well as for long-term learning and memory (Chapters 53 and 54).
برخی از فاکتورهای رونویسی را میتوان مستقیماً با فسفوریلاسیون تنظیم کرد. به عنوان مثال، پروتئین متصل شونده به عنصر پاسخ cAMP (CREB) زمانی فعال میشود که توسط PKA، پروتئین کینازهای وابسته به کلسیم/کالمودولین، PKC یا MAP کیناز فسفریله شود. پس از فعال شدن، CREB رونویسی را با اتصال به توالیهای DNA خاص، عناصر پاسخ cAMP یا CRE، و به کارگیری بخشی از ماشین رونویسی، پروتئین متصل شونده به CREB (CBP) افزایش میدهد. CBP رونویسی را با به کارگیری RNA پلیمراز II و عملکرد به عنوان هیستون استیلاز فعال میکند و گروههای استیل را به باقی ماندههای هیستون لیزین اضافه میکند. استیلاسیون اتصال بین هیستونها و DNA را ضعیف میکند، در نتیجه ساختار کروماتین باز میشود و ژنهای خاص رونویسی میشوند. تغییرات در رونویسی و ساختار کروماتین برای تنظیم رشد عصبی و همچنین برای یادگیری و حافظه طولانی مدت مهم هستند (فصل 53 و 54).
Modulators Can Influence Circuit Function by Altering Intrinsic Excitability or Synaptic Strength
تعدیل کنندهها میتوانند با تغییر تحریک پذیری ذاتی یا قدرت سیناپسی بر عملکرد مدار تأثیر بگذارند.
Most of this chapter has been devoted to understanding the cellular mechanisms and signal transduction pathways that allow neuromodulator-activated pathways to alter the activity of ion channels, receptors, and synapses in individual neurons. However, in the intact brain, modulatory transmitters released either from diffuse projections over large areas of the brain (Chapter 16) or from more locally targeted connections can alter the dynamics of brain circuits in a number of important ways. In this section, we examine one well-studied example of modulatory control of circuit function-the control of crustacean feeding behavior by the neurons of the stomatogastric ganglion to illustrate the following general properties.
بیشتر این فصل به درک مکانیسمهای سلولی و مسیرهای انتقال سیگنال اختصاص داده شده است که به مسیرهای تعدیلکننده عصبی اجازه میدهد تا فعالیت کانالهای یونی، گیرندهها و سیناپسها را در تک تک نورونها تغییر دهند. با این حال، در مغز دست نخورده، ترانسمیترهای تعدیلکننده منتشر شده یا از برجستگیهای منتشر شده در مناطق بزرگ مغز (فصل 16) یا از اتصالات هدفمند محلیتر میتوانند دینامیک مدارهای مغز را به روشهای مهمیتغییر دهند. در این بخش، ما یک نمونه به خوبی مطالعه شده از کنترل تعدیلی عملکرد مدار را بررسی میکنیم – کنترل رفتار تغذیه سخت پوستان توسط نورونهای گانگلیون روزنه معده برای نشان دادن ویژگیهای کلی زیر.

Figure 14-11 Serotonergic interneurons close a K+ channel through the diffusible second-messenger cAMP. Serotonin (5-HT) produces a slow EPSP in Aplysia sensory neurons by closing the serotonin-sensitive or S-type K+ channels. The 5-HT receptor is coupled to Gs, which stimulates adenylyl cyclase. The increase in cAMP activates cAMP-dependent protein kinase A (PKA), which phosphorylates the S-type channel, leading to its closure. Single-channel recordings illustrate the actions of 5-HT, CAMP, and PKA on the S-type channels.
شکل 14-11 نورونهای سروتونرژیک یک کانال +K را از طریق cAMP پیام رسان دوم قابل انتشار میبندند. سروتونین (5-HT) با بستن کانالهای +K حساس به سروتونین یا نوع S، یک EPSP کند در نورونهای حسی Aplysia تولید میکند. گیرنده 5-HT با Gs جفت میشود که آدنیلیل سیکلاز را تحریک میکند. افزایش cAMP پروتئین کیناز A وابسته به cAMP (PKA) را فعال میکند که کانال نوع S را فسفریله میکند و منجر به بسته شدن آن میشود. ثبتهای تک کاناله، عملکرد 5HT ،CAMP و PKA را در کانالهای نوع S نشان میدهد.
A. Addition of 5-HT to the bath closes three of five S-type K+ channels active in this cell-attached patch of membrane. The experiment implicates a diffusible messenger, as the 5-HT applied in the bath has no direct access to the S-type channels in the membrane under the pipette. Each channel opening contributes an outward (positive) current pulse. (Adapted, with permission, from Siegelbaum, Camardo, and Kandel 1982.)
الف. افزودن 5-HT به حمام، سه کانال از پنج کانال +K نوع S فعال در این تکه غشایی متصل به سلول را میبندد. این آزمایش شامل یک پیام رسان قابل انتشار است، زیرا 5-HT اعمال شده در حمام هیچ دسترسی مستقیمی به کانالهای نوع S در غشای زیر پیپت ندارد. هر باز شدن کانال یک پالس جریان رو به بیرون (مثبت) ایجاد میکند. (اقتباس شده، با اجازه، از Siegelbaum، Camardo، and Kandel 1982.)
B. Injection of cAMP into a sensory neuron through a microelectrode closes all three active S-type channels in this patch. The bottom trace shows the closure of the final active channel in the presence of cAMP. (Adapted, with permission, from Siegelbaum, Camardo, and Kandel 1982.)
ب- تزریق cAMP به یک نورون حسی از طریق یک میکروالکترود، هر سه کانال فعال نوع S را در این پچ میبندد. ردپای پایین بسته شدن کانال فعال نهایی را در حضور cAMP نشان میدهد. (اقتباس شده، با اجازه، از Siegelbaum، Camardo، and Kandel 1982.)
C. Application of the purified catalytic subunit of PKA to the cytoplasmic surface of the membrane closes two out of four active S-type K+ channels in this cell-free patch. ATP was added to the solution bathing the inside surface of the membrane to provide the source of phosphate for protein phosphorylation. (Adapted, with permission, from Shuster et al. 1985.)
ج. استفاده از زیرواحد کاتالیزوری خالص شده PKA روی سطح سیتوپلاسمیغشاء، دو کانال از چهار کانال +K نوع S فعال را در این پچ بدون سلول میبندد. ATP به محلول حمام کردن سطح داخلی غشا اضافه شد تا منبع فسفات برای فسفوریلاسیون پروتئین فراهم شود. (اقتباس شده، با اجازه، از شوستر و همکاران 1985.)

Figure 14-12 A single neurotransmitter can have either short-term or long-term effects on an ion channel. In this example, a short exposure to transmitter activates the cAMP second-messenger system (1), which in turn activates PKA (2). The kinase phosphorylates a K+ channel; this leads to a synaptic potential that lasts for several minutes and modifies the excitability of the neuron (3). With sustained activation of the receptor, the kinase translocates to the nucleus, where it phosphorylates one or more transcription factors that turn on gene expression (4). As a result of the new protein synthesis, the synaptic actions are prolonged-closure of the channel and changes in neuronal excitability last days or longer (5). (Pol, polymerase.)
شکل 14-12 یک انتقال دهنده عصبی منفرد میتواند اثرات کوتاه مدت یا طولانی مدت بر روی یک کانال یونی داشته باشد. در این مثال، یک قرار گرفتن کوتاه در معرض ترانسمیتر، سیستم پیام رسان دوم cAMP (1) را فعال میکند، که به نوبه خود PKA (2) را فعال میکند. کیناز یک کانال +K را فسفریله میکند. این منجر به یک پتانسیل سیناپسی میشود که چند دقیقه طول میکشد و تحریک پذیری نورون را تغییر میدهد (3). با فعال شدن مداوم گیرنده، کیناز به هسته منتقل میشود، جایی که یک یا چند فاکتور رونویسی را فسفریله میکند که بیان ژن را فعال میکند (4). در نتیجه سنتز پروتئین جدید، اعمال سیناپسی به صورت طولانی مدت بسته شدن کانال و تغییرات در تحریک پذیری عصبی در روزهای آخر یا بیشتر است (5). (پل، پلیمراز.)
1. Modulatory projection neurons or neurohormones can coordinately influence the properties of large numbers of neurons to change the state of a neural circuit or of the entire animal. For example, modulators released from a relatively small number of neurons are important in the control of the transitions between sleep and wakefulness (Chapter 44).
1. نورونها یا هورمونهای عصبی تعدیلی میتوانند به طور هماهنگ بر خواص تعداد زیادی نورون برای تغییر وضعیت یک مدار عصبی یا کل حیوان تأثیر بگذارند. برای مثال، تعدیلکنندههای آزاد شده از تعداد نسبتاً کمی از نورونها در کنترل انتقال بین خواب و بیداری مهم هستند (فصل 44).
2. Neuromodulators act over intermediate time scales, ranging from many milliseconds to hours. Fast synaptic transmission and rapid action potential propagation are well suited for rapid computation of all kinds of processes important for behavior. Nevertheless, modulators that act over longer time scales can bias a circuit’s dynamics to expand its dynamic range or to adapt it to the behavioral needs of the animal. For example, many sensory processes will evoke very different responses depending on the behavioral state of the animal, and modulators that alter synaptic strength and intrinsic excitability are often involved in such actions.
2. تعدیل کنندههای عصبی در مقیاسهای زمانی متوسط، از چندین میلی ثانیه تا چند ساعت، عمل میکنند. انتقال سریع سیناپسی و انتشار پتانسیل عمل سریع برای محاسبه سریع انواع فرآیندهای مهم برای رفتار مناسب هستند. با این وجود، تعدیلکنندههایی که در مقیاسهای زمانی طولانیتر عمل میکنند، میتوانند دینامیک مدار را برای گسترش دامنه دینامیکی آن یا تطبیق آن با نیازهای رفتاری حیوان سوگیری کنند. به عنوان مثال، بسیاری از فرآیندهای حسی بسته به وضعیت رفتاری حیوان، پاسخهای بسیار متفاوتی را برمیانگیزند و تعدیل کنندههایی که قدرت سیناپسی و تحریک پذیری ذاتی را تغییر میدهند، اغلب در چنین اعمالی دخالت دارند.
Multiple Neuromodulators Can Converge Onto the Same Neuron and Ion Channels
تعدیل کنندههای عصبی متعدد میتوانند به کانالهای عصبی و یونی مشابهی همگرا شوند
We have seen in our discussion of the Aplysia S-channel how the same ion channel can be regulated by different modulatory agents. This is a common theme, as the M-type K+ channel is modulated by acetylcholine, substance P, and a variety of other peptides.
ما در بحث خود در مورد کانال S آپلزیا (Aplysia) دیدیم که چگونه کانال یونی یکسان میتواند توسط عوامل تعدیل کننده مختلف تنظیم شود. این یک موضوع رایج است، زیرا کانال +K نوع M توسط استیل کولین، ماده P و انواع دیگر پپتیدها تعدیل میشود.
One particularly striking example of convergence is seen in the modulatory control of the neurons of the crustacean stomatogastric ganglion. There, a large number of structurally diverse neuropeptides converge to modulate a voltage-dependent inward current (IMI). Although IMI is a small current, it plays an important role in regulating excitability and the generation of plateau and burst potentials. Many neurons express a large number of different receptor types, giving these cells the ability to respond flexibly to different modulatory inputs during different brain states.
یکی از نمونههای بارز همگرایی در کنترل تعدیلی نورونهای گانگلیون معده روزنهای سخت پوست دیده میشود. در آنجا، تعداد زیادی از نوروپپتیدهای ساختاری متنوع برای تعدیل یک جریان داخلی وابسته به ولتاژ (IMI) همگرا میشوند. اگرچه IMI یک جریان کوچک است، اما نقش مهمیدر تنظیم تحریک پذیری و تولید پتانسیلهای فلات و انفجار ایفا میکند. بسیاری از نورونها تعداد زیادی از انواع مختلف گیرنده را بیان میکنند و به این سلولها توانایی پاسخ دادن انعطافپذیر به ورودیهای تعدیلکننده مختلف در طول حالتهای مختلف مغز را میدهند.
The crustacean stomatogastric ganglion (STG) contains 26 to 30 neurons and generates two rhythmic motor patterns important for feeding the gastric rhythm and the pyloric rhythm. One set of STG neurons generates the pyloric rhythm, which is important for filtering food and is continuously active throughout the animal’s life. Another set of neurons generates the gastric mill rhythm, which moves three teeth inside of the stomach that are used to chew and grind food. The gastric mill rhythm is activated in response to food and is therefore only intermittently active in vivo. Whether a particular rhythm is active at any time is under the control of a variety of neuromodulators, some of which activate the pyloric and gastric mill rhythms, while others inhibit them. These modulators can be released at specific synaptic contacts or can act diffusely as neurohormones. Interestingly, modulators can also cause individual neurons to switch between these two circuits, thereby increasing the computational power that this small number of neurons can achieve.
گانگلیون معدی-دهانی سخت پوستان (STG) حاوی 26 تا 30 نورون است و دو الگوی حرکتی ریتمیک برای تغذیه ریتم معده و ریتم پیلور ایجاد میکند. یک مجموعه از نورونهای STG ریتم پیلور را تولید میکند که برای فیلتر کردن غذا مهم است و به طور مداوم در طول زندگی حیوان فعال است. مجموعه دیگری از نورونها ریتم آسیاب معده را تولید میکنند که سه دندان را در داخل معده حرکت میدهد که برای جویدن و آسیاب کردن غذا استفاده میشوند. ریتم آسیاب معده در پاسخ به غذا فعال میشود و بنابراین فقط به طور متناوب در داخل بدن فعال است. اینکه آیا یک ریتم خاص در هر زمان فعال است یا نه، تحت کنترل انواع تعدیل کنندههای عصبی است، که برخی از آنها ریتم پیلور و آسیاب معده را فعال میکنند، در حالی که برخی دیگر آنها را مهار میکنند. این تعدیلکنندهها میتوانند در تماسهای سیناپسی خاص آزاد شوند یا میتوانند بهعنوان هورمونهای عصبی منتشر شوند. جالب توجه است، تعدیلکنندهها همچنین میتوانند باعث شوند که نورونهای منفرد بین این دو مدار سوئیچ کنند و در نتیجه قدرت محاسباتی که این تعداد کمیاز نورونها میتوانند به دست آورند را افزایش دهند.
The fundamental circuit (the kernel) that serves as the pacemaker of the STG pyloric rhythm consists of a single anterior burster (AB) neuron and two pyloric dilator (PD) neurons. Both types of neurons make inhibitory synaptic connections with a third type of neuron, the pyloric (PY) neuron. During bursting, a slowly depolarizing pacemaker potential (slow wave) triggers a burst of action potentials in both AB and PD neurons. As these neurons are strongly coupled by electrical (gap-junction) synapses, they depolarize and synchronously fire bursts of action potentials, resulting in transient inhibition of the downstream PY neuron (Figure 14-13A).
مدار اصلی (هسته) که به عنوان ضربان ساز ریتم پیلور STG عمل میکند از یک نورون منفرد انفجاری قدامی (AB) و دو نورون گشادکننده پیلور (PD) تشکیل شده است. هر دو نوع نورون، اتصالات سیناپسی بازدارنده ای را با نوع سوم نورون، نورون پیلور (PY) ایجاد میکنند. در طول انفجار، یک پتانسیل ضربان ساز به آرامیدپلاریزاسیون (موج آهسته) باعث انفجار پتانسیلهای عمل در هر دو نورون AB و PD میشود. از آنجایی که این نورونها بهشدت توسط سیناپسهای الکتریکی (شکاف-اتصال) جفت میشوند، آنها دپلاریزه میشوند و به طور همزمان پتانسیلهای عمل را شلیک میکنند و در نتیجه باعث مهار موقت نورون PY پاییندست میشوند (شکل 14-13A).
Dopamine, which functions both as a fast neurotransmitter and as a neurohormone in crustaceans, influences feeding behavior by acting on many neurons and synapses to influence synaptic strength and neuronal and muscle excitability. For example, application of dopamine decreases the slow-wave amplitude in the PD neurons but increases the amplitude of the slow wave in the AB neurons. Ron Harris-Warrick found that dopamine modulates different sets of membrane currents in the two neurons, providing a clear example of how a single modulatory transmitter can exert distinct actions in different postsynaptic cells (Figure 14-13B).
دوپامین که هم به عنوان یک انتقال دهنده عصبی سریع و هم به عنوان یک هورمون عصبی در سخت پوستان عمل میکند، با تأثیر بر روی بسیاری از نورونها و سیناپسها برای تأثیرگذاری بر قدرت سیناپسی و تحریک پذیری عصبی و عضلانی، بر رفتار تغذیه تأثیر میگذارد. به عنوان مثال، استفاده از دوپامین دامنه موج آهسته را در نورونهای PD کاهش میدهد اما دامنه موج آهسته را در نورونهای AB افزایش میدهد. ران هریس-واریک دریافت که دوپامین مجموعههای متفاوتی از جریانهای غشایی را در دو نورون تعدیل میکند، و نمونهای واضح از این که چگونه یک ترانسمیتر تعدیلی میتواند اعمال متمایزی را در سلولهای پسسیناپسی مختلف انجام دهد ارائه میکند (شکل 14-13B).
Dopamine also alters the relative timing of the activity of these neurons. Although the PY neuron receives inhibitory input from both the AB and PD neurons, the inhibitory synaptic action from the AB neuron is faster than that from the PD neuron. Thus, dopamine, by inhibiting the PD neuron and suppressing the slow component of the IPSP, acts to speed the time course of the combined IPSP in the PY neurons (Figure 14-13A), contributing to a change in the timing of the activity of the PY neurons relative to that of the pacemaker group. Dopamine also enhances firing in the PY neuron by modulating its intrinsic excitability, by decreasing the transient A-type K+ current (IK,A) while increasing the excitatory slow inward current carried by the HCN channels (Ih) (Figure 14-13B). Thus, the effects of a modulator on the circuit result from its selective actions on a number of voltage-gated channels and synapses in distributed circuit elements.
دوپامین همچنین زمان نسبی فعالیت این نورونها را تغییر میدهد. اگرچه نورون PY ورودی مهاری را از هر دو نورون AB و PD دریافت میکند، عملکرد سیناپسی بازدارنده از نورون AB سریعتر از نورون PD است. بنابراین، دوپامین، با مهار نورون PD و سرکوب مؤلفه آهسته IPSP، برای سرعت بخشیدن به دوره زمانی IPSP ترکیبی در نورونهای PY عمل میکند (شکل 14-13A)، که به تغییر در زمانبندی فعالیت کمک میکند. نورونهای PY نسبت به نورونهای گروه ضربان ساز. دوپامین همچنین با تعدیل تحریک پذیری ذاتی آن، با کاهش جریان +K نوع A گذرا (IK,A) و در عین حال افزایش جریان آهسته تحریکی به سمت داخل که توسط کانالهای HCN (Ih) حمل میشود، شلیک را در نورون PY افزایش میدهد (شکل 14-13B). بنابراین، اثرات یک مدولاتور بر روی مدار ناشی از اعمال انتخابی آن بر روی تعدادی از کانالها و سیناپسهای دارای ولتاژ در عناصر مدار توزیع شده است.
Why So Many Modulators?
چرا تعدیل کنندههای زیادی وجود دارد؟
We now know that the STG is the direct target of 50 or more different neuromodulatory substances, including biogenic amines, amino acids, NO, and a host of neuropeptides that are released from descending modulatory projection neurons and sensory neurons and that circulate as hormones in the hemolymph. Many of these modulators are released as cotransmitters from the terminals of certain descending fibers that are activated by sensory neurons. Many neuromodulators are both released synaptically in the STG neuropil and also function as neurohormones.
ما اکنون میدانیم که STG هدف مستقیم 50 یا بیشتر ماده تعدیل کننده عصبی مختلف است، از جمله آمینهای بیوژنیک، اسیدهای آمینه، NO، و تعدادی نوروپپتید که از نورونهای پیش بینی تعدیلی نزولی و نورونهای حسی آزاد میشوند و به عنوان هورمون در گردش میشوند. همولنف بسیاری از این تعدیلکنندهها به عنوان انتقالدهندههای همزمان از پایانههای فیبرهای نزولی خاصی که توسط نورونهای حسی فعال میشوند، آزاد میشوند. بسیاری از تعدیل کنندههای عصبی هم به صورت سیناپسی در نوروپیل STG آزاد میشوند و هم به عنوان هورمون عصبی عمل میکنند.
Why should a small ganglion composed of only 26 to 30 neurons be modulated by so many substances? At first, it was thought that the richness of the modulatory innervation was important for producing different behaviorally relevant motor outputs. This remains true, but it is now also evident that some modulators may be used exclusively in special circumstances, such as molting, and that different modulators with similar effects ensure that important functions are preserved even if one modulatory system is lost. Thus, diverse modulators may be used in the service of both plasticity and stability.
چرا یک گانگلیون کوچک که فقط از 26 تا 30 نورون تشکیل شده است باید توسط این همه مواد تعدیل شود؟ در ابتدا، تصور میشد که غنای عصب تعدیلی برای تولید خروجیهای حرکتی مرتبط با رفتار متفاوت مهم است. این درست است، اما در حال حاضر نیز مشهود است که برخی از تعدیلکنندهها ممکن است منحصراً در شرایط خاص مانند پوستاندازی استفاده شوند و تعدیلکنندههای مختلف با اثرات مشابه تضمین میکنند که عملکردهای مهم حفظ میشوند حتی اگر یک سیستم تعدیلی از بین برود. بنابراین، تعدیل کنندههای متنوعی ممکن است در خدمت انعطاف پذیری و پایداری استفاده شوند.


Figure 14-13 The modulatory action of dopamine on the pyloric rhythm of the lobster stomatogastric ganglion results from numerous actions.
شکل 14-13 اثر تعدیلی دوپامین بر ریتم پیلور گانگلیون معدی-دهانی خرچنگ ناشی از اعمال متعدد است.
A. A circuit diagram shows the interactions between three of the pyloric circuit neurons. The anterior burster (AB) and pyloric dilator (PD) neurons are strongly electrically coupled by gap-junction channels. Both the AB and PD neurons form inhibitory synapses with the pyloric (PY) neuron that generate inhibitory postsynaptic potentials (IPSPs) in this cell. Intracellular voltage recordings illustrate phases of pyloric rhythm from PD, AB, and PY neurons without dopaminergic input (control) and with dopamine. On the right, the voltage traces from control cells (C) and cells with dopaminergic input (DA) are overlaid. Dopamine enhances the amplitude of the slow-wave burst in the AB neuron (in this neuron, axonal action potentials are highly attenuated by the cable properties of the neuron and appear in the soma as faint ripples) but hyperpolarizes and decreases the amplitude of the slow wave in the PD neurons. These combined actions result in a shorter IPSP in the PY neuron, enabling it to fire earlier relative to the PD neurons. (Adapted, with permission, from Eisen and Marder 1984.)
الف. نمودار مداری برهمکنش بین سه نورون مدار پیلور را نشان میدهد. نورونهای برستر قدامی (AB) و گشادکننده پیلور (PD) به شدت توسط کانالهای اتصال شکاف جفت میشوند. هر دو نورون AB و PD سیناپسهای مهاری را با نورون پیلور (PY) تشکیل میدهند که پتانسیلهای پسسیناپسی مهاری (IPSPs) را در این سلول ایجاد میکند. ثبتهای ولتاژ داخل سلولی فازهای ریتم پیلور را از نورونهای PD، AB و PY بدون ورودی دوپامینرژیک (کنترل) و با دوپامین نشان میدهند. در سمت راست، ردیابی ولتاژ از سلولهای کنترل (C) و سلولهای با ورودی دوپامینرژیک (DA) روی هم قرار گرفتهاند. دوپامین دامنه انفجار موج آهسته را در نورون AB افزایش میدهد (در این نورون، پتانسیلهای عمل آکسونی به دلیل ویژگیهای کابلی نورون بسیار ضعیف میشوند و در سوما به صورت موجهای ضعیف ظاهر میشوند) اما هیپرپلاریزه میشود و دامنه آهسته را کاهش میدهد. موج در نورونهای PD این اقدامات ترکیبی منجر به IPSP کوتاهتر در نورون PY میشود و آن را قادر میسازد نسبت به نورونهای PD زودتر شلیک کند. (اقتباس شده، با اجازه، از آیزن و ماردر 1984.)
B. Dopamine modulates a number of different voltage-dependent channels in the AB, PD, and PY neurons. These include Ca2+ currents (Ica), a calcium-activated K+ current (Ik.ca), an inactivating K+ current (IK,A), a delayed rectifier K+ current (Ikv), the hyperpolarization-activated cation current (Ih), and a persistent Na+ current (INa). Lines with arrowheads indicate current increase, lines ending in short line segment indicate current decrease. (Adapted, with permission, from Marder and Bucher 2007. For effects of dopamine on the complete pyloric circuit, see Harris-Warrick, 2011.)
ب. دوپامین تعدادی از کانالهای مختلف وابسته به ولتاژ را در نورونهای AB، PD و PY تعدیل میکند. اینها شامل جریانهای Ca2+ (Ica)، یک جریان +K فعال شده با کلسیم (Ik.ca)، یک جریان +K غیرفعال (IK,A)، یک جریان +K یکسوکننده تأخیری (Ikv)، جریان کاتیون فعال شده با هیپرپلاریزاسیون (Ih)، و یک جریان +Na پایدار (INa). خطوط با نوک پیکان نشان دهنده افزایش جریان و خطوطی که به پاره خط کوتاه ختم میشوند نشان دهنده کاهش جریان هستند. (اقتباس شده، با اجازه، از Marder و Bucher 2007. برای اثرات دوپامین بر مدار کامل پیلور، به Harris-Warrick، 2011 مراجعه کنید.)
Highlights
نکات برجسته
1. Neuromodulators are substances that bind to receptors, most of which are metabotropic, to alter the excitability of neurons, the likelihood of transmitter release, or the functional state of receptors on postsynaptic neurons.
1. تعدیل کنندههای عصبی موادی هستند که به گیرندههایی که بیشتر آنها متابوتروپیک هستند، متصل میشوند تا تحریک پذیری نورونها، احتمال آزاد شدن ترانسمیتر یا وضعیت عملکردی گیرندههای نورونهای پسسیناپسی را تغییر دهند.
2. When neuromodulators activate second- messenger pathways, the modulator can influence the properties of ion channels and other targets at some distance from the site of release.
2. هنگامی که تعدیل کنندههای عصبی مسیرهای پیام رسان دوم را فعال میکنند، تعدیل کننده میتواند بر خواص کانالهای یونی و سایر اهداف در فاصله ای از محل انتشار تأثیر بگذارد.
3. Some neuromodulatory systems have wide- spread and pronounced actions over many neurons and many brain areas.
3. برخی از سیستمهای تعدیل کننده عصبی دارای اعمال گسترده و مشخص بر روی بسیاری از نورونها و بسیاری از مناطق مغز هستند.
4. There are two major families of metabotropic receptors: G protein-coupled receptors and receptor tyrosine kinases. Many important brain signaling molecules, such as norepinephrine, ACh, GABA, glutamate, serotonin, dopamine, and many diverse neuropeptides, activate metabotropic receptors; many of these same substances also activate ionotropic receptors.
4. دو خانواده اصلی از گیرندههای متابوتروپیک وجود دارد: گیرندههای جفت شده با پروتئین G و گیرنده تیروزین کیناز. بسیاری از مولکولهای مهم سیگنال دهنده مغز، مانند نوراپی نفرین، ACh، GABA، گلوتامات، سروتونین، دوپامین، و بسیاری از نوروپپتیدهای متنوع، گیرندههای متابوتروپیک را فعال میکنند. بسیاری از همین مواد گیرندههای یونوتروپیک را نیز فعال میکنند.
5. The cyclic AMP pathway is among the best- understood second-messenger signaling cascades. Metabotropic receptor activation triggers a sequence of biochemical reactions that result in activation of adenylyl cyclase, which synthesizes CAMP, which in turn activates protein kinase A. The kinase then phosphorylates target proteins, altering their functional state. Important targets for PKA include voltage- and ligand-gated ion channels as well as proteins important in vesicle release.
5. مسیر AMP حلقوی یکی از بهترین آبشارهای سیگنال دهی پیام رسان دوم است. فعالسازی گیرنده متابوتروپیک، دنبالهای از واکنشهای بیوشیمیایی را تحریک میکند که منجر به فعال شدن آدنیلیل سیکلاز میشود، که cAMP را سنتز میکند، که به نوبه خود پروتئین کیناز A را فعال میکند. سپس کیناز پروتئینهای هدف را فسفریله میکند و حالت عملکردی آنها را تغییر میدهد. اهداف مهم برای PKA شامل کانالهای یونی دریچهدار وابسته به ولتاژ و وابسته به لیگاند و همچنین پروتئینهای مهم در آزادسازی وزیکول میباشد.
6. Hydrolysis of phospholipids by phospholipase C produces DAG and IP3, which plays an important role in intracellular Ca2+ handling. Endocannabinoids are synthesized from lipid precursors and can act across synapses as retrograde messengers. Another generalized signaling molecule is the gas nitric oxide, which diffuses across membranes and stimulates cyclic GMP synthesis.
6. هیدرولیز فسفولیپیدها توسط فسفولیپاز C باعث تولید DAG و IP3 میشود که نقش مهمیدر جابجایی +Ca2 داخل سلولی ایفا میکند. اندوکانابینوئیدها از پیش سازهای لیپیدی سنتز میشوند و میتوانند در سراسر سیناپسها به عنوان پیام رسانهای رتروگراد عمل کنند. یکی دیگر از مولکولهای سیگنالدهنده تعمیمیافته، گاز نیتریک اکسید است که در غشاها پخش میشود و سنتز GMP حلقوی را تحریک میکند.
7. The receptor tyrosine kinases also gate ion channels indirectly in response to binding a variety of peptide hormones.
7. گیرنده تیروزین کیناز همچنین کانالهای یونی را به طور غیر مستقیم در پاسخ به اتصال انواع هورمونهای پپتیدی دروازه میکند.
8. Neuromodulators can close ion channels, thus producing decreases in membrane conductance. The M-type current is a slowly activating voltage- gated K+ current that underlies action potential adaptation. ACh and several neuropeptides decrease M-type current amplitude, thereby producing a slow depolarization and decreasing adaptation. The S-type K+ channel contributes to the resting K+ conductance of certain neurons, including a class of sensory neurons mediating the Aplysia gill withdrawal reflex. Closure of the channel by serotonin, acting through a cAMP signaling cascade, depolarizes the resting membrane, increases excitability, and enhances transmitter release from sensory neuron terminals. Prolonged exposure to serotonin can alter gene transcription to produce long-term changes in synaptic strength.
8. نورومدولاتورها میتوانند کانالهای یونی را ببندند، بنابراین باعث کاهش رسانایی غشایی میشوند. جریان نوع M یک جریان +K با ولتاژ با آهسته فعال است که زمینه ساز انطباق پتانسیل عمل است. ACh و چندین نوروپپتید دامنه جریان نوع M را کاهش میدهند، در نتیجه دپلاریزاسیون آهسته ایجاد میکنند و سازگاری را کاهش میدهند. کانال +K نوع S به هدایت +K استراحت نورونهای خاص، از جمله یک کلاس از نورونهای حسی که واسطه رفلکس خروج آبشش حلزون دریایی Aplysia هستند، کمک میکند. بسته شدن کانال توسط سروتونین، که از طریق یک آبشار سیگنالینگ cAMP عمل میکند، غشای در حال استراحت را دپولاریزه میکند، تحریک پذیری را افزایش میدهد و آزادسازی ترانسمیتر از پایانههای عصبی حسی را افزایش میدهد. قرار گرفتن طولانیمدت در معرض سروتونین میتواند رونویسی ژن را برای ایجاد تغییرات طولانیمدت در قدرت سیناپسی تغییر دهد.
9. Modulators can alter the output of neuronal circuits by acting on numerous circuit targets.
9. تعدیل کنندهها میتوانند خروجی مدارهای عصبی را با عمل بر روی اهداف مداری متعدد تغییر دهند.
10. Given that all brain neurons and synapses are likely to be modulated by one or more substances, it is remarkable that brain circuits are only rarely “overmodulated” so that they lose their function. Much additional research is needed to understand the rules that allow robust and stable net- work performance in the face of the modulators that allow network plasticity.
10. با توجه به اینکه همه نورونها و سیناپسهای مغز احتمالاً توسط یک یا چند ماده تعدیل میشوند، قابل توجه است که مدارهای مغز به ندرت «بیش از حد تعدیل» میشوند تا عملکرد خود را از دست بدهند. تحقیقات بیشتری برای درک قوانینی که امکان عملکرد قوی و پایدار شبکه را در مواجهه با تعدیلکنندههایی که انعطافپذیری شبکه را امکانپذیر میکنند، مورد نیاز است.
11. Except in a few notable cases such as small ganglia or the retina, it is likely that we still have only a partial catalog of the total number of neuromodulatory substances that are present and active.
11. به جز در چند مورد قابل توجه مانند عقدههای کوچک یا شبکیه، این احتمال وجود دارد که ما هنوز فقط یک کاتالوگ جزئی از تعداد کل مواد تعدیل کننده عصبی موجود و فعال داشته باشیم.
12. Much of what we know about neuromodulatory actions comes from in vitro studies. Much less is known about how neuromodulatory concentrations are controlled in behaving animals.
12. بسیاری از آنچه که ما در مورد اعمال تعدیل کننده عصبی میدانیم از مطالعات آزمایشگاهی ناشی میشود. اطلاعات کمتری در مورد چگونگی کنترل غلظتهای عصبی در رفتار حیوانات وجود دارد.
Steven A. Siegelbaum
David E. Clapham
Eve Marder
کلیک کنید تا Selected Reading نمایش داده شود
Berridge MJ. 2016. The inositol trisphosphate/calcium signaling pathway in health and disease. Physiol Rev 96:1261-1296.
Greengard P. 2001. The neurobiology of slow synaptic trans- mission. Science 294:1024-1030.
Hille B, Dickson EJ, Kruse M, Vivas O, Suh BC. 2015. Phosphoinositides regulate ion channels. Biochim Biophys Acta 1851:844-856.
Kobilka B. 2013. The structural basis of G-protein-coupled receptor signaling (Nobel Lecture). Angew Chem Int Ed Engl 52:6380-6388.
Levitan IB. 1999. Modulation of ion channels by protein phosphorylation. How the brain works. Adv Second Mes- senger Phosphoprotein Res 33:3-22.
Lu HC, Mackie K. 2016. An introduction to the endogenous cannabinoid system. Biol Psychiatry 79:516-525.
Marder E. 2012. Neuromodulation of neuronal circuits: back to the future. Neuron 76:1-11.
Schwartz JH. 2001. The many dimensions of cAMP signaling. Proc Natl Acad Sci USA 98:13482-13484.
Syrovatkina V, Alegre KO, Dey R, Huang XY. 2016. Regulation, signaling, and physiological functions of G-proteins. J Mol Biol 428:3850-3868.
Takemoto-Kimura S, Suzuki K, Horigane SI, et al. 2017. Calmodulin kinases: essential regulators in health and disease. J Neurochem 141:808-818.
کلیک کنید تا References نمایش داده شود
Adams PR, Jones SW, Pennefather P, Brown DA, Koch C, Lancaster B. 1986. Slow synaptic transmission in frog sympathetic ganglia. J Exp Biol 124:259–285.
Alberts B, Bray D, Lewis J, Raff M, Roberts K, Watson JD. 1994. Molecular Biology of the Cell, 3rd ed. New York:
Garland.
Eisen JS, Marder E. 1984. A mechanism for the production of phase shifts in a pattern generator. J Neurophysiol 51:1375-1393.
Fantl WJ, Johnson DE, Williams LT. 1993. Signalling by receptor tyrosine kinases. Annu Rev Biochem 62:453-481.
Frielle T, Kobilka B, Dohlman H, Caron MG, Lefkowitz RJ. 1989. The ẞ-adrenergic receptor and other receptors coupled to guanine nucleotide regulatory proteins. In: S Chien (ed). Molecular Biology in Physiology, pp. 79-91. New York: Raven.
Halpain S, Girault JA, Greengard P. 1990. Activation of NMDA receptors induces dephosphorylation of DARPP-32
in rat striatal slices. Nature 343:369-372.
Harris-Warrick, RM. 2011. Neuromodulation and flexibility in central pattern generating networks. Curr Opin Neuro- biol 21:685-692.
Logothetis DE, Kurachi Y, Galper J, Neer EJ, Clapham DE. 1987. The By subunits of GTP-binding proteins activate the muscarinic K+ channel in heart. Nature 325:321-326.
Marder E, Bucher D. 2007. Understanding circuit dynamics using the stomatogastric nervous system of lobsters and crabs. Annu Rev Physiol 69:291-316.
Nusbaum MP, Blitz DM, Marder E. 2017. Functional consequences of neuropeptide/small molecule cotransmission. Nature Rev Neurosci 18:389–403.
Osten P, Valsamis L, Harris A, Sacktor TC. 1996. Protein synthesis-dependent formation of protein kinase Mzeta in long-term potentiation. J Neurosci 16:2444-2451.
Pfaffinger PJ, Martin JM, Hunter DD, Nathanson NM, Hille B. 1985. GTP-binding proteins couple cardiac muscarinic receptors to a K channel. Nature 317:536–538.
Phillis JW, Horrocks LA, Farooqui AA. 2006. Cyclooxygenases, lipoxygenases, and epoxygenases in CNS: their role and involvement in neurological disorders. Brain Res Rev 52:201-243.
Shuster MJ, Camardo JS, Siegelbaum SA, Kandel ER. 1985. Cyclic AMP-dependent protein kinase closes the serotonin- sensitive K+ channels of Aplysia sensory neurones in cell- free membrane patches. Nature 313:392-395.
Siegelbaum SA, Camardo JS, Kandel ER. 1982. Serotonin and cyclic AMP close single K+ channels in Aplysia sensory neurones. Nature 299:413-417.
Soejima M, Noma A. 1984. Mode of regulation of the ACh- sensitive K-channel by the muscarinic receptor in rabbit atrial cells. Pflugers Arch 400:424-431.
Tedford HW, Zamponi GW. 2006. Direct G protein modulation of Cav2 calcium channels. Pharmacol Rev 58: 837-862.
Toda N, West TC. 1967. Interactions of K, Na, and vagal stimulation in the S-A node of the rabbit. Am J Physiol 212:416-423.
Whorton MR, MacKinnon R. 2013. X-ray structure of the mammalian GIRK2-betagamma G-protein complex. Nature 498:190-197.
Zeng L, Webster SV, Newton PM. 2012. The biology of protein kinase C. Adv Exp Med Biol 740:639–661.
»» ابتدای فصل: قسمت اول فصل مدولاسیون انتقال سیناپسی و تحریک پذیری عصبی: پیام رسانهای دوم
»» فصل قبل: فصل سیزدهم: ادغام سیناپسی در سیستم عصبی مرکزی
»» فصل بعد: فصل پانزدهم: آزادسازی انتقالدهنده
»» تمامی کتاب

 ورود/ ثبت نام با جیمیل
ورود/ ثبت نام با جیمیل
