» Guyton and Hall Textbook of Medical Physiology, 14th Ed
»» CHAPTER 8
Excitation and Contraction of Smooth Muscle
CONTRACTION OF SMOOTH MUSCLE
Smooth muscle is composed of small fibers that are usu- ally 1 to 5 micrometers in diameter and only 20 to 500 micrometers in length. In contrast, skeletal muscle fibers are as much as 30 times greater in diameter and hundreds of times as long. Many of the same principles of contrac- tion apply to smooth muscle as to skeletal muscle. Most important, essentially the same attractive forces between myosin and actin filaments cause contraction in smooth muscle as in skeletal muscle, but the internal physical arrangement of smooth muscle fibers is different.
انقباض عضله صاف
ماهیچه صاف از فیبرهای کوچکی تشکیل شده است که معمولاً ۱ تا ۵ میکرومتر قطر و تنها ۲۰ تا ۵۰۰ میکرومتر طول دارند. در مقابل، فیبرهای عضلانی اسکلتی ۳۰ برابر قطر و صدها برابر طول دارند. بسیاری از اصول انقباض در مورد عضلات صاف مانند ماهیچههای اسکلتی اعمال میشود. مهمتر از همه، اساساً همان نیروهای جذاب بین رشتههای میوزین و اکتین باعث انقباض در ماهیچه صاف مانند ماهیچه اسکلتی میشود، اما آرایش فیزیکی داخلی رشتههای عضلانی صاف متفاوت است.
TYPES OF SMOOTH MUSCLE
The smooth muscle of each organ is distinctive from that of most other organs in several ways: (1) physical dimensions; (2) organization into bundles or sheets; (3) response to different types of stimuli; (4) characteristics of innervation; and (5) function. Yet, for the sake of sim- plicity, smooth muscle can generally be divided into two major types, which are shown in Figure 8-1, multi-unit smooth muscle and unitary (or single-unit) smooth muscle.
انواع ماهیچههای صاف
عضله صاف هر اندام از چند جهت متمایز از اکثر اندامهای دیگر است: (۱) ابعاد فیزیکی. (۲) سازماندهی به بستهها یا برگهها. (۳) پاسخ به انواع مختلف محرک. (۴) ویژگیهای عصب. و (۵) عملکرد. با این حال، برای سادگی، عضله صاف را میتوان به طور کلی به دو نوع عمده تقسیم کرد، که در شکل ۸-۱ نشان داده شده است، عضله صاف چند واحدی و عضله صاف واحد (یا تک واحدی).
Figure 8-1
Multi-unit (A) and unitary (B) smooth muscle.
شکل ۸-۱
عضله صاف چند واحدی (A) و واحدی (B).
Multi-Unit Smooth Muscle. Multi-unit smooth muscle is composed of discrete, separate, smooth muscle fibers. Each fiber operates independently of the others and often is innervated by a single nerve ending, as occurs for skele- tal muscle fibers. Furthermore, the outer surfaces of these fibers, like those of skeletal muscle fibers, are covered by a thin layer of basement membrane-like substance, a mix- ture of fine collagen and glycoprotein that helps insulate the separate fibers from one another.
ماهیچه صاف چند واحدی. عضله صاف چند واحدی از فیبرهای عضلانی مجزا، مجزا و صاف تشکیل شده است. هر فیبر مستقل از فیبرهای دیگر عمل میکند و اغلب توسط یک انتهای عصبی عصب دهی میشود، همانطور که برای فیبرهای عضلانی اسکلتی اتفاق میافتد. علاوه بر این، سطوح بیرونی این الیاف، مانند فیبرهای عضلانی اسکلتی، توسط یک لایه نازک از ماده غشای پایه مانند، مخلوطی از کلاژن ریز و گلیکوپروتئین پوشیده شده است که به عایق بندی الیاف جداگانه از یکدیگر کمک میکند.
Important characteristics of multi-unit smooth muscle fibers are that each fiber can contract indepen- dently of the others, and their control is exerted mainly by nerve signals. In contrast, a major share of control of unitary smooth muscle is exerted by non-nervous stimuli. Some examples of multi-unit smooth muscle are the ciliary muscle of the eye, the iris muscle of the eye, and the piloerector muscles that cause erection of the hairs when stimulated by the sympathetic nervous system.
ویژگیهای مهم فیبرهای عضلانی صاف چند واحدی این است که هر فیبر میتواند مستقل از بقیه منقبض شود و کنترل آنها عمدتاً توسط سیگنالهای عصبی اعمال میشود. در مقابل، سهم عمده ای از کنترل عضله صاف واحد توسط محرکهای غیر عصبی اعمال میشود. برخی از نمونههای ماهیچه صاف چند واحدی عبارتند از: عضله مژگانی چشم، عنبیه چشم و ماهیچههای پیلوئرکتور که با تحریک سیستم عصبی سمپاتیک باعث نعوظ موها میشوند.
Unitary Smooth Muscle. Unitary smooth muscle is also called syncytial smooth muscle or visceral smooth muscle. The term unitary does not mean single muscle fibers. Instead, it means a mass of hundreds to thousands of smooth muscle fibers that contract together as a single unit. The fibers usually are arranged in sheets or bundles, and their cell membranes are adherent to one another at multiple points so that force generated in one muscle fiber can be transmitted to the next. In addition, the cell membranes are joined by many gap junctions through which ions can flow freely from one muscle cell to the next so that ac- tion potentials, or ion flow without action potentials, can travel from one fiber to the next and cause the muscle fib- ers to contract together. This type of smooth muscle is also known as syncytial smooth muscle because of its syncyt- ial interconnections among fibers. It is also called visceral smooth muscle because it is found in the walls of most vis- cera of the body, including the gastrointestinal tract, bile ducts, ureters, uterus, and many blood vessels.
عضله صاف واحد. عضله صاف یکپارچه به عضله صاف سنسیشیال یا عضله صاف احشایی نیز گفته میشود. اصطلاح واحد به معنای فیبرهای عضلانی منفرد نیست. در عوض، به معنای تودهای متشکل از صدها تا هزاران فیبر عضلانی صاف است که به صورت یک واحد منقبض میشوند. فیبرها معمولاً در صفحات یا بستههایی قرار میگیرند و غشای سلولی آنها در چندین نقطه به یکدیگر میچسبند تا نیروی ایجاد شده در یک فیبر عضلانی به رشته بعدی منتقل شود. علاوه بر این، غشاهای سلولی با اتصالات شکاف زیادی به هم میپیوندند که از طریق آن یونها میتوانند آزادانه از یک سلول عضلانی به سلول دیگر جریان پیدا کنند، به طوری که پتانسیل عمل یا جریان یون بدون پتانسیل عمل، میتواند از یک فیبر به فیبر دیگر حرکت کند و باعث شود تا فیبرهای عضلانی با هم منقبض شوند. این نوع ماهیچه صاف به دلیل اتصالات متقابل سنسیتیال بین فیبرها به عضله صاف سنسیشیال نیز معروف است. به آن عضله صاف احشایی نیز میگویند زیرا در دیوارههای اکثر احشاء بدن، از جمله دستگاه گوارش، مجاری صفراوی، حالب، رحم و بسیاری از رگهای خونی یافت میشود.
CONTRACTILE MECHANISM IN SMOOTH MUSCLE
مکانیسم انقباض در عضله صاف
Chemical Basis for Smooth Muscle Contraction
Smooth muscle contains both actin and myosin filaments, having chemical characteristics similar to those of the actin and myosin filaments in skeletal muscle. It does not contain the troponin complex that is required for the con- trol of skeletal muscle contraction, and thus the mecha- nism for controlling contraction is different. This topic is discussed in more detail later in this chapter.
مبنای شیمیایی برای انقباض عضلات صاف
ماهیچه صاف دارای هر دو رشته اکتین و میوزین است که دارای ویژگیهای شیمیایی مشابه رشتههای اکتین و میوزین در ماهیچههای اسکلتی است. حاوی کمپلکس تروپونین نیست که برای کنترل انقباض ماهیچههای اسکلتی لازم است و بنابراین مکانیسم کنترل انقباض متفاوت است. این موضوع در ادامه این فصل با جزئیات بیشتر مورد بحث قرار میگیرد.
Chemical studies have shown that actin and myo- sin filaments derived from smooth muscle interact with each other in much the same way that they do in skeletal muscle. Furthermore, the contractile process is activated by calcium ions, and adenosine triphosphate (ATP) is degraded to adenosine diphosphate (ADP) to provide the energy for contraction.
مطالعات شیمیایی نشان دادهاند که رشتههای اکتین و میوسین مشتق شده از ماهیچههای صاف، به همان شیوهای که در ماهیچههای اسکلتی انجام میدهند، با یکدیگر تعامل دارند. علاوه بر این، فرآیند انقباض توسط یونهای کلسیم فعال میشود و آدنوزین تری فسفات (ATP) به آدنوزین دی فسفات (ADP) تجزیه میشود تا انرژی برای انقباض فراهم شود.
There are, however, major differences between the physical organization of smooth muscle and that of skeletal muscle, as well as differences in excitation- contraction coupling, control of the contractile process by calcium ions, duration of contraction, and the amount of energy required for contraction.
با این حال، تفاوتهای عمدهای بین سازمان فیزیکی ماهیچه صاف و ماهیچه اسکلتی وجود دارد، همچنین تفاوتهایی در جفت شدن تحریک-انقباض، کنترل فرآیند انقباض توسط یونهای کلسیم، مدت زمان انقباض و مقدار انرژی مورد نیاز برای انقباض وجود دارد.
Physical Basis for Smooth Muscle Contraction
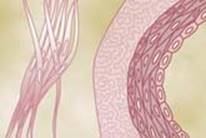
Smooth muscle does not have the same striated arrange- ment of actin and myosin filaments as is found in skeletal muscle. Instead, electron micrographic techniques sug- gest the physical organization shown in Figure 8-2, which illustrates large numbers of actin filaments attached to dense bodies. Some of these bodies are attached to the cell membrane, and others are dispersed inside the cell. Some of the membrane-dense bodies of adjacent cells are bonded together by intercellular protein bridges. It is mainly through these bonds that the force of contraction is transmitted from one cell to the next.
مبنای فیزیکی برای انقباض عضلات صاف
عضله صاف آرایش مخطط رشتههای اکتین و میوزین را ندارد که در ماهیچههای اسکلتی یافت میشود. در عوض، تکنیکهای میکروگرافیک الکترونی سازماندهی فیزیکی نشاندادهشده در شکل ۸-۲ را نشان میدهد که تعداد زیادی از رشتههای اکتین متصل به اجسام متراکم را نشان میدهد. برخی از این اجسام به غشای سلولی متصل میشوند و برخی دیگر در داخل سلول پراکنده میشوند. برخی از اجسام غشایی سلولهای مجاور توسط پلهای پروتئینی بین سلولی به یکدیگر متصل میشوند. عمدتاً از طریق این پیوندها است که نیروی انقباض از یک سلول به سلول دیگر منتقل میشود.
Interspersed among the actin filaments in the muscle fiber are myosin filaments. These filaments have a diam- eter more than twice that of the actin filaments. In elec- tron micrographs, 5 to 10 times as many actin filaments as myosin filaments are usually found.
در میان رشتههای اکتین در فیبر عضلانی رشتههای میوزین قرار دارند. این رشتهها دارای قطری بیش از دو برابر رشتههای اکتین هستند. در میکروگرافهای الکترونی، معمولاً ۵ تا ۱۰ برابر رشتههای اکتینی نسبت به رشتههای میوزین یافت میشود.
To the right in Figure 8-2 is a postulated structure of an individual contractile unit in a smooth muscle cell, show- ing large numbers of actin filaments radiating from two dense bodies; the ends of these filaments overlap a myosin filament located midway between the dense bodies. This contractile unit is similar to the contractile unit of skeletal muscle, but without the regularity of the skeletal muscle structure. In fact, the dense bodies of smooth muscle serve the same role as the Z disks in skeletal muscle.
در سمت راست در شکل ۸-۲ یک ساختار فرضی از یک واحد انقباضی منفرد در یک سلول ماهیچه صاف است که تعداد زیادی رشته اکتین را نشان میدهد که از دو جسم متراکم تابش میکنند. انتهای این رشتهها بر روی یک رشته میوزین که در میانه راه بین اجسام متراکم قرار دارد همپوشانی دارند. این واحد انقباضی شبیه واحد انقباضی عضله اسکلتی است، اما بدون منظم بودن ساختار عضله اسکلتی. در واقع، اجسام متراکم عضله صاف همان نقش دیسکهای Z را در عضله اسکلتی ایفا میکنند.
Another difference is that most of the myosin filaments have “side polar” cross-bridges arranged so that the bridges on one side hinge in one direction, and those on the other side hinge in the opposite direction. This configuration allows the myosin to pull an actin filament in one direction on one side while simultaneously pulling another actin fila- ment in the opposite direction on the other side. The value of this organization is that it allows smooth muscle cells to contract as much as 80% of their length instead of being limited to less than 30%, as occurs in skeletal muscle.
تفاوت دیگر این است که بیشتر رشتههای میوزین دارای پلهای متقاطع «قطبی جانبی» هستند که بهگونهای مرتب شدهاند که پلهای یک طرف در یک جهت و پلهای طرف دیگر در جهت مخالف قرار میگیرند. این پیکربندی به میوزین اجازه میدهد تا یک رشته اکتین را در یک جهت در یک طرف بکشد در حالی که همزمان یک رشته اکتین دیگر را در جهت مخالف در سمت دیگر بکشد. ارزش این سازمان در این است که به سلولهای ماهیچه صاف اجازه میدهد به جای محدود شدن به کمتر از ۳۰ درصد، همانطور که در ماهیچههای اسکلتی اتفاق میافتد، تا ۸۰ درصد طولشان منقبض شوند.
Figure 8-2
Physical structure of smooth muscle. The fiber on the up- per left shows actin filaments radiating from dense bodies. The fiber on the lower left and at right demonstrate the relation of myosin filaments to actin filaments.
شکل ۸-۲
ساختار فیزیکی عضلات صاف فیبر در سمت چپ بالا رشتههای اکتین را نشان میدهد که از اجسام متراکم تابش میکنند. فیبر در سمت چپ پایین و در سمت راست رابطه رشتههای میوزین را با رشتههای اکتین نشان میدهد.
Comparison of Smooth Muscle Contraction and Skeletal Muscle Contraction
Although most skeletal muscles contract and relax rap- idly, most smooth muscle contraction is prolonged tonic contraction, sometimes lasting hours or even days. Therefore, it is to be expected that both the physical and chemical characteristics of smooth muscle versus skeletal muscle contraction would differ. Some of the differences are noted in the following sections.
مقایسه انقباض عضلات صاف و انقباض عضلات اسکلتی
اگرچه اکثر عضلات اسکلتی به سرعت منقبض و شل میشوند، اکثر انقباضات ماهیچه صاف انقباض طولانی مدت تونیک است که گاهی چند ساعت یا حتی چند روز طول میکشد. بنابراین، انتظار میرود که هر دو ویژگی فیزیکی و شیمیایی عضله صاف در مقابل انقباض عضلات اسکلتی متفاوت باشد. برخی از تفاوتها در بخشهای بعدی ذکر شده است.
Slow Cycling of the Myosin Cross-Bridges. The rapid- ity of cycling of the myosin cross-bridges in smooth mus- cle—that is, their attachment to actin, then release from the actin, and reattachment for the next cycle–is much slower than in skeletal muscle. The frequency is as little as 1/10 to 1/300 that in skeletal muscle. Yet, the fraction of time that the cross-bridges remain attached to the ac- tin filaments, which is a major factor that determines the force of contraction, is believed to be greatly increased in smooth muscle. A possible reason for the slow cycling is that the cross-bridge heads have far less ATPase activ- ity than in skeletal muscle; thus, degradation of the ATP that energizes the movements of the cross-bridge heads is greatly reduced, with corresponding slowing of the rate of cycling.
دوچرخهسواری آهسته پلهای متقاطع میوزین. سرعت چرخش پلهای متقاطع میوزین در ماهیچههای صاف – یعنی اتصال آنها به اکتین، سپس آزاد شدن از اکتین و اتصال مجدد برای چرخه بعدی – بسیار کندتر از عضله اسکلتی است. فرکانس آن به اندازه ۱/۱۰ تا ۱/۳۰۰ در عضله اسکلتی است. با این حال، اعتقاد بر این است که کسری از زمانی که پلهای متقاطع به رشتههای اکتین متصل میشوند، که عامل اصلی تعیین کننده نیروی انقباض است، در عضله صاف به میزان زیادی افزایش مییابد. یک دلیل احتمالی برای دوچرخهسواری آهسته این است که سرهای پل متقاطع نسبت به ماهیچههای اسکلتی فعالیت ATPase بسیار کمتری دارند. بنابراین، تخریب ATP که به حرکات سر پلهای متقاطع انرژی میدهد تا حد زیادی کاهش مییابد، با کاهش متناظر سرعت دوچرخهسواری.
Low Energy Requirement to Sustain Smooth Muscle Contraction. Only 1/10 to 1/300 as much energy is required to sustain the same tension of contraction in smooth muscle as in skeletal muscle. This, too, is believed to result from the slow attachment and detachment cycling of the cross-bridges, and because only one molecule of ATP is required for each cycle, regardless of its duration.
نیاز به انرژی کم برای حفظ انقباض عضلات صاف. فقط ۱/۱۰ تا ۱/۳۰۰ انرژی برای حفظ همان تنش انقباض در عضله صاف مانند عضلات اسکلتی مورد نیاز است. اعتقاد بر این است که این نیز از اتصال آهسته و چرخه جداشدگی پلهای متقاطع ناشی میشود، و به این دلیل که برای هر چرخه، صرف نظر از مدت زمان، تنها یک مولکول ATP مورد نیاز است.
This low energy utilization by smooth muscle is impor- tant to the overall energy economy of the body because organs such as the intestines, urinary bladder, gallbladder, and other viscera often maintain tonic muscle contraction almost indefinitely.
این مصرف کم انرژی توسط عضلات صاف برای صرفه جویی انرژی کلی بدن مهم است زیرا اندامهایی مانند روده، مثانه، کیسه صفرا و سایر احشاء اغلب انقباض عضلات مقوی را تقریباً به طور نامحدود حفظ میکنند.
Slowness of Onset of Contraction and Relaxation of the Total Smooth Muscle Tissue. A typical smooth muscle tissue begins to contract 50 to 100 milliseconds after it is excited, reaches full contraction about 0.5 sec- ond later, and then declines in contractile force in another 1 to 2 seconds, giving a total contraction time of 1 to 3 seconds. This is about 30 times as long as a single contrac- tion of an average skeletal muscle fiber. However, because there are so many types of smooth muscle, contraction of some types can be as short as 0.2 second or as long as 30 seconds.
کندی شروع انقباض و شل شدن بافت ماهیچه صاف کل. یک بافت عضله صاف معمولی ۵۰ تا ۱۰۰ میلی ثانیه پس از برانگیختگی شروع به انقباض میکند، حدود ۰.۵ ثانیه بعد به انقباض کامل میرسد و سپس در ۱ تا ۲ ثانیه دیگر از نیروی انقباضی کاسته میشود و زمان کلی انقباض را بین ۱ تا ۳ ثانیه میدهد. این حدود ۳۰ برابر طول یک انقباض یک فیبر عضلانی اسکلتی متوسط است. با این حال، از آنجایی که انواع زیادی از ماهیچههای صاف وجود دارد، انقباض برخی از انواع آن میتواند ۰.۲ ثانیه یا ۳۰ ثانیه طول بکشد.
The slow onset of contraction of smooth muscle, as well as its prolonged contraction, is caused by the slow- ness of attachment and detachment of the cross-bridges with the actin filaments. In addition, the initiation of con- traction in response to calcium ions is much slower than in skeletal muscle, as will be discussed later.
شروع آهسته انقباض ماهیچه صاف و همچنین انقباض طولانی مدت آن به دلیل کندی اتصال و جدا شدن پلهای متقاطع با رشتههای اکتین است. علاوه بر این، شروع انقباض در پاسخ به یونهای کلسیم بسیار کندتر از ماهیچههای اسکلتی است، همانطور که بعداً مورد بحث قرار خواهد گرفت.
Maximum Force of Contraction Is Often Greater in Smooth Muscle Than in Skeletal Muscle. Despite the relatively few myosin filaments in smooth muscle, and despite the slow cycling time of the cross-bridges, the maximum force of contraction of smooth muscle is often greater than that of skeletal muscle, as much as 4 to 6 kg/ cm2 cross-sectional area for smooth muscle in compari- son with 3 to 4 kilograms for skeletal muscle. This great force of smooth muscle contraction results from the pro- Chapter 8 Excitation and Contraction of Smooth Muscle longed period of attachment of the myosin cross-bridges to the actin filaments.
حداکثر نیروی انقباض اغلب در عضلات صاف بیشتر از عضلات اسکلتی است. علیرغم تعداد نسبتاً کمیرشتههای میوزین در عضله صاف، و با وجود زمان چرخش آهسته پلهای متقاطع، حداکثر نیروی انقباض عضله صاف اغلب بیشتر از عضله اسکلتی است، به اندازه ۴ تا ۶ کیلوگرم بر سانتی متر مربع سطح مقطع برای عضلات صاف در مقایسه با ۳ تا ۴ کیلوگرم برای عضلات اسکلتی. این نیروی عظیم انقباض عضله صاف ناشی از دوره طولانی مدت چسبیدن پلهای متقاطع میوزین به رشتههای اکتین است.
Latch Mechanism Facilitates Prolonged Holding of Contractions of Smooth Muscle. Once smooth muscle has developed full contraction, the amount of continu- ing excitation can usually be reduced to far less than the initial level, even though the muscle maintains its full force of contraction. Furthermore, the energy consumed to maintain contraction is often minuscule, sometimes as little as 1/300 of the energy required for comparable sustained skeletal muscle contraction. This mechanism is called the latch mechanism.
مکانیسم چفت نگه داشتن طولانی مدت انقباضات ماهیچه صاف را تسهیل میکند. هنگامیکه عضله صاف انقباض کامل را ایجاد کرد، مقدار تحریک مداوم معمولاً میتواند به بسیار کمتر از سطح اولیه کاهش یابد، حتی اگر عضله تمام نیروی انقباض خود را حفظ کند. علاوه بر این، انرژی مصرف شده برای حفظ انقباض اغلب ناچیز است، گاهی اوقات به اندازه ۱/۳۰۰ انرژی مورد نیاز برای انقباض پایدار عضلانی اسکلتی. به این مکانیسم مکانیسم چفت میگویند.
The importance of the latch mechanism is that it can maintain prolonged tonic contraction in smooth muscle for hours, with little use of energy. Little continued excit- atory signal is required from nerve fibers or hormonal sources.
اهمیت مکانیسم لچ در این است که میتواند انقباض طولانی مدت تونیک در ماهیچه صاف را برای ساعتها و با استفاده کم از انرژی حفظ کند. سیگنال تحریک مداوم کمیاز رشتههای عصبی یا منابع هورمونی مورد نیاز است.
Stress-Relaxation of Smooth Muscle. Another im- portant characteristic of smooth muscle, especially the visceral unitary type of smooth muscle of many hollow organs, is its ability to return to nearly its original force of contraction seconds or minutes after it has been elongated or shortened. For example, a sudden increase in fluid volume in the urinary bladder, thus stretching the smooth muscle in the bladder wall, causes an im- mediate large increase in pressure in the bladder. How- ever, during about the next 15 to 60 seconds, despite continued stretch of the bladder wall, the pressure re- turns almost exactly back to the original level. Then, when the volume is increased by another step, the same effect occurs again.
استرس-آرامش عضلات صاف. یکی دیگر از ویژگیهای مهم عضله صاف، به ویژه نوع واحد احشایی ماهیچه صاف بسیاری از اندامهای توخالی، توانایی آن برای بازگشت تقریباً به نیروی اولیه انقباض چند ثانیه یا چند دقیقه پس از کشیده شدن یا کوتاه شدن آن است. به عنوان مثال، افزایش ناگهانی حجم مایع در مثانه، در نتیجه کشش ماهیچه صاف در دیواره مثانه، باعث افزایش فوری و زیاد فشار در مثانه میشود. با این حال، در حدود ۱۵ تا ۶۰ ثانیه بعد، علیرغم کشش مداوم دیواره مثانه، فشار تقریباً دقیقاً به سطح اولیه باز میگردد. سپس، هنگامیکه صدا یک مرحله دیگر افزایش مییابد، دوباره همان اثر رخ میدهد.
Conversely, when the volume is suddenly decreased, the pressure falls drastically at first but then rises in another few seconds or minutes to or near the original level. These phenomena are called stress-relaxation and reverse stress-relaxation. Their importance is that except for short periods, they allow a hollow organ to main- tain about the same amount of pressure inside its lumen despite sustained large changes in volume.
برعکس، هنگامیکه حجم به طور ناگهانی کاهش مییابد، فشار در ابتدا به شدت کاهش مییابد، اما پس از چند ثانیه یا چند دقیقه دیگر به سطح اولیه یا نزدیک به آن افزایش مییابد. به این پدیدهها تنش-آرامش و استرس-آرامش معکوس میگویند. اهمیت آنها در این است که به جز برای دورههای کوتاه، آنها به اندام توخالی اجازه میدهند تا با وجود تغییرات زیاد در حجم، تقریباً همان مقدار فشار را در داخل لومن خود حفظ کند.
REGULATION OF CONTRACTION BY CALCIUM IONS
As is true for skeletal muscle, the initiating stimulus for most smooth muscle contraction is an increase in intra- cellular calcium ions. This increase can be caused in dif- ferent types of smooth muscle by nerve stimulation of the smooth muscle fiber, hormonal stimulation, stretch of the fiber, or even changes in the chemical environment of the fiber.
تنظیم انقباض توسط یونهای کلسیم
همانطور که برای ماهیچههای اسکلتی صدق میکند، محرک آغاز کننده اکثر انقباضات ماهیچه صاف افزایش یونهای کلسیم درون سلولی است. این افزایش میتواند در انواع مختلف عضله صاف با تحریک عصبی فیبر ماهیچه صاف، تحریک هورمونی، کشش فیبر یا حتی تغییر در محیط شیمیایی فیبر ایجاد شود.
Smooth muscle does not contain troponin, the regu- latory protein that is activated by calcium ions to cause skeletal muscle contraction. Instead, smooth muscle con- traction is activated by an entirely different mechanism, as described in the next section.
ماهیچه صاف حاوی تروپونین نیست، پروتئین تنظیم کننده ای که توسط یونهای کلسیم فعال میشود و باعث انقباض عضلات اسکلتی میشود. در عوض، انقباض عضله صاف با مکانیسم کاملا متفاوتی فعال میشود، همانطور که در بخش بعدی توضیح داده شد.
Calcium lons Combine with Calmodulin to Cause Ac- tivation of Myosin Kinase and Phosphorylation of the Myosin Head. In place of troponin, smooth muscle cells contain a large amount of another regulatory protein called calmodulin (Figure 8-3). Although this protein is similar to troponin, it is different in the manner in which it initiates contraction. Calmodulin initiates contraction by activating the myosin cross-bridges. This activation and subsequent contraction occur in the following sequence:
کلسیم لونها با کالمودولین ترکیب میشوند تا باعث فعال شدن میوزین کیناز و فسفوریلاسیون سر میوزین شوند. به جای تروپونین، سلولهای ماهیچه صاف حاوی مقدار زیادی پروتئین تنظیم کننده دیگر به نام کالمودولین هستند (شکل ۸-۳). اگرچه این پروتئین شبیه تروپونین است، اما از نظر نحوه شروع انقباض متفاوت است. کالمودولین با فعال کردن پلهای متقاطع میوزین شروع به انقباض میکند. این فعال سازی و انقباض بعدی به ترتیب زیر رخ میدهد:
۱. The calcium concentration in the cytosolic fluid of the smooth muscle increases as a result of the influx of calcium from the extracellular fluid through cal- cium channels and/or release of calcium from the sarcoplasmic reticulum.
2. The calcium ions bind reversibly with calmodulin.
3. The calmodulin-calcium complex then joins with and activates myosin light chain kinase, a phospho- rylating enzyme.
4. One of the light chains of each myosin head, called the regulatory chain, becomes phosphorylated in response to this myosin kinase. When this chain is not phosphorylated, the attachment-detachment cycling of the myosin head with the actin filament does not occur. However, when the regulatory chain is phosphorylated, the head has the capability of binding repetitively with the actin filament and pro- ceeding through the entire cycling process of inter- mittent pulls, the same as what occurs for skeletal muscle, thus causing muscle contraction.
۱. غلظت کلسیم در مایع سیتوزولی عضله صاف در نتیجه هجوم کلسیم از مایع خارج سلولی از طریق کانالهای کلسیم و/یا آزاد شدن کلسیم از شبکه سارکوپلاسمیافزایش مییابد.
2. یونهای کلسیم به طور برگشت پذیر با کالمودولین متصل میشوند.
3. سپس کمپلکس کالمودولین-کلسیم با زنجیره سبک کیناز میوزین که یک آنزیم فسفریله کننده است میپیوندد و آن را فعال میکند.
4. یکی از زنجیرههای سبک هر سر میوزین به نام زنجیره تنظیم کننده در پاسخ به این میوزین کیناز فسفریله میشود. هنگامیکه این زنجیره فسفریله نشده باشد، چرخه اتصال- جدا شدن سر میوزین با رشته اکتین رخ نمیدهد. با این حال، هنگامیکه زنجیره تنظیمیفسفریله میشود، سر این قابلیت را دارد که به طور مکرر با رشته اکتین متصل شود و در کل فرآیند دوچرخه سواری کششهای متناوب انجام شود، همان چیزی که برای عضله اسکلتی اتفاق میافتد، بنابراین باعث انقباض عضلانی میشود.
Figure 8-3 Intracellular calcium ion (Ca2+) concentration increases when Ca2+ enters the cell through calcium channels in the cell mem- brane or is released from the sarcoplasmic reticulum. The Ca2+ binds to calmodulin (CaM) to form a Ca2+-CaM complex, which then acti- vates myosin light chain kinase (MLCK). The active MLCK phospho- rylates the myosin light chain, leading to attachment of the myosin head with the actin filament and contraction of the smooth muscle. ADP, Adenosine diphosphate; ATP, adenosine triphosphate; P, phos- phate.
شکل ۸-۳ غلظت یون کلسیم داخل سلولی (Ca2+) زمانی افزایش مییابد که Ca2+ از طریق کانالهای کلسیمیدر غشای سلولی وارد سلول میشود یا از شبکه سارکوپلاسمیآزاد میشود. Ca2+ به کالمودولین (CaM) متصل میشود تا یک کمپلکس Ca2+-CaM تشکیل دهد که سپس زنجیره سبک کیناز میوزین (MLCK) را فعال میکند. MLCK فعال زنجیره سبک میوزین را فسفریله میکند و منجر به اتصال سر میوزین به رشته اکتین و انقباض عضله صاف میشود. ADP، آدنوزین دی فسفات؛ ATP، آدنوزین تری فسفات؛ P، فسفات.
Source of Calcium lons That Cause Contraction
Although the contractile process in smooth muscle, as in skeletal muscle, is activated by calcium ions, the source of the calcium ions differs. An important difference is that the sarcoplasmic reticulum, which provides virtu- ally all the calcium ions for skeletal muscle contraction, is only slightly developed in most smooth muscle. Instead, most of the calcium ions that cause contraction enter the muscle cell from the extracellular fluid at the time of the action potential or other stimulus. That is, the concentra- tion of calcium ions in the extracellular fluid is greater than 10-3 molar, in comparison with less than 10-7 molar inside the smooth muscle cell; this causes rapid diffusion of the calcium ions into the cell from the extracellular fluid when the calcium channels open. The time required for this diffusion to occur averages 200 to 300 millisec- onds and is called the latent period before contraction begins. This latent period is about 50 times as great for smooth muscle as for skeletal muscle contraction.
منبع کلسیم lons که باعث انقباض میشود
اگرچه فرآیند انقباض در عضلات صاف، مانند ماهیچههای اسکلتی، توسط یونهای کلسیم فعال میشود، منبع یونهای کلسیم متفاوت است. یک تفاوت مهم این است که شبکه سارکوپلاسمی، که تقریباً تمام یونهای کلسیم را برای انقباض ماهیچههای اسکلتی فراهم میکند، در اکثر ماهیچههای صاف فقط اندکی توسعه یافته است. در عوض، بیشتر یونهای کلسیم که باعث انقباض میشوند، از مایع خارج سلولی در زمان پتانسیل عمل یا سایر محرکها وارد سلول عضلانی میشوند. یعنی، غلظت یونهای کلسیم در مایع خارج سلولی بیشتر از ۱۰-۳ مولار است، در مقایسه با کمتر از ۱۰-۷ مولر در داخل سلول ماهیچه صاف. این باعث انتشار سریع یونهای کلسیم به داخل سلول از مایع خارج سلولی با باز شدن کانالهای کلسیم میشود. زمان لازم برای این انتشار به طور متوسط ۲۰۰ تا ۳۰۰ میلی ثانیه است و دوره نهفته قبل از شروع انقباض نامیده میشود. این دوره نهفته برای ماهیچه صاف حدود ۵۰ برابر انقباض عضلات اسکلتی است.
Role of the Smooth Muscle Sarcoplasmic Reticulum. Figure 8-4 shows a few slightly developed sarcoplasmic tubules that lie near the cell membrane in some larger smooth muscle cells. Small invaginations of the cell mem- brane, called caveolae, abut the surfaces of these tubules. The caveolae suggest a rudimentary analog of the trans-verse tubule system of skeletal muscle. When an action potential is transmitted into the caveolae, this is believed to excite calcium ion release from the abutting sarcoplas- mic tubules in the same way that action potentials in skel- etal muscle transverse tubules cause release of calcium ions from the skeletal muscle longitudinal sarcoplasmic tubules. In general, the more extensive the sarcoplasmic reticulum in the smooth muscle fiber, the more rapidly it contracts.
نقش شبکه سارکوپلاسمیعضلات صاف. شکل ۸-۴ چند لوله سارکوپلاسمیکمیتوسعه یافته را نشان میدهد که در نزدیکی غشای سلولی در برخی از سلولهای عضله صاف بزرگتر قرار دارند. فرورفتگیهای کوچک غشای سلولی به نام caveolae، در مجاورت سطوح این لولهها قرار دارند. caveolae یک آنالوگ ابتدایی از سیستم لوله عرضی عضله اسکلتی را نشان میدهد. هنگامیکه یک پتانسیل عمل به داخل کاوئول منتقل میشود، اعتقاد بر این است که این امر باعث تحریک آزاد شدن یون کلسیم از لولههای سارکوپلاسمیمجاور میشود، به همان صورتی که پتانسیل عمل در لولههای عرضی عضله اسکلتی باعث آزاد شدن یونهای کلسیم از لولههای سارکوپلاسمیطولی عضله اسکلتی میشود. به طور کلی، هرچه شبکه سارکوپلاسمیدر فیبر ماهیچه صاف گسترده تر باشد، سریعتر منقبض میشود.
Figure 8-4
Sarcoplasmic tubules in a large smooth muscle fiber showing their relation to invaginations in the cell membrane called caveolae.
شکل ۸-۴
لولههای سارکوپلاسمیدر یک فیبر عضلانی صاف بزرگ که رابطه خود را با فرورفتگی در غشای سلولی به نام caveolae نشان میدهد.
Smooth Muscle Contraction Is Dependent on Extra- cellular Calcium Ion Concentration. Whereas chang- ing the extracellular fluid calcium ion concentration from normal has little effect on the force of contraction of skeletal muscle, this is not true for most smooth muscle. When the extracellular fluid calcium ion concentration decreases to about 1/3 to 1/10 normal, smooth muscle contraction usually ceases. Therefore, the force of con- traction of smooth muscle is usually highly dependent on the extracellular fluid calcium ion concentration.
انقباض عضلات صاف به غلظت یون کلسیم خارج سلولی بستگی دارد. در حالی که تغییر غلظت یون کلسیم مایع خارج سلولی از نرمال تأثیر کمیبر نیروی انقباض عضله اسکلتی دارد، این برای اکثر عضلات صاف صادق نیست. هنگامیکه غلظت یون کلسیم مایع خارج سلولی به حدود ۱/۳ تا ۱/۱۰ نرمال کاهش مییابد، انقباض عضلات صاف معمولاً متوقف میشود. بنابراین، نیروی انقباض عضله صاف معمولاً به شدت به غلظت یون کلسیم مایع خارج سلولی وابسته است.
A Calcium Pump Is Required to Cause Smooth Muscle Relaxation. To cause relaxation of smooth muscle after it has contracted, the calcium ions must be removed from the intracellular fluids. This removal is achieved by a cal- cium pump that pumps calcium ions out of the smooth muscle fiber back into the extracellular fluid, or into a sarcoplasmic reticulum, if it is present (Figure 8-5). This pump requires ATP and is slow acting in comparison with the fast-acting sarcoplasmic reticulum pump in skeletal muscle. Therefore, a single smooth muscle contraction of- ten lasts for seconds rather than hundredths to tenths of a second, as occurs for skeletal muscle.
پمپ کلسیم برای ایجاد آرامش عضلات صاف مورد نیاز است. برای ایجاد شل شدن عضلات صاف پس از انقباض، یونهای کلسیم باید از مایعات داخل سلولی خارج شوند. این حذف توسط یک پمپ کلسیم انجام میشود که یونهای کلسیم را از فیبر ماهیچه صاف به داخل مایع خارج سلولی یا در صورت وجود به شبکه سارکوپلاسمیپمپ میکند (شکل ۸-۵). این پمپ به ATP نیاز دارد و در مقایسه با پمپ شبکه سارکوپلاسمیسریع الاثر در عضله اسکلتی کند عمل میکند. بنابراین، یک انقباض عضله صاف اغلب برای چند ثانیه طول میکشد تا صدم تا دهم ثانیه، همانطور که برای ماهیچههای اسکلتی اتفاق میافتد.
Figure 8-5
Relaxation of smooth muscle occurs when the calcium ion (Ca2+) concentration decreases below a critical level as Ca2+ is pumped out of the cell or into the sarcoplasmic reticulum. Ca2+ is then released from calmodulin (CaM), and myosin phosphatase removes phosphate from the myosin light chain, causing detach- ment of the myosin head from the actin filament and relaxation of the smooth muscle. ADP, Adenosine diphosphate; ATP, adenosine triphosphate; Na+, sodium; P, phosphate.
شکل ۸-۵
شل شدن عضلات صاف زمانی اتفاق میافتد که غلظت یون کلسیم (Ca2+) به زیر سطح بحرانی کاهش مییابد، زیرا Ca2+ از سلول یا داخل شبکه سارکوپلاسمیپمپ میشود. سپس Ca2+ از کالمودولین (CaM) آزاد میشود و میوزین فسفاتاز فسفات را از زنجیره سبک میوزین حذف میکند و باعث جدا شدن سر میوزین از رشته اکتین و شل شدن عضله صاف میشود. ADP، آدنوزین دی فسفات؛ ATP، آدنوزین تری فسفات؛ Na+، سدیم؛ P، فسفات.
Myosin Phosphatase Is Important in Cessation of Con- traction. Relaxation of the smooth muscle occurs when the calcium channels close and the calcium pump trans- ports calcium ions out of the cytosolic fluid of the cell. When the calcium ion concentration falls below a critical level, the aforementioned processes automatically reverse, except for the phosphorylation of the myosin head. Re- versal of this situation requires another enzyme, myosin phosphatase (see Figure 8-5), located in the cytosol of the smooth muscle cell, which splits the phosphate from the regulatory light chain. Then the cycling stops, and contrac- tion ceases. The time required for the relaxation of muscle contraction, therefore, is determined to a great extent by the amount of active myosin phosphatase in the cell.
میوزین فسفاتاز در توقف انقباض مهم است. شل شدن ماهیچه صاف زمانی اتفاق میافتد که کانالهای کلسیم بسته میشوند و پمپ کلسیم یونهای کلسیم را از مایع سیتوزولی سلول خارج میکند. هنگامیکه غلظت یون کلسیم به زیر سطح بحرانی میرسد، فرآیندهای فوق به طور خودکار معکوس میشوند، به جز فسفوریلاسیون سر میوزین. معکوس شدن این وضعیت نیازمند آنزیم دیگری به نام میوزین فسفاتاز (نگاه کنید به شکل ۸-۵) است که در سیتوزول سلول عضله صاف قرار دارد و فسفات را از زنجیره سبک تنظیمیجدا میکند. سپس دوچرخه سواری متوقف میشود و انقباض متوقف میشود. بنابراین زمان مورد نیاز برای شل شدن انقباض عضلانی تا حد زیادی توسط مقدار میوزین فسفاتاز فعال در سلول تعیین میشود.
Possible Mechanism for Regulating the Latch Phe- nomenon. Because of the importance of the latch phe- nomenon in smooth muscle, and because this phenom- enon allows for the long-term maintenance of tone in many smooth muscle organs without much expenditure of energy, many attempts have been made to explain it. Among the many mechanisms that have been postulated, one of the simplest is the following.
مکانیسم ممکن برای تنظیم پدیده چفت. به دلیل اهمیت پدیده لچ در عضله صاف، و به دلیل اینکه این پدیده امکان حفظ طولانی مدت تون را در بسیاری از اندامهای ماهیچه صاف بدون صرف انرژی زیاد میدهد، تلاشهای زیادی برای توضیح آن صورت گرفته است. در میان بسیاری از مکانیسمهایی که فرض شده است، یکی از ساده ترین آنها موارد زیر است.
When the myosin kinase and myosin phosphatase enzymes are both strongly activated, the cycling frequency of the myosin heads and the velocity of contraction are great. Then, as activation of the enzymes decreases, the cycling frequency decreases but, at the same time, the deactivation of these enzymes allows the myosin heads to remain attached to the actin filament for a longer and lon- ger proportion of the cycling period. Therefore, the num- ber of heads attached to the actin filament at any given time remains large. Because the number of heads attached to the actin determines the static force of contraction, ten- sion is maintained, or latched, yet little energy is used by the muscle because ATP is not degraded to ADP, except on the rare occasion when a head detaches.
هنگامیکه آنزیمهای میوزین کیناز و میوزین فسفاتاز هر دو به شدت فعال میشوند، فرکانس چرخش سرهای میوزین و سرعت انقباض زیاد است. سپس، با کاهش فعال شدن آنزیمها، فرکانس چرخش کاهش مییابد، اما در عین حال، غیرفعال شدن این آنزیمها به سرهای میوزین اجازه میدهد تا برای مدت طولانی تر و طولانی تری از دوره دوچرخه سواری به رشته اکتین متصل بمانند. بنابراین، تعداد سرهای متصل به رشته اکتین در هر زمان معین زیاد باقی میماند. از آنجایی که تعداد سرهای متصل به اکتین، نیروی ساکن انقباض را تعیین میکند، تنش حفظ میشود یا بسته میشود، اما انرژی کمیتوسط عضله استفاده میشود زیرا ATP به ADP تجزیه نمیشود، مگر در موارد نادری که سر جدا میشود.
NERVOUS AND HORMONAL CONTROL OF SMOOTH MUSCLE CONTRACTION
Although skeletal muscle fibers are stimulated exclusively by the nervous system, smooth muscle can be stimulated to contract by nervous signals, hormonal stimulation, stretch of the muscle, and several other ways. The prin- cipal reason for the difference is that the smooth muscle membrane contains many types of receptor proteins that can initiate the contractile process. Still other recep- tor proteins inhibit smooth muscle contraction, which is another difference from skeletal muscle. Therefore, in this section, we discuss nervous control of smooth mus- cle contraction, followed by hormonal control and other means of control.
کنترل عصبی و هورمونی انقباض ماهیچههای صاف
اگرچه فیبرهای عضلانی اسکلتی منحصراً توسط سیستم عصبی تحریک میشوند، عضله صاف را میتوان با سیگنالهای عصبی، تحریک هورمونی، کشش عضله و چندین روش دیگر تحریک کرد. دلیل اصلی این تفاوت این است که غشای عضله صاف حاوی انواع مختلفی از پروتئینهای گیرنده است که میتوانند فرآیند انقباض را آغاز کنند. پروتئینهای گیرنده دیگر انقباض عضلات صاف را مهار میکنند که این تفاوت دیگری با ماهیچههای اسکلتی است. بنابراین، در این بخش، کنترل عصبی انقباض عضله صاف و به دنبال آن کنترل هورمونی و سایر ابزارهای کنترل را مورد بحث قرار میدهیم.
NEUROMUSCULAR JUNCTIONS OF SMOOTH MUSCLE
اتصالات عصبی عضلانی ماهیچههای صاف
Physiologic Anatomy of Smooth Muscle Neuromus- cular Junctions. Neuromuscular junctions of the highly structured type found on skeletal muscle fibers do not oc- cur in smooth muscle. Instead, the autonomic nerve fibers that innervate smooth muscle generally branch diffusely on top of a sheet of muscle fibers, as shown in Figure 8-6. In most cases, these fibers do not make direct contact with the smooth muscle fiber cell membranes but instead form diffuse junctions that secrete their transmitter substance into the matrix coating of the smooth muscle, often a few nanometers to a few micrometers away from the muscle cells. The transmitter substance then diffuses to the cells. Furthermore, where there are many layers of muscle cells, the nerve fibers often innervate only the outer layer. Mus- cle excitation travels from this outer layer to the inner lay- ers by action potential conduction in the muscle mass or by additional diffusion of the transmitter substance.
آناتومیفیزیولوژیک اتصالات عصبی عضلانی عضلات صاف. اتصالات عصبی عضلانی از نوع بسیار ساختار یافته که در فیبرهای عضلانی اسکلتی یافت میشود در ماهیچه صاف رخ نمیدهد. درعوض، رشتههای عصبی خودمختار که عضله صاف را عصب دهی میکنند، عموماً به طور پراکنده در بالای ورقه ای از رشتههای عضلانی منشعب میشوند، همانطور که در شکل ۸-۶ نشان داده شده است. در بیشتر موارد، این فیبرها تماس مستقیمیبا غشای سلولی فیبر ماهیچه صاف ندارند، اما در عوض اتصالات پراکندهای ایجاد میکنند که ماده فرستنده خود را در پوشش ماتریکس عضله صاف ترشح میکنند، که اغلب در فاصله چند نانومتری تا چند میکرومتری از سلولهای عضلانی است. سپس ماده فرستنده به سلولها منتشر میشود. علاوه بر این، در جایی که لایههای زیادی از سلولهای عضلانی وجود دارد، رشتههای عصبی اغلب فقط لایه بیرونی را عصب میکنند. تحریک عضلانی از این لایه بیرونی به لایههای داخلی توسط هدایت پتانسیل عمل در توده عضلانی یا با انتشار اضافی ماده فرستنده حرکت میکند.
The axons that innervate smooth muscle fibers do not have the typical branching end feet of the type found in the motor end plate on skeletal muscle fibers. Instead, most of the fine terminal axons have multiple varicosi- ties distributed along their axes. At these points, the Schwann cells that envelop the axons are interrupted so that transmitter substance can be secreted through the walls of the varicosities. In the varicosities are vesicles similar to those in the skeletal muscle end plate that con- tain transmitter substance. However, in contrast to the vesicles of skeletal muscle junctions, which always con- tain acetylcholine, the vesicles of the autonomic nerve fiber endings contain acetylcholine in some fibers and norepinephrine in others and occasionally other sub- stances as well.
آکسونهایی که فیبرهای عضلانی صاف را عصب دهی میکنند، دارای پایههای انتهایی انشعاب معمولی از نوع موجود در صفحه انتهایی حرکتی فیبرهای عضلانی اسکلتی نیستند. در عوض، اکثر آکسونهای انتهایی ظریف دارای واریسهای متعددی هستند که در امتداد محورهای خود توزیع شده اند. در این نقاط، سلولهای شوان که آکسونها را میپوشانند، قطع میشوند تا ماده فرستنده از طریق دیوارههای واریس ترشح شود. در واریسها، وزیکولهایی شبیه به وزیکولهای موجود در صفحه انتهایی عضله اسکلتی وجود دارد که حاوی ماده فرستنده است. با این حال، بر خلاف وزیکولهای اتصالات ماهیچههای اسکلتی، که همیشه حاوی استیل کولین هستند، وزیکولهای انتهای رشتههای عصبی خودمختار حاوی استیل کولین در برخی از رشتهها و نوراپی نفرین در برخی دیگر و گاهی اوقات مواد دیگر نیز هستند.
In a few cases, particularly in the multi-unit type of smooth muscle, the varicosities are separated from the muscle cell membrane by as little as 20 to 30 nanome- ters-the same width as the synaptic cleft that is found in the skeletal muscle junction. These are called contact junctions, and they function in much the same way as the skeletal muscle neuromuscular junction. The rapidity of contraction of these smooth muscle fibers is consid- erably faster than that of fibers stimulated by the diffuse junctions.
در چند مورد، به ویژه در نوع چند واحدی ماهیچه صاف، واریسها از غشای سلول عضلانی به اندازه ۲۰ تا ۳۰ نانومتر جدا میشوند – همان عرض شکاف سیناپسی که در محل اتصال ماهیچههای اسکلتی یافت میشود. اینها اتصالات تماسی نامیده میشوند و عملکرد آنها تقریباً مانند اتصال عصبی عضلانی عضله اسکلتی است. سرعت انقباض این فیبرهای عضلانی صاف به طور قابل توجهی سریعتر از الیاف تحریک شده توسط اتصالات منتشر است.
Figure 8-6
Innervation of smooth muscle by autonomic nerve fib- ers that branch diffusely and secrete neurotransmitter from multiple varicosities. Unitary (visceral) smooth muscle cells are connected by gap junctions so that depolarization can rapidly spread from one cell to another, permitting the muscle cells to contract as a single unit. In multi-unit smooth muscle, each cell is stimulated independently by a neurotransmitter released from closely associated autonomic nerve varicosities.
شکل ۸-۶
عصب دهی عضله صاف توسط رشتههای عصبی خودمختار که به طور منتشر منشعب میشوند و انتقال دهندههای عصبی را از واریسهای متعدد ترشح میکنند. سلولهای عضله صاف واحد (احشایی) با اتصالات شکافی به هم متصل میشوند، به طوری که دپلاریزاسیون میتواند به سرعت از یک سلول به سلول دیگر گسترش یابد و به سلولهای عضلانی اجازه میدهد تا به صورت یک واحد منقبض شوند. در عضله صاف چند واحدی، هر سلول به طور مستقل توسط یک انتقال دهنده عصبی آزاد شده از واریکوسیته عصب اتونوم نزدیک مرتبط تحریک میشود.
Excitatory and Inhibitory Transmitter Substances Se- creted at the Smooth Muscle Neuromuscular Junc- tion. The most important transmitter substances secret- ed by the autonomic nerves innervating smooth muscle are acetylcholine and norepinephrine, but they are never secreted by the same nerve fibers. Acetylcholine is an ex- citatory transmitter substance for smooth muscle fibers in some organs but an inhibitory transmitter for smooth muscle in other organs. When acetylcholine excites a muscle fiber, norepinephrine ordinarily inhibits it. Con- versely, when acetylcholine inhibits a fiber, norepineph- rine usually excites it.
مواد فرستنده تحریکی و مهاری که در محل اتصال عصبی عضلانی ماهیچه صاف ترشح میشوند. مهمترین مواد فرستنده ترشح شده توسط اعصاب اتونومیک عضله صاف استیل کولین و نوراپی نفرین هستند، اما هرگز توسط رشتههای عصبی مشابهی ترشح نمیشوند. استیل کولین یک ماده انتقال دهنده تحریک کننده برای فیبرهای عضلانی صاف در برخی از اندامها است، اما یک فرستنده بازدارنده برای عضلات صاف در سایر اندامها است. هنگامیکه استیل کولین فیبر عضلانی را تحریک میکند، نوراپی نفرین معمولاً آن را مهار میکند. برعکس، وقتی استیل کولین فیبر را مهار میکند، نوراپی نفرین معمولاً آن را تحریک میکند.
Why are these responses different? The answer is that both acetylcholine and norepinephrine excite or inhibit smooth muscle by first binding with a receptor protein on the surface of the muscle cell membrane. Some of the receptor proteins are excitatory receptors, whereas others are inhibitory receptors. Thus, the type of receptor deter- mines whether the smooth muscle is inhibited or excited and also determines which of the two transmitters, ace- tylcholine or norepinephrine, is effective in causing the excitation or inhibition. These receptors are discussed in more detail in Chapter 61 in regard to function of the autonomic nervous system.
چرا این پاسخها متفاوت است؟ پاسخ این است که هم استیل کولین و هم نوراپی نفرین با اتصال به پروتئین گیرنده در سطح غشای سلول عضلانی، عضله صاف را تحریک یا مهار میکنند. برخی از پروتئینهای گیرنده گیرندههای تحریک کننده هستند، در حالی که برخی دیگر گیرندههای بازدارنده هستند. بنابراین، نوع گیرنده تعیین میکند که آیا ماهیچه صاف مهار یا برانگیخته شده است و همچنین تعیین میکند که کدام یک از دو فرستنده، استیل کولین یا نوراپی نفرین، در ایجاد تحریک یا مهار مؤثر است. این گیرندهها با جزئیات بیشتری در فصل ۶۱ در رابطه با عملکرد سیستم عصبی خودمختار مورد بحث قرار میگیرند.
MEMBRANE POTENTIALS AND ACTION POTENTIALS IN SMOOTH MUSCLE
پتانسیلهای غشاء و پتانسیلهای عمل در ماهیچههای صاف
Membrane Potentials in Smooth Muscle. The quantitative voltage of the membrane potential of smooth muscle depends on the momentary condition of the muscle. In the normal resting state, the intracellular potential is usually about -50 to -60 millivolts, which is about 30 millivolts less negative than in skeletal muscle.
پتانسیلهای غشایی در عضلات صاف. ولتاژ کمیپتانسیل غشایی عضله صاف به وضعیت لحظه ای عضله بستگی دارد. در حالت استراحت طبیعی، پتانسیل درون سلولی معمولاً حدود ۵۰- تا ۶۰- میلی ولت است که حدود ۳۰ میلی ولت کمتر از ماهیچههای اسکلتی منفی است.
Action Potentials in Unitary Smooth Muscle. Action potentials occur in unitary smooth muscle (e.g., visceral muscle) in the same way that they occur in skeletal muscle. They do not normally occur in most multi-unit types of smooth muscle, as discussed in a subsequent section.
پتانسیلهای عمل در عضله صاف واحد. پتانسیل عمل در ماهیچه صاف واحد (مثلاً عضله احشایی) به همان شکلی که در عضله اسکلتی رخ میدهد رخ میدهد. همانطور که در بخش بعدی توضیح داده شد، معمولاً در اکثر انواع ماهیچههای صاف چند واحدی رخ نمیدهند.
The action potentials of visceral smooth muscle occur in one of two forms-(1) spike potentials or (2) action potentials with plateaus.
پتانسیل عمل عضله صاف احشایی به یکی از دو شکل (۱) پتانسیل سنبله یا (۲) پتانسیل عمل با پلاتو رخ میدهد.
Spike Potentials. Typical spike action potentials, such as those seen in skeletal muscle, occur in most types of unitary smooth muscle. The duration of this type of action potential is 10 to 50 milliseconds, as shown in Figure 8-7A. Such action potentials can be elicited in many ways-for example, by electrical stimulation, by the action of hormones on the smooth muscle, by the action of transmitter substances from nerve fibers, by stretch, or as a result of spontaneous generation in the muscle fiber itself, as discussed subsequently.
پتانسیلهای سنبله. پتانسیل عمل سنبله معمولی، مانند آنهایی که در عضله اسکلتی مشاهده میشود، در اکثر انواع عضله صاف واحد رخ میدهد. مدت زمان این نوع پتانسیل عمل ۱۰ تا ۵۰ میلی ثانیه است که در شکل ۸-۷ الف نشان داده شده است. چنین پتانسیلهای عملی را میتوان به روشهای مختلفی برانگیخت – به عنوان مثال، با تحریک الکتریکی، با عمل هورمونها بر روی ماهیچه صاف، با عمل مواد فرستنده از رشتههای عصبی، با کشش، یا در نتیجه تولید خود به خود در فیبر عضلانی، همانطور که در ادامه بحث شد.
Figure 8-7
A, Typical smooth muscle action potential (spike potential) elicited by an external stimulus. B, Repetitive spike potentials, elicited by slow rhythmical electrical waves that occur spontaneously in the smooth muscle of the intestinal wall. C, Action potential with a plateau, recorded from a smooth muscle fiber of the uterus.
شکل ۸-۷
A، پتانسیل عمل عضله صاف معمولی (پتانسیل سنبله) که توسط یک محرک خارجی ایجاد میشود. ب، پتانسیلهای پیک تکراری، که توسط امواج الکتریکی ریتمیک آهسته که به طور خود به خود در عضله صاف دیواره روده ایجاد میشود، ایجاد میشود. ج، پتانسیل عمل با یک پلاتو، ثبت شده از فیبر عضلانی صاف رحم.
Action Potentials with Plateaus. Figure 8-7C shows a smooth muscle action potential with a plateau. The onset of this action potential is similar to that of the typical spike potential. However, instead of rapid repolarization of the muscle fiber membrane, the repolarization is delayed for several hundred to as much as 1000 milliseconds (1 second). The importance of the plateau is that it can account for the prolonged contraction that occurs in some types of smooth muscle, such as the ureter, the uterus under Chapter 8 Excitation and Contraction of Smooth Muscle some conditions, and certain types of vascular smooth muscle. Also, this is the type of action potential seen in cardiac muscle fibers that have a prolonged period of con- traction, as discussed in Chapters 9 and 10.
پتانسیلهای عمل با فلاتها. شکل ۸-7C پتانسیل عمل عضله صاف را با یک فلات نشان میدهد. شروع این پتانسیل عمل شبیه به پتانسیل سنبله معمولی است. با این حال، به جای رپولاریزاسیون سریع غشای فیبر عضلانی، رپلاریزاسیون برای چند صد تا ۱۰۰۰ میلی ثانیه (۱ ثانیه) به تعویق میافتد. اهمیت پلاتو در این است که میتواند علت انقباض طولانی مدتی باشد که در برخی از انواع ماهیچههای صاف مانند حالب، رحم تحت فصل ۸ تحریک و انقباض ماهیچه صاف برخی شرایط و انواع خاصی از عضلات صاف عروق رخ میدهد. همچنین، این نوع پتانسیل عملی است که در فیبرهای عضلانی قلبی که دوره طولانی انقباض دارند، همانطور که در فصلهای ۹ و ۱۰ بحث شد، مشاهده میشود.
Calcium Channels Are Important in Generating the Smooth Muscle Action Potential. The smooth muscle cell membrane has far more voltage-gated calcium channels than skeletal muscle but few voltage-gated sodium channels. Therefore, sodium does not participate much in the generation of the action potential in most smooth muscle. Instead, the flow of calcium ions to the interior of the fiber is mainly responsible for the action potential. This flow occurs in the same self-regenerative way as occurs for the sodium channels in nerve fibers and in skeletal muscle fibers. However, the calcium channels open many times more slowly than sodium channels, and they also remain open much longer. These characteristics largely account for the prolonged plateau action potentials of some smooth muscle fibers.
کانالهای کلسیم در ایجاد پتانسیل عمل ماهیچههای صاف مهم هستند. غشای سلولی عضله صاف کانالهای کلسیمیبسیار بیشتری نسبت به ماهیچههای اسکلتی دارد، اما کانالهای سدیم دارای ولتاژ کمیدارند. بنابراین، سدیم در تولید پتانسیل عمل در اکثر عضلات صاف شرکت چندانی ندارد. در عوض، جریان یونهای کلسیم به داخل فیبر عمدتاً مسئول پتانسیل عمل است. این جریان به همان روشی که برای کانالهای سدیم در رشتههای عصبی و در رشتههای عضلانی اسکلتی رخ میدهد، اتفاق میافتد. با این حال، کانالهای کلسیم چندین برابر کندتر از کانالهای سدیم باز میشوند و همچنین برای مدت طولانی تری باز میمانند. این ویژگیها تا حد زیادی پتانسیلهای طولانیمدت عمل فلات برخی از فیبرهای عضلانی صاف را توضیح میدهند.
Another important feature of calcium ion entry into the cells during the action potential is that the calcium ions act directly on the smooth muscle contractile mechanism to cause contraction. Thus, the calcium performs two tasks at once.
کی دیگر از ویژگیهای مهم ورود یون کلسیم به داخل سلولها در طی پتانسیل عمل این است که یونهای کلسیم مستقیماً بر روی مکانیسم انقباض ماهیچه صاف عمل کرده و باعث انقباض میشوند. بنابراین، کلسیم دو وظیفه را همزمان انجام میدهد.
Slow Wave Potentials in Unitary Smooth Muscle Can Lead to Spontaneous Generation of Action Potentials. Some smooth muscle is self-excitatory—that is, action potentials arise within the smooth muscle cells with- out an extrinsic stimulus. This activity is often associated with a basic slow wave rhythm of the membrane potential. A typical slow wave in a visceral smooth muscle of the gut is shown in Figure 8-7B. The slow wave is not the action potential. That is, it is not a self-regenerative process that spreads progressively over the membranes of the muscle fibers. Instead, it is a local property of the smooth muscle fibers that make up the muscle mass.
پتانسیلهای موج آهسته در عضله صاف یکپارچه میتواند منجر به تولید خودبه خودی پتانسیلهای عمل شود. برخی از عضلات صاف خود تحریککننده هستند، یعنی پتانسیلهای عمل در سلولهای ماهیچه صاف بدون محرک بیرونی ایجاد میشوند. این فعالیت اغلب با یک ریتم پایه موج آهسته پتانسیل غشا همراه است. یک موج آهسته معمولی در عضله صاف احشایی روده در شکل ۸-7B نشان داده شده است. موج آهسته پتانسیل عمل نیست. یعنی این یک فرآیند خودبازسازی نیست که به تدریج روی غشاهای فیبرهای عضلانی پخش شود. در عوض، این خاصیت محلی فیبرهای عضلانی صاف است که توده عضلانی را تشکیل میدهند.
The cause of the slow wave rhythm is unknown. One suggestion is that the slow waves are caused by waxing and waning of the pumping of positive ions (presumably sodium ions) outward through the muscle fiber mem- brane. That is, the membrane potential becomes more negative when sodium is pumped rapidly and less negative when the sodium pump becomes less active. Another suggestion is that the conductances of the ion channels increase and decrease rhythmically.
علت ریتم امواج آهسته ناشناخته است. یک پیشنهاد این است که امواج آهسته ناشی از اپیلاسیون و کاهش پمپاژ یونهای مثبت (احتمالاً یونهای سدیم) به بیرون از طریق غشای فیبر عضلانی است. یعنی وقتی سدیم سریع پمپ میشود پتانسیل غشا منفی تر میشود و وقتی پمپ سدیم کمتر فعال میشود منفی میشود. پیشنهاد دیگر این است که رسانایی کانالهای یونی به صورت ریتمیک افزایش و کاهش مییابد.
The importance of the slow waves is that when they are strong enough, they can initiate action potentials. The slow waves themselves cannot cause muscle contraction. However, when the peak of the negative slow wave potential inside the cell membrane rises in the positive direction, from -60 to about -35 millivolts (the approximate threshold for eliciting action potentials in most vis-ceral smooth muscle), an action potential develops and spreads over the muscle mass and contraction occurs. Figure 8-7B demonstrates this effect, showing that at each peak of the slow wave, one or more action potentials occur. These repetitive sequences of action potentials elicit rhythmical contraction of the smooth muscle mass. Therefore, the slow waves are called pacemaker waves. In Chapter 63, we see that this type of pacemaker activity controls the rhythmical contractions of the gut.
اهمیت امواج آهسته در این است که وقتی به اندازه کافی قوی هستند، میتوانند پتانسیلهای عمل را آغاز کنند. امواج آهسته خود نمیتوانند باعث انقباض عضلانی شوند. با این حال، هنگامیکه اوج پتانسیل موج آهسته منفی در داخل غشای سلولی در جهت مثبت از ۶۰- تا حدود ۳۵- میلی ولت افزایش مییابد (آستانه تقریبی برای برانگیختن پتانسیلهای عمل در بیشتر عضلات صاف احشایی)، پتانسیل عمل ایجاد میشود و بر روی توده عضلانی پخش میشود و انقباض رخ میدهد. شکل ۸-7B این اثر را نشان میدهد و نشان میدهد که در هر پیک موج آهسته، یک یا چند پتانسیل عمل رخ میدهد. این توالیهای تکراری پتانسیلهای عمل باعث انقباض ریتمیک توده عضلانی صاف میشود. بنابراین امواج آهسته را امواج ضربان ساز مینامند. در فصل ۶۳ میبینیم که این نوع فعالیت پیس میکر انقباضات ریتمیک روده را کنترل میکند.
Excitation of Visceral Smooth Muscle by Muscle Stretch. When visceral (unitary) smooth muscle is stretched sufficiently, spontaneous action potentials are usually generated. They result from a combination of the following: (1) the normal slow wave potentials; and (2) a decrease in overall negativity of the membrane potential caused by the stretch. This response to stretch allows the gut wall, when excessively stretched, to contract automatically and rhythmically. For example, when the gut is overfilled by intestinal contents, local automatic contractions often set up peristaltic waves that move the contents away from the overfilled intestine, usually in the direction of the anus.
تحریک عضله صاف احشایی با کشش عضلانی. وقتی عضله صاف احشایی (یونیتی) به اندازه کافی کشیده شود، معمولاً پتانسیلهای عمل خود به خودی ایجاد میشود. آنها از ترکیبی از موارد زیر حاصل میشوند: (۱) پتانسیل موج آهسته معمولی. و (۲) کاهش منفی کلی پتانسیل غشا ناشی از کشش. این پاسخ به کشش باعث میشود که دیواره روده، زمانی که بیش از حد کشیده میشود، به طور خودکار و ریتمیک منقبض شود. به عنوان مثال، زمانی که روده بیش از حد از محتویات روده پر میشود، انقباضات خودکار موضعی اغلب امواج پریستالتیک ایجاد میکند که محتویات را از روده پر شده دور میکند، معمولاً در جهت مقعد.
DEPOLARIZATION OF MULTI-UNIT SMOOTH MUSCLE WITHOUT ACTION POTENTIALS
The smooth muscle fibers of multi-unit smooth muscle (e.g., the muscle of the iris of the eye or the piloerector muscle of each hair) normally contract mainly in response to nerve stimuli. The nerve endings secrete acetylcholine in the case of some multi-unit smooth muscles and norepinephrine in the case of others. In both cases, the transmitter substances cause depolarization of the smooth muscle membrane, and this depolarization in turn elicits contraction. Action potentials usually do not develop because the fibers are too small to generate an action potential. (When action potentials are elicited in visceral unitary smooth muscle, 30 to 40 smooth muscle fibers must depolarize simultaneously before a self-propagating action potential ensues.) However, in small smooth muscle cells, even with- out an action potential, the local depolarization (called the junctional potential) caused by the nerve transmitter sub- stance spreads “electrotonically” over the entire fiber and is all that is necessary to cause muscle contraction.
دپولاریزاسیون عضله صاف چند واحدی بدون پتانسیل عمل
فیبرهای عضلانی صاف عضله صاف چند واحدی (مثلاً عضله عنبیه چشم یا عضله پیلوئرکتور هر مو) معمولاً در پاسخ به محرکهای عصبی منقبض میشوند. پایانههای عصبی در مورد برخی از عضلات صاف چند واحدی استیل کولین و در مورد برخی دیگر نوراپی نفرین ترشح میکنند. در هر دو مورد، مواد فرستنده باعث دپلاریزاسیون غشای عضله صاف میشوند و این دپلاریزاسیون به نوبه خود باعث انقباض میشود. پتانسیل عمل معمولاً ایجاد نمیشود زیرا الیاف برای ایجاد پتانسیل عمل بسیار کوچک هستند. (زمانی که پتانسیل عمل در عضله صاف واحد احشایی برانگیخته میشود، ۳۰ تا ۴۰ فیبر عضله صاف باید به طور همزمان دپلاریزه شوند قبل از اینکه پتانسیل عمل خود تکثیر پیدا کند.) با این حال، در سلولهای کوچک عضله صاف، حتی بدون پتانسیل عمل، دپلاریزاسیون موضعی (به نام پتانسیل پیوندی) ناشی از انتشار کل رشته عصبی از طریق فشار عصبی است. و تمام چیزی است که برای ایجاد انقباض عضلانی لازم است.
Local Tissue Factors and Hormones Can Cause Smooth Muscle Contraction Without Action Potentials
Approximately half of all smooth muscle contraction is likely initiated by stimulatory factors acting directly on the smooth muscle contractile machinery and without action potentials. Two types of non-nervous and nonaction potential stimulating factors often involved are (1) local tissue chemical factors and (2) various hormones.
عوامل بافتی و هورمونهای محلی میتوانند باعث انقباض عضلات صاف بدون پتانسیل عمل شوند.
تقریباً نیمیاز تمام انقباضات ماهیچه صاف احتمالاً توسط عوامل محرکی شروع میشود که مستقیماً بر روی دستگاه انقباض ماهیچه صاف و بدون پتانسیل عمل تأثیر میگذارد. دو نوع از عوامل تحریک کننده بالقوه غیر عصبی و غیرعملی اغلب درگیر هستند (۱) عوامل شیمیایی بافت محلی و (۲) هورمونهای مختلف.
Smooth Muscle Contraction in Response to Local Tis- sue Chemical Factors. In Chapter 17, we discuss control of contraction of the arterioles, meta-arterioles, and pre- capillary sphincters. The smallest of these vessels have little or no nervous supply. Yet, the smooth muscle is highly contractile, responding rapidly to changes in local chemical conditions in the surrounding interstitial fluid and to stretch caused by changes in blood pressure.
انقباض ماهیچه صاف در پاسخ به عوامل شیمیایی بافت محلی. در فصل ۱۷، کنترل انقباض شریانها، متا شریانها و اسفنکترهای پیش مویرگی را مورد بحث قرار میدهیم. کوچکترین این رگها منبع عصبی کمیدارند یا اصلاً وجود ندارند. با این حال، عضله صاف بسیار انقباض است و به سرعت به تغییرات شرایط شیمیایی موضعی در مایع بینابینی اطراف و کشش ناشی از تغییرات فشار خون پاسخ میدهد.
In the normal resting state, many of these small blood vessels remain contracted. However, when extra blood flow to the tissue is necessary, multiple factors can relax the vessel wall, thus allowing for increased flow. In this way, a powerful local feedback control system controls the blood flow to the local tissue area. Some of the specific control factors are as follows:
در حالت استراحت طبیعی، بسیاری از این رگهای خونی کوچک منقبض میمانند. با این حال، زمانی که جریان خون اضافی به بافت ضروری است، عوامل متعددی میتوانند دیواره رگ را شل کنند و در نتیجه باعث افزایش جریان میشوند. به این ترتیب، یک سیستم کنترل بازخورد محلی قدرتمند، جریان خون را در ناحیه بافت محلی کنترل میکند. برخی از عوامل کنترل خاص به شرح زیر است:
۱. Lack of oxygen in the local tissues causes smooth muscle relaxation and, therefore, vasodilation.
2. Excess carbon dioxide causes vasodilation.
3. Increased hydrogen ion concentration causes vasodilation.
۱. کمبود اکسیژن در بافتهای موضعی باعث شل شدن عضلات صاف و در نتیجه اتساع عروق میشود.
2. دی اکسید کربن اضافی باعث اتساع عروق میشود.
3. افزایش غلظت یون هیدروژن باعث اتساع عروق میشود.
Adenosine, lactic acid, increased potassium ions, nitric oxide, and increased body temperature can all cause local vasodilation. Decreased blood pressure, by causing decreased stretch of the vascular smooth muscle, also causes these small blood vessels to dilate.
آدنوزین، اسید لاکتیک، افزایش یون پتاسیم، اکسید نیتریک و افزایش دمای بدن همگی میتوانند باعث اتساع موضعی عروق شوند. کاهش فشار خون، با کاهش کشش ماهیچه صاف عروق، باعث گشاد شدن این عروق خونی کوچک نیز میشود.
Effects of Hormones on Smooth Muscle Contraction. Many circulating hormones in the blood affect smooth muscle contraction to some degree, and some have pro- found effects. Among the more important of these hormones are norepinephrine, epinephrine, angiotensin II, endothelin, vasopressin, oxytocin, serotonin, and histamine.
اثرات هورمونها بر انقباض عضلات صاف. بسیاری از هورمونهای در گردش خون تا حدودی بر انقباض عضلات صاف تأثیر میگذارند و برخی نیز اثرات آشکاری دارند. از مهمترین این هورمونها میتوان به نوراپی نفرین، اپی نفرین، آنژیوتانسین II، اندوتلین، وازوپرسین، اکسی توسین، سروتونین و هیستامین اشاره کرد.
A hormone causes contraction of a smooth muscle when the muscle cell membrane contains hormone-gated excitatory receptors for the respective hormone. Conversely, the hormone causes inhibition if the membrane contains inhibitory receptors for the hormone rather than excitatory receptors.
زمانی که غشای سلولی عضلانی حاوی گیرندههای تحریک کننده هورمون مربوطه باشد، یک هورمون باعث انقباض ماهیچه صاف میشود. برعکس، اگر غشاء حاوی گیرندههای بازدارنده هورمون باشد نه گیرندههای تحریک کننده، هورمون باعث مهار میشود.
Mechanisms of Smooth Muscle Excitation or Inhibition by Hormones or Local Tissue Factors. Some hormone receptors in the smooth muscle membrane open sodium or calcium ion channels and depolarize the mem- brane, the same as after nerve stimulation. Sometimes, action potentials result, or action potentials that are al- ready occurring may be enhanced. In other cases, depolarization occurs without action potentials, and this depolarization allows for calcium ion entry into the cell, which promotes the contraction.
مکانیسمهای تحریک یا مهار عضلات صاف توسط هورمونها یا فاکتورهای بافتی محلی. برخی از گیرندههای هورمونی در غشای عضله صاف، کانالهای یونی سدیم یا کلسیم را باز میکنند و غشاء را مانند پس از تحریک عصبی دپولاریزه میکنند. گاهی اوقات، پتانسیلهای عمل منتج میشوند، یا پتانسیلهای عملی که از قبل در حال رخ دادن هستند، ممکن است تقویت شوند. در موارد دیگر، دپلاریزاسیون بدون پتانسیل عمل رخ میدهد و این دپلاریزاسیون اجازه ورود یون کلسیم به سلول را میدهد که باعث افزایش انقباض میشود.
Inhibition, in contrast, occurs when the hormone (or other tissue factor) closes the sodium and calcium channels to prevent entry of these positive ions; inhibition also occurs if the normally closed potassium channels are opened, allowing positive potassium ions to diffuse out of the cell. Both these actions increase the degree of negativity inside the muscle cell, a state called hyperpolarization, which strongly inhibits muscle contraction.
در مقابل، مهار زمانی اتفاق میافتد که هورمون (یا سایر عوامل بافتی) کانالهای سدیم و کلسیم را ببندد تا از ورود این یونهای مثبت جلوگیری کند. همچنین اگر کانالهای پتاسیم بسته به طور معمول باز شوند، بازداری میشود و به یونهای مثبت پتاسیم اجازه میدهد به خارج از سلول منتشر شوند. هر دوی این اقدامات درجه منفی بودن را در داخل سلول عضلانی افزایش میدهند، حالتی به نامهایپرپلاریزاسیون، که به شدت انقباض عضلانی را مهار میکند.
Sometimes, smooth muscle contraction or inhibition is initiated by hormones without directly causing any change in the membrane potential. In these cases, the hormone may activate a membrane receptor that does not open any ion channels but, instead, causes an internal change in the muscle fiber, such as release of calcium ions from the intracellular sarcoplasmic reticulum; the calcium then induces contraction. To inhibit contraction, other receptor mechanisms are known to activate the enzyme adenylate cyclase or guanylate cyclase in the cell membrane. The portions of the receptors that protrude to the interior of the cells are coupled to these enzymes, causing the for-mation of cyclic adenosine monophosphate (cAMP) or cyclic guanosine monophosphate (cGMP), so-called second messengers. cAMP or cGMP has many effects, one of which is to change the degree of phosphorylation of several enzymes that indirectly inhibit contraction. The pump that moves calcium ions from the sarcoplasm into the sarcoplasmic reticulum is activated, as well as the cell membrane pump that moves calcium ions out of the cell; these effects reduce the calcium ion concentration in the sarcoplasm, thereby inhibiting contraction.
گاهی اوقات، انقباض یا مهار عضلات صاف توسط هورمونها بدون ایجاد تغییر مستقیم در پتانسیل غشاء آغاز میشود. در این موارد، هورمون ممکن است گیرنده غشایی را فعال کند که هیچ کانال یونی را باز نمیکند، اما در عوض باعث ایجاد تغییر درونی در فیبر عضلانی میشود، مانند آزاد شدن یونهای کلسیم از شبکه سارکوپلاسمیداخل سلولی. سپس کلسیم باعث انقباض میشود. برای مهار انقباض، مکانیسمهای گیرنده دیگری شناخته شدهاند که آنزیم آدنیلات سیکلاز یا گوانیلات سیکلاز را در غشای سلولی فعال میکنند. بخشهایی از گیرندههایی که به داخل سلولها بیرون زدهاند با این آنزیمها جفت میشوند و باعث تشکیل آدنوزین مونوفسفات حلقوی (cAMP) یا گوانوزین مونوفسفات حلقوی (cGMP) میشوند که به اصطلاح پیامرسانهای دوم نامیده میشوند. cAMP یا cGMP اثرات زیادی دارد که یکی از آنها تغییر درجه فسفوریلاسیون چندین آنزیم است که به طور غیر مستقیم انقباض را مهار میکنند. پمپی که یونهای کلسیم را از سارکوپلاسم به شبکه سارکوپلاسمیمنتقل میکند، و همچنین پمپ غشای سلولی که یونهای کلسیم را از سلول خارج میکند، فعال میشود. این اثرات باعث کاهش غلظت یون کلسیم در سارکوپلاسم میشود و در نتیجه انقباض را مهار میکند.
Smooth muscles have considerable diversity in how they initiate contraction or relaxation in response to different hormones, neurotransmitters, and other sub- stances. In some cases, the same substance may cause either relaxation or contraction of smooth muscles in different locations. For example, norepinephrine inhibits contraction of smooth muscle in the intestine but stimulates contraction of smooth muscle in blood vessels.
ماهیچههای صاف در نحوه شروع انقباض یا آرامش در پاسخ به هورمونهای مختلف، انتقال دهندههای عصبی و سایر مواد تنوع قابل توجهی دارند. در برخی موارد، همان ماده ممکن است باعث شل شدن یا انقباض عضلات صاف در نقاط مختلف شود. به عنوان مثال، نوراپی نفرین انقباض ماهیچه صاف روده را مهار میکند اما انقباض ماهیچه صاف در رگهای خونی را تحریک میکند.
Bibliography
کتابشناسی
Also see the bibliography for Chapters 5 and 6.
Behringer EJ, Segal SS: Spreading the signal for vasodilatation: im- plications for skeletal muscle blood flow control and the effects of aging. J Physiol 590:6277, 2012.
Berridge MJ: Smooth muscle cell calcium activation mechanisms. J Physiol 586:5047, 2008.
Blaustein MP, Lederer WJ: Sodium/calcium exchange: its physiological implications. Physiol Rev 79:763, 1999.
Chapter 8 Excitation and Contraction of Smooth Muscle
Brozovich FV, Nicholson CJ, Degen CV, Gao YZ, Aggarwal M, Mor- gan KG: Mechanisms of vascular smooth muscle contraction and the basis for pharmacologic treatment of smooth muscle disorders. Pharmacol Rev 68:476, 2016.
Burnstock G. Purinergic signaling in the cardiovascular system. Circ Res 120:207, 2017.
Cheng H, Lederer WJ: Calcium sparks. Physiol Rev 88:1491, 2008. Davis MJ: Perspective: physiological role(s) of the vascular myogenic response. Microcirculation 19:99, 2012.
Dopico AM, Bukiya AN, Jaggar JH. Calcium- and voltage-gated BK channels in vascular smooth muscle. Pflugers Arch 470:1271, 2018.
Dora KA. Endothelial-smooth muscle cell interactions in the regulation of vascular tone in skeletal muscle. Microcirculation 23:626, 2016.
Drummond HA, Grifoni SC, Jernigan NL: A new trick for an old dogma: ENaC proteins as mechanotransducers in vascular smooth muscle. Physiology (Bethesda) 23:23, 2008.
Hill MA, Meininger GA. Small artery mechanobiology: roles of cellular and non-cellular elements. Microcirculation 23:611, 2016.
Huizinga JD, Lammers WJ: Gut peristalsis is governed by a multitude of cooperating mechanisms. Am J Physiol Gastrointest Liver Physiol 296:G1, 2009.
Kauffenstein G, Laher I, Matrougui K, et al: Emerging role of G protein-coupled receptors in microvascular myogenic tone. Cardio- vasc Res 95:223, 2012.
Lacolley P, Regnault V, Segers P, Laurent S. Vascular smooth muscle cells and arterial stiffening: relevance in development, aging, and disease. Physiol Rev 97:1555, 2017.
Morgan KG, Gangopadhyay SS: Cross-bridge regulation by thin filament-associated proteins. J Appl Physiol 91:953, 2001.
Ratz PH. Mechanics of vascular smooth muscle. Compr Physiol 6:111, 2015.
Sanders KM, Kito Y, Hwang SJ, Ward SM. Regulation of gastroin- testinal smooth muscle function by interstitial cells. Physiology (Bethesda) 31:316, 2016.
Somlyo AP, Somlyo AV: Ca2+ sensitivity of smooth muscle and non- muscle myosin II: modulated by G proteins, kinases, and myosin phosphatase. Physiol Rev 83:1325, 2003.
Tykocki NR, Boerman EM, Jackson WF. Smooth muscle ion channels and regulation of vascular tone in resistance arteries and arterioles. Compr Physiol 7:485, 2017.
Webb RC: Smooth muscle contraction and relaxation. Adv Physiol Educ 27:201, 2003.
109
تحریک و انقباض ماهیچه صاف
انواع ماهیچههای صاف
عضله صاف هر اندام از جهات مختلفی از سایر اندامها متمایز است: (۱) ابعاد فیزیکی، (۲) سازماندهی به بستهها یا صفحات، (۳) پاسخ به انواع مختلف محرکها، (۴) ویژگیهای عصب، و (۵) عملکرد. با این حال، برای سادگی، عضله صاف را میتوان به طور کلی به دو نوع عمده تقسیم کرد که در شکل ۸-۱ نشان داده شده است: عضله صاف چندواحدی (multiunit) و عضله صاف واحد یا تکواحدی (single-unit).
 شکل ۸-۱ عضله صاف چند واحدی (A) و تکواحدی (B).
شکل ۸-۱ عضله صاف چند واحدی (A) و تکواحدی (B).
ماهیچه صاف چند واحدی
این نوع ماهیچه صاف از فیبرهای عضلانی صاف مجزا تشکیل شده است. هر فیبر مستقل از فیبرهای دیگر عمل میکند و اغلب توسط یک انتهای عصبی، عصبدهی میشود، همانطور که برای فیبرهای عضلانی اسکلتی اتفاق میافتد. علاوه بر این، سطوح خارجی این الیاف، مانند فیبرهای عضلانی اسکلتی، توسط یک لایه نازک از ماده شبیه غشای پایه، مخلوطی از کلاژن نازک و گلیکوپروتئین پوشیده شده است که به عایق بندی الیاف جداگانه از یکدیگر کمک میکند.
مهمترین ویژگی فیبرهای عضلانی صاف چند واحدی این است که هر فیبر میتواند مستقل از بقیه منقبض شود و کنترل آنها عمدتاً توسط سیگنالهای عصبی اعمال میشود. در مقابل، سهم عمدهای از کنترل عضله صاف واحد توسط محرکهای غیرعصبی اعمال میشود. نمونههایی از این نوع ماهیچه صاف عبارتند از: عضله مژگانی چشم، عنبیه چشم و ماهیچههای پیلوئرکتور (piloerector muscles) یا ماهیچههای سیخ کننده مو که با تحریک سیستم عصبی سمپاتیک باعث راست شدن موها میشوند.
عضله صاف تکواحدی
به این نوع عضله، ماهیچه صاف سنسیشیال (syncytial smooth muscle) یا عضله صاف احشایی (visceral smooth muscle) نیز گفته میشود. اصطلاح “یونیتاری” (unitary) گیج کننده است زیرا به معنای فیبرهای عضلانی منفرد نیست. در عوض، این به معنای توده ای متشکل از صدها تا هزاران فیبر عضلانی صاف است که به عنوان یک واحد با هم منقبض میشوند. فیبرها معمولاً در صفحات یا بستههایی قرار میگیرند و غشای سلولی آنها در چندین نقطه به یکدیگر میچسبند تا نیروی ایجاد شده در یک فیبر عضلانی به رشته بعدی منتقل شود. علاوه بر این، غشاهای سلولی با اتصالات شکافی (Gap Junction) زیادی به هم میپیوندند که از طریق آن یونها میتوانند آزادانه از یک سلول عضلانی به سلول دیگر جریان پیدا کنند، به طوری که پتانسیلهای عمل یا جریان یونی ساده بدون پتانسیل عمل میتوانند از یک فیبر به فیبر دیگر حرکت کنند و باعث انقباض فیبرهای عضلانی با هم شوند. این نوع ماهیچه صاف به دلیل اتصالات متقابل سنسیشیال بین فیبرها به عضله صاف سنسیشیال نیز معروف است. به آن ماهیچه صاف احشایی نیز می گویند زیرا در دیوارههای اکثر احشاء بدن از جمله دستگاه گوارش، مجاری صفراوی، حالب، رحم و بسیاری از رگهای خونی یافت میشود.
مکانیسم انقباض در عضلات صاف
مبنای شیمیایی برای انقباض عضلات صاف
ماهیچه صاف دارای هر دو رشته اکتین و میوزین است که دارای ویژگیهای شیمیایی مشابه رشتههای اکتین و میوزین در ماهیچههای اسکلتی است. این شامل کمپلکس تروپونین طبیعی نیست که در کنترل انقباض عضلات اسکلتی مورد نیاز است، بنابراین مکانیسم کنترل انقباض متفاوت است. این موضوع بعداً در این فصل به تفصیل مورد بحث قرار میگیرد.
مطالعات شیمیایی نشان دادهاند که رشتههای اکتین و میوزین مشتق شده از ماهیچههای صاف، به همان شیوهای که در ماهیچههای اسکلتی انجام میدهند، با یکدیگر تعامل دارند. علاوه بر این، فرآیند انقباض توسط یونهای کلسیم فعال میشود و آدنوزین تری فسفات (ATP) به آدنوزین دی فسفات (ADP) تجزیه میشود تا انرژی برای انقباض فراهم شود.
با این حال، تفاوتهای عمدهای بین سازمان فیزیکی ماهیچه صاف و ماهیچه اسکلتی وجود دارد، همچنین تفاوتهایی در جفت شدن تحریک-انقباض، کنترل فرآیند انقباض توسط یونهای کلسیم، مدت زمان انقباض و مقدار انرژی مورد نیاز برای انقباض وجود دارد.
مبنای فیزیکی برای انقباض عضلات صاف
عضله صاف فاقد آرایش مخطط رشتههای اکتین و میوزین است که در ماهیچههای اسکلتی وجود دارد. در عوض، تکنیکهای میکروگرافیک الکترونی سازمان فیزیکی نشان داده شده در شکل ۸-۲ را پیشنهاد میکنند. این شکل تعداد زیادی رشته اکتین متصل به اجسام به اصطلاح متراکم (dense bodies) را نشان میدهد. برخی از این اجسام به غشای سلولی متصل هستند. برخی دیگر در داخل سلول پراکنده میشوند. برخی از اجسام غشایی سلولهای مجاور توسط پلهای پروتئینی بین سلولی به یکدیگر متصل میشوند. عمدتاً از طریق این پیوندها است که نیروی انقباض از یک سلول به سلول دیگر منتقل میشود.
 شکل ۸-۲ ساختار فیزیکی ماهیچه صاف. فیبر سمت چپ بالایی رشتههای اکتین را نشان میدهد که به صورت شعاعی از اجسام متراکم خارج شدهاند. فیبر پایین سمت چپ و نمودار سمت راست رابطه رشتههای میوزین را با رشتههای اکتین نشان میدهد.
شکل ۸-۲ ساختار فیزیکی ماهیچه صاف. فیبر سمت چپ بالایی رشتههای اکتین را نشان میدهد که به صورت شعاعی از اجسام متراکم خارج شدهاند. فیبر پایین سمت چپ و نمودار سمت راست رابطه رشتههای میوزین را با رشتههای اکتین نشان میدهد.
در میان رشتههای اکتین در فیبر عضلانی رشتههای میوزین قرار دارند. قطر آنها بیش از دو برابر رشتههای اکتین است. در میکروگرافهای الکترونی، معمولاً رشتههای اکتینی نسبت به رشتههای میوزین ۵ تا ۱۰ برابر میباشند.
در سمت راست شکل ۸-۲ یک ساختار فرضی از یک واحد انقباضی منفرد در یک سلول ماهیچه صاف نشان داده شده است. در هر واحد انقباضی سلول ماهیچه صاف تعداد زیادی رشته اکتین به صورت شعاعی از دو جسم متراکم خارج شدهاند. انتهای این رشتهها بر روی یک رشته میوزین که در میانه راه بین اجسام متراکم قرار دارد همپوشانی دارند. این واحد انقباضی شبیه به واحد انقباضی عضله اسکلتی است، اما بدون منظم بودن ساختار عضله اسکلتی. در واقع، اجسام متراکم ماهیچههای صاف همان نقش دیسکها و صفحههای Z را در عضله اسکلتی ایفا میکنند.
تفاوت دیگری نیز وجود دارد: بیشتر رشتههای میوزین دارای پلهای عرضی «قطبی کناری» (sidepolar) هستند که بهگونهای مرتب شدهاند که پلهای یک طرف در یک جهت و پلهای طرف دیگر در جهت مخالف قرار میگیرند. این به میوزین اجازه میدهد تا یک رشته اکتین را در یک جهت در یک سمت بکشد در حالی که همزمان رشته دیگری را در جهت مخالف در سمت دیگر بکشد. ارزش این سازمان این است که به سلولهای ماهیچه صاف اجازه میدهد تا ۸۰ درصد طول خود را منقبض کنند، بهجای اینکه به کمتر از ۳۰ درصد محدود شوند، همانطور که در ماهیچههای اسکلتی اتفاق میافتد. [فاصله مفید انقباض عضله اسکلتی تنها حدود یک چهارم تا یک سوم طول زمان استراحت آن است در حالیکه عضله صاف غالباً میتواند خود را بیش از دو سوم طول کشیده اش منقبض کند یعنی درصد کوتاه شدن عضله صاف بیش از درصد کوتاه شدن عضله اسکلتی است]
مقایسه انقباض عضلات صاف و انقباض عضلات اسکلتی
اگرچه اکثر ماهیچههای اسکلتی به سرعت منقبض و شل میشوند، اکثر انقباضات ماهیچه صاف انقباض طولانی مدت تونیک است که گاهی چند ساعت یا حتی چند روز طول میکشد. بنابراین، انتظار میرود که هر دو ویژگی فیزیکی و شیمیایی عضله صاف در مقایسه با انقباض عضلات اسکلتی متفاوت باشد. در زیر به برخی از تفاوتها اشاره میشود:
چرخه کند پلهای عرضی میوزین
سرعت چرخش پلهای عرضی میوزین در ماهیچه صاف – یعنی اتصال آنها به اکتین، سپس آزاد شدن از اکتین و اتصال مجدد برای چرخه بعدی – بسیار کندتر از عضله اسکلتی است. در واقع، فرکانس آن به اندازه ۱/۱۰ تا ۱/۳۰۰ در عضله اسکلتی است. با این حال اعتقاد بر این است که کسری از زمانی که پلهای عرضی به رشتههای اکتین متصل میمانند، که عامل اصلی تعیین کننده نیروی انقباض است، در عضله صاف بسیار افزایش مییابد. یکی از دلایل احتمالی چرخه کُند این است که سرهای پل عرضی نسبت به ماهیچههای اسکلتی فعالیت ATPase بسیار کمتری دارند، بنابراین تخریب ATP که به حرکات سر پلهای عرضی انرژی میدهد همزمان با کاهش سرعت مربوط به جرخه، تا حد زیادی کاهش مییابد.
نیاز به انرژی کم برای حفظ انقباض عضلات صاف
انرژی لازم برای حفظ انقباض در عضله صاف تنها ۱/۱۰ تا ۱/۳۰۰ انرژی لازم در عضله اسکلتی است. اعتقاد بر این است که این نیز ناشی از اتصال آهسته و چرخه جداشدگی پلهای عرضی است و به این دلیل که برای هر چرخه، صرف نظر از مدت زمان، تنها یک مولکول ATP مورد نیاز است.
این مصرف کم انرژی توسط عضلات صاف برای صرفه جویی انرژی کلی بدن بسیار مهم است، زیرا اندامهایی مانند روده، مثانه، کیسه صفرا و سایر احشاء اغلب انقباض تونیک عضلات را تقریباً به طور نامحدود حفظ میکنند.
کندی شروع انقباض و شل شدن بافت ماهیچه صاف
یک بافت عضله صاف معمولی ۵۰ تا ۱۰۰ میلی ثانیه پس از تحریک شدن شروع به انقباض میکند، حدود ۰.۵ ثانیه بعد به انقباض کامل میرسد و سپس در ۱ تا ۲ ثانیه دیگر از نیروی انقباضی کاسته میشود و مجموع زمان انقباض آن ۱ تا ۳ ثانیه است. این حدود ۳۰ برابر طولانیتر از انقباض یک فیبر عضلانی اسکلتی است. اما از آنجایی که ماهیچههای صاف انواع زیادی دارند، انقباض برخی از انواع آن میتواند ۰.۲ ثانیه یا ۳۰ ثانیه باشد.
شروع آهسته انقباض ماهیچه صاف و همچنین انقباض طولانی مدت آن به دلیل کندی اتصال و جدا شدن پلهای عرضی با رشتههای اکتین است. علاوه بر این، شروع انقباض در پاسخ به یونهای کلسیم بسیار کندتر از ماهیچههای اسکلتی است، همانطور که لحظاتی بعد بحث میشود.
اغلب در عضلات صاف حداکثر نیروی انقباض بیشتر از عضلات اسکلتی است.
علیرغم تعداد نسبتاً کم رشتههای میوزین در عضله صاف، و با وجود زمان چرخه آهسته پلهای عرضی، حداکثر نیروی انقباض عضله صاف اغلب بیشتر از عضله اسکلتی است – برای عضلات صاف، به اندازهٔ ۴ تا ۶ کیلوگرم بر سانتی متر مربع سطح مقطع عرضی در مقایسه با ۳ تا ۴ کیلوگرم سطح مقطع عرضی برای عضلات اسکلتی. این نیروی بزرگ انقباض ماهیچههای صاف ناشی از دوره طولانی اتصال پلهای عرضی میوزین به رشتههای اکتین است.
مکانیسم “قفل شدن” حفظ طولانیمدت انقباضات ماهیچه صاف را تسهیل میکند
هنگامیکه عضله صاف انقباض کامل را ایجاد کرد، مقدار تحریک مداوم معمولاً میتواند به بسیار کمتر از سطح اولیه کاهش یابد، اما عضله نیروی کامل انقباض خود را حفظ میکند. علاوه بر این، انرژی مصرف شده برای حفظ انقباض اغلب ناچیز است، گاهی اوقات به اندازه ۱/۳۰۰ انرژی مورد نیاز برای انقباض پایدار عضلات اسکلتی. این مکانیسم “قفل شدن” (Latch) نامیده میشود.
اهمیت مکانیسم قفل شدن در این است که میتواند انقباضات تونیک طولانی مدت در ماهیچه صاف را با وجود مصرف کم انرژی برای ساعتها حفظ کند. در ضمن سیگنال تحریکی کمیاز رشتههای عصبی یا منابع هورمونی مورد نیاز است.
تنش-آرامش عضلات صاف
یکی دیگر از ویژگیهای مهم عضله صاف، به ویژه نوع واحد احشایی ماهیچه صاف بسیاری از اندامهای توخالی، توانایی آن برای بازگشت تقریباً به نیروی اولیه انقباض چند ثانیه یا چند دقیقه پس از کشیده شدن یا کوتاه شدن آن است. به عنوان مثال، افزایش ناگهانی حجم مایع در مثانه، در نتیجه کشش ماهیچه صاف در دیواره مثانه، باعث افزایش فوری فشار در مثانه میشود. با این حال، در طول ۱۵ ثانیه تا یک دقیقه یا بیشتر، با وجود ادامه کشش دیواره مثانه، فشار تقریباً دقیقاً به سطح اولیه باز میگردد. سپس، هنگامیکه حجم یک مرحله دیگر افزایش مییابد، دوباره همان اثر رخ میدهد.
برعکس، هنگامیکه حجم به طور ناگهانی کاهش مییابد، فشار در ابتدا به شدت کاهش مییابد اما پس از چند ثانیه یا چند دقیقه دیگر به سطح اولیه یا نزدیک به آن افزایش مییابد. به این پدیدهها تنش-آرامش (stress-relaxation) و تنش-آرامش معکوس (reverse stress-relaxation) میگویند. اهمیت آنها در این است که، به جز برای دورههای زمانی کوتاه، به اندام توخالی اجازه میدهند تا با وجود تغییرات طولانی مدت و زیاد در حجم، تقریباً همان مقدار فشار را در داخل لومن خود حفظ کند.
تنظیم انقباض توسط یونهای کلسیم
همانطور که در مورد ماهیچههای اسکلتی صدق میکند، محرک آغاز کننده اکثر انقباضات عضلات صاف افزایش یونهای کلسیم داخل سلولی است. این افزایش میتواند در انواع مختلف عضله صاف به دلیل تحریک عصبی فیبر ماهیچه صاف، تحریک هورمونی، کشش فیبر و یا حتی تغییر در محیط شیمیایی فیبر ایجاد شود.
با این حال ماهیچه صاف حاوی تروپونین نیست، پروتئین تنظیمیکه توسط یونهای کلسیم فعال میشود و باعث انقباض عضلات اسکلتی میشود. در عوض، انقباض عضله صاف با مکانیسم کاملا متفاوتی فعال میشود، به شرح زیر.
ترکیب یونهای کلسیم با کالمودولین باعث فعال شدن میوزین کیناز و فسفوریلاسیون سر میوزین میشود.
به جای تروپونین، سلولهای ماهیچه صاف حاوی مقدار زیادی پروتئین تنظیم کننده دیگر به نام کالمودولین هستند (شکل ۸-۳). اگرچه این پروتئین شبیه تروپونین است، اما از نظر نحوه شروع انقباض متفاوت است. کالمودولین این کار را با فعال کردن پلهای متقاطع میوزین انجام میدهد. این فعال سازی و انقباض بعدی به ترتیب زیر رخ میدهد:
۱. یونهای کلسیم با کالمودولین متصل میشوند.
۲. سپس کمپلکس کالمودولین-کلسیم با زنجیره سبک کیناز میوزین که یک آنزیم فسفریله کننده است میپیوندد و آن را فعال میکند.
۳. یکی از زنجیرههای سبک هر سر میوزین به نام زنجیره تنظیم کننده در پاسخ به این میوزین کیناز فسفریله میشود. هنگامیکه این زنجیره فسفریله نشده باشد، چرخه اتصال- جدا شدن سر میوزین با رشته اکتین رخ نمیدهد. اما هنگامیکه زنجیره تنظیمیفسفریله میشود، سر این قابلیت را دارد که به طور مکرر با رشته اکتین متصل شود و در کل فرآیند دوچرخه سواری “کششهای” متناوب انجام شود، همان چیزی که برای ماهیچههای اسکلتی اتفاق میافتد، بنابراین باعث انقباض عضلانی میشود.
 شکل ۸-۳ غلظت یون کلسیم داخل سلولی (++Ca) زمانی افزایش مییابد که ++Ca از طریق کانالهای کلسیم در غشای سلولی یا شبکه سارکوپلاسمی(SR) وارد سلول میشود. ++Ca به کالمودولین متصل میشود تا یک کمپلکس Ca++ -calmodulin تشکیل دهد که سپس زنجیره سبک کیناز میوزین (MLCK) را فعال میکند. MLCK زنجیره سبک میوزین (MLC) را فسفریله میکند که منجر به انقباض عضله صاف میشود. هنگامیکه غلظت ++Ca کاهش مییابد، به دلیل پمپاژ کلسیم به خارج از سلول، این روند معکوس میشود و میوزین فسفاتاز فسفات را از MLC حذف میکند و منجر به آرامش میشود.
شکل ۸-۳ غلظت یون کلسیم داخل سلولی (++Ca) زمانی افزایش مییابد که ++Ca از طریق کانالهای کلسیم در غشای سلولی یا شبکه سارکوپلاسمی(SR) وارد سلول میشود. ++Ca به کالمودولین متصل میشود تا یک کمپلکس Ca++ -calmodulin تشکیل دهد که سپس زنجیره سبک کیناز میوزین (MLCK) را فعال میکند. MLCK زنجیره سبک میوزین (MLC) را فسفریله میکند که منجر به انقباض عضله صاف میشود. هنگامیکه غلظت ++Ca کاهش مییابد، به دلیل پمپاژ کلسیم به خارج از سلول، این روند معکوس میشود و میوزین فسفاتاز فسفات را از MLC حذف میکند و منجر به آرامش میشود.
میوزین فسفاتاز در قطع انقباض مهم است
هنگامیکه غلظت یون کلسیم به زیر سطح بحرانی میرسد، فرآیندهای فوق بهجز فسفوریلاسیون سر میوزین به طور خودکار معکوس میشوند. معکوس شدن این امر به آنزیم دیگری به نام میوزین فسفاتاز (نگاه کنید به شکل ۸-۳) نیاز دارد که در سیتوزول سلول عضله صاف قرار دارد و فسفات را از زنجیره سبک تنظیمیجدا میکند. سپس دوچرخه سواری متوقف میشود و انقباض متوقف میشود. بنابراین زمان مورد نیاز برای شل شدن انقباض عضلانی تا حد زیادی با مقدار میوزین فسفاتاز فعال در سلول تعیین میشود.
مکانیسم ممکن برای تنظیم پدیده چفت
به دلیل اهمیت پدیده لچ در عضله صاف و به دلیل اینکه این پدیده امکان حفظ طولانی مدت تون در بسیاری از اندامهای ماهیچه صاف را بدون صرف انرژی زیاد میدهد، تلاشهای زیادی برای توضیح آن صورت گرفته است. در میان بسیاری از مکانیسمهایی که فرض شده است، یکی از ساده ترین آنها موارد زیر است.
هنگامیکه آنزیمهای میوزین کیناز و میوزین فسفاتاز هر دو به شدت فعال میشوند، فرکانس چرخش سرهای میوزین و سرعت انقباض زیاد است. سپس، با کاهش فعال شدن آنزیمها، فرکانس چرخش کاهش مییابد، اما در عین حال، غیرفعال شدن این آنزیمها به سرهای میوزین اجازه میدهد تا برای مدت طولانی تر و طولانی تری از دوره دوچرخه سواری به رشته اکتین متصل بمانند. بنابراین، تعداد سرهای متصل به رشته اکتین در هر زمان معین زیاد باقی میماند. از آنجا که تعداد سرهای متصل به اکتین، نیروی ساکن انقباض را تعیین میکند، کشش حفظ میشود یا “چفت” میشود. اما انرژی کمیتوسط عضله استفاده میشود زیرا ATP به ADP تجزیه نمیشود مگر در موارد نادری که سر جدا میشود.
کنترل عصبی و هورمونی انقباض عضلات صاف
اگرچه فیبرهای عضلانی اسکلتی منحصراً توسط سیستم عصبی تحریک میشوند، ماهیچه صاف را میتوان با انواع مختلفی از سیگنالها تحریک کرد تا منقبض شوند: با سیگنالهای عصبی، توسط تحریک هورمونی، با کشش عضله، و به چندین روش دیگر. دلیل اصلی این تفاوت این است که غشای عضله صاف حاوی انواع مختلفی از پروتئینهای گیرنده است که میتوانند فرآیند انقباض را آغاز کنند. پروتئینهای گیرنده دیگر انقباض عضلات صاف را مهار میکنند که تفاوت دیگری با ماهیچههای اسکلتی است. بنابراین در این قسمت به کنترل عصبی انقباض عضلات صاف و به دنبال آن کنترل هورمونی و سایر روشهای کنترل میپردازیم.
اتصالات عصبی عضلانی عضلات صاف
آناتومیفیزیولوژیک اتصالات عصبی عضلانی عضلات صاف
اتصالات عصبی عضلانی از نوع بسیار ساختار یافته که در فیبرهای عضلانی اسکلتی یافت میشود در ماهیچه صاف رخ نمیدهد. درعوض، رشتههای عصبی خودمختار که عضله صاف را عصب دهی میکنند، عموماً به طور پراکنده در بالای ورقه ای از رشتههای عضلانی منشعب میشوند، همانطور که در شکل ۸-۴ نشان داده شده است. در بیشتر موارد، این فیبرها تماس مستقیمیبا غشای سلولی فیبر عضلانی صاف ندارند، اما در عوض به اصطلاح اتصالات پراکنده ایجاد میکنند. که ماده فرستنده خود را در پوشش ماتریکس عضله صاف ترشح میکنند که اغلب در فاصله چند نانومتری تا چند میکرومتری از سلولهای ماهیچه ای قرار دارند. سپس ماده فرستنده به سلولها منتشر میشود. علاوه بر این، در جایی که لایههای زیادی از سلولهای عضلانی وجود دارد، رشتههای عصبی اغلب فقط لایه بیرونی را عصب میکنند. تحریک عضلانی از این لایه بیرونی به لایههای داخلی با هدایت پتانسیل عمل در توده عضلانی یا با انتشار اضافی ماده فرستنده حرکت میکند.
 شکل ۸-۴ عصب دهی ماهیچه صاف.
شکل ۸-۴ عصب دهی ماهیچه صاف.
آکسونهایی که فیبرهای عضلانی صاف را عصب دهی میکنند، دارای پایههای انتهایی انشعاب معمولی از نوع صفحه انتهایی حرکتی روی رشتههای عضلانی اسکلتی نیستند. در عوض، اکثر آکسونهای انتهایی ظریف دارای واریسهای متعددی هستند که در امتداد محورهای خود توزیع شده اند. در این نقاط سلولهای شوان که آکسونها را میپوشانند قطع میشوند تا ماده فرستنده از طریق دیوارههای واریکوسیته ترشح شود. در واریسها وزیکولهایی شبیه به وزیکولهای موجود در صفحه انتهایی عضله اسکلتی وجود دارد که حاوی ماده فرستنده است. اما بر خلاف وزیکولهای محل اتصال ماهیچههای اسکلتی که همیشه حاوی استیل کولین هستند، وزیکولهای انتهای رشتههای عصبی خودمختار حاوی استیل کولین در برخی رشتهها و نوراپی نفرین هستند. در سایرین – و گاهاً مواد دیگر نیز.
در موارد معدودی، بهویژه در نوع چند واحدی ماهیچههای صاف، واریکوسیتهها از غشای سلول عضلانی به اندازه ۲۰ تا ۳۰ نانومتر جدا میشوند – همان عرض شکاف سیناپسی که در محل اتصال ماهیچههای اسکلتی رخ میدهد. اینها اتصالات تماسی نامیده میشوند و عملکرد آنها تقریباً مشابه اتصال عصبی عضلانی عضله اسکلتی است. سرعت انقباض این فیبرهای عضلانی صاف به طور قابل توجهی سریعتر از الیاف تحریک شده توسط اتصالات منتشر است.
مواد فرستنده تحریکی و بازدارنده ترشح شده در محل اتصال عصبی عضلانی ماهیچه صاف
مهم ترین مواد فرستنده ترشح شده توسط اعصاب خودمختار عضله صاف استیل کولین و نوراپی نفرین هستند، اما هرگز توسط رشتههای عصبی مشابهی ترشح نمیشوند. استیل کولین یک ماده فرستنده تحریک کننده برای فیبرهای عضلانی صاف در برخی از اندامها است اما یک فرستنده بازدارنده برای عضلات صاف در سایر اندامها است. هنگامیکه استیل کولین فیبر عضلانی را تحریک میکند، نوراپی نفرین معمولاً آن را مهار میکند. برعکس، وقتی استیل کولین فیبر را مهار میکند، نوراپی نفرین معمولاً آن را تحریک میکند.
اما چرا این پاسخها متفاوت است؟ پاسخ این است که هم استیل کولین و هم نوراپی نفرین با اتصال به پروتئین گیرنده در سطح غشای سلول عضلانی، عضله صاف را تحریک یا مهار میکنند. برخی از پروتئینهای گیرنده گیرندههای تحریک کننده هستند، در حالی که برخی دیگر گیرندههای مهاری هستند. بنابراین، نوع گیرنده تعیین میکند که آیا ماهیچه صاف مهار یا برانگیخته شده است و همچنین تعیین میکند که کدام یک از دو فرستنده، استیل کولین یا نوراپی نفرین، در ایجاد تحریک یا مهار مؤثر است. این گیرندهها با جزئیات بیشتری در فصل ۶۰ در رابطه با عملکرد سیستم عصبی خودمختار مورد بحث قرار میگیرند.
پتانسیلهای غشایی و پتانسیلهای عمل در عضلات صاف
پتانسیلهای غشایی در عضلات صاف
ولتاژ کمیپتانسیل غشایی عضله صاف به وضعیت لحظه ای عضله بستگی دارد. در حالت استراحت طبیعی، پتانسیل درون سلولی معمولاً حدود ۵۰- تا ۶۰- میلی ولت است که حدود ۳۰ میلی ولت کمتر از ماهیچههای اسکلتی منفی است.
پتانسیلهای عمل در عضلات صاف یکپارچه
پتانسیل عمل در ماهیچه صاف واحد (مانند عضله احشایی) به همان شکلی که در عضله اسکلتی رخ میدهد رخ میدهد. همانطور که در بخش بعدی توضیح داده شد، معمولاً در اکثر انواع ماهیچههای صاف چند واحدی رخ نمیدهند.
پتانسیل عمل عضله صاف احشایی به یکی از دو شکل رخ میدهد: (۱) پتانسیل سنبله یا (۲) پتانسیل عمل با پلاتو.
پتانسیلهای سنبله
پتانسیل عمل سنبله معمولی، مانند آنهایی که در عضله اسکلتی مشاهده میشود، در اکثر انواع عضله صاف واحد رخ میدهد. مدت زمان این نوع پتانسیل عمل ۱۰ تا ۵۰ میلی ثانیه است، همانطور که در شکل ۸-۵ A نشان داده شده است. چنین پتانسیلهای عملی را میتوان به روشهای مختلفی برانگیخت، به عنوان مثال، با تحریک الکتریکی، با عمل هورمونها بر روی ماهیچه صاف، با عمل مواد فرستنده از رشتههای عصبی، با کشش، یا در نتیجه تولید خود به خود در عضله. خود فیبر، همانطور که در ادامه بحث شد.
 شکل ۸-۵ الف، پتانسیل عمل عضله صاف معمولی (پتانسیل سنبله) که توسط یک محرک خارجی ایجاد میشود. ب، پتانسیلهای پیک تکراری، که توسط امواج الکتریکی ریتمیک آهسته که به طور خود به خود در عضله صاف دیواره روده ایجاد میشود، ایجاد میشود. ج، پتانسیل عمل با یک پلاتو، ثبت شده از فیبر عضلانی صاف رحم.
شکل ۸-۵ الف، پتانسیل عمل عضله صاف معمولی (پتانسیل سنبله) که توسط یک محرک خارجی ایجاد میشود. ب، پتانسیلهای پیک تکراری، که توسط امواج الکتریکی ریتمیک آهسته که به طور خود به خود در عضله صاف دیواره روده ایجاد میشود، ایجاد میشود. ج، پتانسیل عمل با یک پلاتو، ثبت شده از فیبر عضلانی صاف رحم.
پتانسیلهای عمل با Plateaus
شکل ۸-۵ C پتانسیل عمل عضله صاف را با فلات نشان میدهد. شروع این پتانسیل عمل شبیه به پتانسیل سنبله معمولی است. با این حال، به جای رپولاریزاسیون سریع غشای فیبر عضلانی، رپلاریزاسیون چند صد تا ۱۰۰۰ میلی ثانیه (۱ ثانیه) به تاخیر میافتد. اهمیت فلات در این است که میتواند دلیل انقباض طولانی مدتی باشد که در برخی از انواع ماهیچههای صاف مانند حالب، رحم تحت برخی شرایط و انواع خاصی از عضلات صاف عروق رخ میدهد. (همچنین، این نوع پتانسیل عملی است که در فیبرهای عضلانی قلبی که دوره انقباض طولانی دارند دیده میشود، همانطور که در فصلهای ۹ و ۱۰ مورد بحث قرار گرفت.)
کانالهای کلسیم در ایجاد پتانسیل عمل ماهیچههای صاف مهم هستند
غشای سلولی عضله صاف کانالهای کلسیمیبسیار بیشتری نسبت به ماهیچههای اسکلتی دارد، اما کانالهای سدیمیبا ولتاژ کمیدارند. بنابراین، سدیم در تولید پتانسیل عمل در اکثر عضلات صاف شرکت کمیدارد. در عوض، جریان یونهای کلسیم به داخل فیبر عمدتاً مسئول پتانسیل عمل است. این امر به همان روشی که برای کانالهای سدیم در رشتههای عصبی و در رشتههای عضلانی اسکلتی رخ میدهد، اتفاق میافتد. با این حال، کانالهای کلسیم چندین برابر کندتر از کانالهای سدیم باز میشوند و همچنین برای مدت طولانی تری باز میمانند. این به میزان زیادی پتانسیل عمل طولانی مدت فلات برخی از فیبرهای عضلانی صاف را نشان میدهد.
یکی دیگر از ویژگیهای مهم ورود یون کلسیم به سلولها در طی پتانسیل عمل این است که یونهای کلسیم مستقیماً بر روی مکانیسم انقباض ماهیچه صاف عمل کرده و باعث انقباض میشوند. بنابراین، کلسیم دو وظیفه را همزمان انجام میدهد.
پتانسیلهای موج آهسته در عضلات صاف یکپارچه میتواند منجر به تولید خودبهخودی پتانسیلهای عمل شود.
برخی از ماهیچههای صاف خود تحریک کننده هستند. یعنی پتانسیل عمل در درون خود سلولهای عضله صاف بدون محرک بیرونی ایجاد میشود. این اغلب با یک ریتم اصلی موج آهسته پتانسیل غشا همراه است. یک موج آهسته معمولی در عضله صاف احشایی روده در شکل ۸-۵ B نشان داده شده است. خود موج آهسته پتانسیل عمل نیست. یعنی این یک فرآیند خودبازسازی نیست که به تدریج روی غشاهای فیبرهای عضلانی پخش شود. در عوض، این خاصیت محلی فیبرهای عضلانی صاف است که توده عضلانی را تشکیل میدهند.
علت ریتم امواج آهسته ناشناخته است. یک پیشنهاد این است که امواج آهسته ناشی از اپیلاسیون و کاهش پمپاژ یونهای مثبت (احتمالاً یونهای سدیم) به بیرون از طریق غشای فیبر عضلانی است. یعنی وقتی سدیم به سرعت پمپ میشود پتانسیل غشا منفی تر میشود و وقتی پمپ سدیم کمتر فعال میشود منفی تر میشود. پیشنهاد دیگر این است که رسانایی کانالهای یونی به صورت ریتمیک افزایش و کاهش مییابد.
اهمیت امواج آهسته در این است که وقتی به اندازه کافی قوی باشند، میتوانند پتانسیلهای عمل را آغاز کنند. امواج آهسته خود نمیتوانند باعث انقباض عضلانی شوند. با این حال، هنگامیکه اوج پتانسیل موج آهسته منفی در داخل غشای سلولی در جهت مثبت از ۶۰- به حدود ۳۵- میلی ولت (آستانه تقریبی برای برانگیختن پتانسیلهای عمل در بیشتر عضلات صاف احشایی) افزایش مییابد، یک پتانسیل عمل ایجاد میشود و در آن گسترش مییابد. توده عضلانی و انقباض رخ میدهد. شکل ۸-۵ B این اثر را نشان میدهد و نشان میدهد که در هر پیک موج آهسته، یک یا چند پتانسیل عمل رخ میدهد. این توالیهای تکراری پتانسیلهای عمل باعث انقباض ریتمیک توده عضلانی صاف میشود. بنابراین امواج آهسته را امواج ضربان ساز مینامند. که در در فصل ۶۲، میبینیم که این نوع فعالیت ضربان ساز، انقباضات ریتمیک روده را کنترل میکند.
تحریک عضله صاف احشایی با کشش عضلانی
وقتی عضله صاف احشایی (یونیتی) به اندازه کافی کشیده شود، معمولاً پتانسیلهای عمل خود به خودی ایجاد میشود. آنها از ترکیبی از (۱) پتانسیل موج آهسته معمولی و (۲) کاهش منفی کلی پتانسیل غشا ناشی از خود کشش ناشی میشوند. این پاسخ به کشش باعث میشود که دیواره روده، زمانی که بیش از حد کشیده میشود، به طور خودکار و ریتمیک منقبض شود. به عنوان مثال، هنگامیکه روده بیش از حد از محتویات روده پر میشود، انقباضات خودکار موضعی اغلب امواج پریستالتیک ایجاد میکند که محتویات را از روده پر شده دور میکند، معمولاً در جهت مقعد.
دپلاریزاسیون عضله صاف چند واحدی بدون پتانسیل عمل
فیبرهای عضلانی صاف عضله صاف چند واحدی (مانند عضله عنبیه چشم یا عضله پیلوئرکتور هر مو) معمولاً در پاسخ به محرکهای عصبی منقبض میشوند. پایانههای عصبی در مورد برخی از عضلات صاف چند واحدی استیل کولین و در مورد برخی دیگر نوراپی نفرین ترشح میکنند. در هر دو مورد، مواد فرستنده باعث دپلاریزاسیون غشای عضله صاف میشوند و این به نوبه خود باعث انقباض میشود. پتانسیل عمل معمولاً توسعه نمییابد. دلیل آن این است که الیاف برای تولید پتانسیل عمل بسیار کوچک هستند. (زمانی که پتانسیل عمل در عضله صاف واحد احشایی برانگیخته میشود، ۳۰ تا ۴۰ فیبر عضلانی صاف باید به طور همزمان دپلاریزه شوند قبل از اینکه پتانسیل عمل خود تکثیر پیدا کند.) با این حال در سلولهای ماهیچه صاف کوچک، حتی بدون پتانسیل عمل، دپلاریزاسیون موضعی (به نام پتانسیل اتصالی) ناشی از خود ماده فرستنده عصبی گسترش مییابد. به صورت الکتروتونیک روی کل فیبر قرار میگیرد و تمام چیزی است که برای ایجاد انقباض عضلانی لازم است.
تأثیر عوامل و هورمونهای بافتی موضعی در ایجاد انقباض عضلات صاف بدون پتانسیل عمل
احتمالاً نیمیاز تمام انقباضات ماهیچه صاف توسط عوامل محرکی شروع میشود که مستقیماً بر روی دستگاه انقباض ماهیچه صاف و بدون پتانسیل عمل تأثیر میگذارد. دو نوع از عوامل تحریک کننده بالقوه غیر عصبی و غیرعملی اغلب درگیر هستند (۱) عوامل شیمیایی بافت محلی و (۲) هورمونهای مختلف.
انقباض عضلات صاف در پاسخ به عوامل شیمیایی بافت محلی
در فصل ۱۷، کنترل انقباض شریانها، متا شریانها و اسفنکترهای پیش مویرگی را مورد بحث قرار میدهیم. کوچکترین این رگها منبع عصبی کمیدارند یا اصلاً وجود ندارند. با این حال عضله صاف بسیار انقباض است و به سرعت به تغییرات شرایط شیمیایی موضعی در مایع بینابینی اطراف واکنش نشان میدهد.
در حالت استراحت طبیعی، بسیاری از این رگهای خونی کوچک منقبض میمانند. اما زمانی که جریان خون اضافی به بافت ضروری باشد، عوامل متعددی میتوانند دیواره رگ را شل کنند و در نتیجه جریان را افزایش دهند. به این ترتیب، یک سیستم کنترل بازخورد محلی قدرتمند، جریان خون را در ناحیه بافت محلی کنترل میکند. برخی از عوامل کنترل خاص به شرح زیر است:
۱. کمبود اکسیژن در بافتهای موضعی باعث شل شدن عضلات صاف و در نتیجه اتساع عروق میشود.
۲. دی اکسید کربن اضافی باعث اتساع عروق میشود.
۳. افزایش غلظت یون هیدروژن باعث اتساع عروق میشود.
آدنوزین، اسید لاکتیک، افزایش یونهای پتاسیم، کاهش غلظت یون کلسیم و افزایش دمای بدن همگی میتوانند باعث اتساع موضعی عروق شوند.
اثرات هورمونها بر انقباض عضلات صاف
بسیاری از هورمونهای در گردش خون تا حدی بر انقباض عضلات صاف تأثیر میگذارند و برخی نیز تأثیرات عمیقی دارند. از جمله مهمترین آنها میتوان به نوراپی نفرین، اپی نفرین، استیل کولین، آنژیوتانسین، اندوتلین، وازوپرسین، اکسی توسین، سروتونین و هیستامین اشاره کرد.
زمانی که غشای سلولی عضلانی حاوی گیرندههای تحریک کننده هورمون مربوطه باشد، یک هورمون باعث انقباض ماهیچه صاف میشود. برعکس، اگر غشاء حاوی گیرندههای بازدارنده هورمون باشد تا گیرندههای تحریک کننده، هورمون باعث مهار میشود.
مکانیسمهای تحریک یا مهار عضلات صاف توسط هورمونها یا عوامل بافتی محلی
برخی از گیرندههای هورمونی در غشای عضله صاف، کانالهای یونی سدیم یا کلسیم را باز میکنند و غشا را دپولاریزه میکنند، مانند پس از تحریک عصبی. گاهی اوقات پتانسیلهای عمل به نتیجه میرسند، یا پتانسیلهای عملی که در حال حاضر رخ میدهند ممکن است تقویت شوند. در موارد دیگر، دپلاریزاسیون بدون پتانسیل عمل رخ میدهد و این دپلاریزاسیون اجازه ورود یون کلسیم به سلول را میدهد که انقباض را تقویت میکند.
در مقابل، مهار زمانی اتفاق میافتد که هورمون (یا سایر عوامل بافتی) کانالهای سدیم و کلسیم را ببندد تا از ورود این یونهای مثبت جلوگیری کند. همچنین اگر کانالهای پتاسیمیکه معمولاً بسته هستند باز شوند و یونهای مثبت پتاسیم به بیرون از سلول منتشر شوند، مهار نیز رخ میدهد . هر دوی این اقدامات درجه منفی بودن را در داخل سلول عضلانی افزایش میدهند، حالتی به نام هایپرپلاریزاسیون که به شدت انقباض عضلانی را مهار میکند.
گاهی اوقات انقباض یا مهار ماهیچه صاف توسط هورمونها بدون ایجاد تغییر مستقیم در پتانسیل غشاء آغاز میشود. در این موارد، هورمون ممکن است گیرنده غشایی را فعال کند که هیچ کانال یونی را باز نمیکند، اما در عوض باعث تغییر درونی فیبر عضلانی، مانند آزاد شدن یونهای کلسیم از شبکه سارکوپلاسمیداخل سلولی میشود. سپس کلسیم باعث انقباض میشود. برای مهار انقباض، مکانیسمهای گیرنده دیگری شناخته شدهاند که آنزیم آدنیلات سیکلاز یا گوانیلات سیکلاز را در غشای سلولی فعال میکنند. بخشهایی از گیرندههایی که به داخل سلولها بیرون زدهاند به این آنزیمها جفت میشوند و باعث تشکیل آدنوزین مونوفسفات حلقوی (cAMP) میشوند.گوانوزین مونوفسفات حلقوی (cGMP)، به اصطلاح پیام رسان دوم. cAMP یا cGMP اثرات زیادی دارد که یکی از آنها تغییر درجه فسفوریلاسیون چندین آنزیم است که به طور غیر مستقیم انقباض را مهار میکنند. پمپی که یونهای کلسیم را از سارکوپلاسم به داخل شبکه سارکوپلاسمیمنتقل میکند، و همچنین پمپ غشای سلولی که یونهای کلسیم را از خود سلول خارج میکند، فعال میشود. این اثرات باعث کاهش غلظت یون کلسیم در سارکوپلاسم میشود و در نتیجه انقباض را مهار میکند.
ماهیچههای صاف تنوع قابل توجهی در نحوه شروع انقباض یا آرامش در پاسخ به هورمونها، انتقال دهندههای عصبی و سایر مواد مختلف دارند. در برخی موارد، یک ماده ممکن است باعث شل شدن یا انقباض عضلات صاف در مکانهای مختلف شود. به عنوان مثال، نوراپی نفرین انقباض ماهیچه صاف روده را مهار میکند اما انقباض ماهیچه صاف در رگهای خونی را تحریک میکند.
منبع یونهای کلسیم که باعث انقباض از طریق غشای سلولی و از شبکه سارکوپلاسمیمیشوند.
اگرچه فرآیند انقباض در عضلات صاف، مانند ماهیچههای اسکلتی، توسط یونهای کلسیم فعال میشود، منبع یونهای کلسیم متفاوت است. یک تفاوت مهم این است که شبکه سارکوپلاسمی، که تقریباً تمام یونهای کلسیم را برای انقباض ماهیچههای اسکلتی فراهم میکند، در اکثر عضلات صاف تنها اندکی توسعه یافته است. در عوض، بیشتر یونهای کلسیم که باعث انقباض میشوند، از مایع خارج سلولی در زمان پتانسیل عمل یا سایر محرکها وارد سلول عضلانی میشوند. یعنی غلظت یونهای کلسیم در مایع خارج سلولی بیشتر از ۱۰-۳ مولر است، در مقایسه با کمتر از ۱۰-۷. مولر داخل سلول عضله صاف؛ این باعث انتشار سریع یونهای کلسیم به داخل سلول از مایع خارج سلولی با باز شدن کانالهای کلسیم میشود. زمان لازم برای این انتشار به طور متوسط ۲۰۰ تا ۳۰۰ میلی ثانیه است و دوره نهفته قبل از شروع انقباض نامیده میشود. این دوره نهفته برای ماهیچه صاف حدود ۵۰ برابر انقباض عضلات اسکلتی است.
نقش شبکه سارکوپلاسمیعضلات صاف
شکل ۸-۶ چند لوله سارکوپلاسمیکمیتوسعه یافته را نشان میدهد که در نزدیکی غشای سلولی در برخی از سلولهای عضله صاف بزرگتر قرار دارند. فرورفتگیهای کوچک غشای سلولی به نام caveolae به سطوح این لولهها میرسد. caveolae یک آنالوگ ابتدایی از سیستم لوله عرضی عضله اسکلتی را نشان میدهد. هنگامیکه یک پتانسیل عمل به داخل caveolae منتقل میشود، اعتقاد بر این است که این امر باعث تحریک آزاد شدن یون کلسیم از لولههای سارکوپلاسمینزدیک میشود، به همان روشی که پتانسیل عمل در لولههای عرضی عضله اسکلتی باعث آزاد شدن یونهای کلسیم از لولههای سارکوپلاسمیطولی عضله اسکلتی میشود. به طور کلی، هرچه شبکه سارکوپلاسمیدر فیبر ماهیچه صاف گسترده تر باشد، سریعتر منقبض میشود.
 شکل ۸-۶ لولههای سارکوپلاسمی در یک فیبر عضلانی صاف بزرگ که رابطه آنها را با فرورفتگی در غشای سلولی به نام caveolae نشان میدهد.
شکل ۸-۶ لولههای سارکوپلاسمی در یک فیبر عضلانی صاف بزرگ که رابطه آنها را با فرورفتگی در غشای سلولی به نام caveolae نشان میدهد.
انقباض عضلات صاف به غلظت یون کلسیم خارج سلولی بستگی دارد
اگرچه تغییر غلظت یون کلسیم مایع خارج سلولی از حالت طبیعی تأثیر کمیبر نیروی انقباض عضله اسکلتی دارد، اما این برای اکثر عضلات صاف صادق نیست. هنگامیکه غلظت یون کلسیم مایع خارج سلولی به حدود ۱/۳ تا ۱/۱۰ طبیعی کاهش مییابد، انقباض عضلات صاف معمولاً متوقف میشود. بنابراین، نیروی انقباض عضله صاف معمولاً به شدت به غلظت یون کلسیم مایع خارج سلولی وابسته است.
پمپ کلسیم برای ایجاد آرامش عضلات صاف مورد نیاز است
برای ایجاد آرامش عضله صاف پس از انقباض، یونهای کلسیم باید از مایعات داخل سلولی خارج شوند. این حذف توسط یک پمپ کلسیم انجام میشود که یونهای کلسیم را از فیبر ماهیچه صاف به داخل مایع خارج سلولی یا در صورت وجود به شبکه سارکوپلاسمیپمپ میکند. این پمپ در مقایسه با پمپ شبکه سارکوپلاسمیسریع الاثر در عضله اسکلتی کند عمل میکند. بنابراین، یک انقباض عضله صاف اغلب برای چند ثانیه طول میکشد تا صدم تا دهم ثانیه، همانطور که برای ماهیچههای اسکلتی اتفاق میافتد.
کتاب درسی فیزیولوژی پزشکی گایتون وهال، ویرایش دوازدهم فصل ۸
کلیک کنید «بیبلیوگرافی: فهرست کتب مربوطه»
independent contractions in smooth muscles. Am J Med. 2003;115(Suppl 3A):24S.
Hilgers R.H., Webb R.C. Molecular aspects of arterial smooth muscle contraction: focus on Rho. Exp Biol Med (Maywood). 2005;230:829.
House S.J., Potier M., Bisaillon J., Singer H.A., Trebak M. The non-excitable smooth muscle: calcium signaling and phenotypic switching during vascular disease. Pflugers Arch. 2008;456:769.
Huizinga J.D., Lammers W.J. Gut peristalsis is governed by a multitude of cooperating mechanisms. Am J Physiol Gastrointest Liver Physiol. 2009;296:G1.
Kuriyama H., Kitamura K., Itoh T., Inoue R. Physiological features of visceral smooth muscle cells, with special reference to receptors and ion channels. Physiol Rev. 1998;78:811.
Morgan K.G., Gangopadhyay S.S. Cross-bridge regulation by thin filament-associated proteins. J Appl Physiol. 2001;91:953.
Somlyo A.P., Somlyo A.V. Ca۲+ sensitivity of smooth muscle and nonmuscle myosin II: modulated by G proteins, kinases, and myosin phosphatase. Physiol Rev. 2003;83:1325.
Stephens N.L. Airway smooth muscle. Lung. 2001;179:333.
Touyz R.M. Transient receptor potential melastatin 6 and 7 channels, magnesium transport, and vascular biology: implications in hypertension. Am J Physiol Heart Circ Physiol. 2008;294:H1103.
Walker J.S., Wingard C.J., Murphy R.A. Energetics of crossbridge phosphorylation and contraction in vascular smooth muscle. Hypertension. 1994;23:1106.
Wamhoff B.R., Bowles D.K., Owens G.K. Excitation-transcription coupling in arterial smooth muscle. Circ Res. 2006;98:868.
Webb R.C. Smooth muscle contraction and relaxation. Adv Physiol Educ. 2003;27:201.
