فیزیولوژی پزشکی گانونگ؛ رفلکس و کنترل ارادی وضعیت و حرکت

دعای مطالعه [ نمایش ]
بِسْمِ الله الرَّحْمنِ الرَّحیمِ
اَللّهُمَّ اَخْرِجْنى مِنْ ظُلُماتِ الْوَهْمِ
خدايا مرا بيرون آور از تاريكىهاى وهم،
وَ اَكْرِمْنى بِنُورِ الْفَهْمِ
و به نور فهم گرامى ام بدار،
اَللّهُمَّ افْتَحْ عَلَيْنا اَبْوابَ رَحْمَتِكَ
خدايا درهاى رحمتت را به روى ما بگشا،
وَانْشُرْ عَلَيْنا خَزائِنَ عُلُومِكَ بِرَحْمَتِكَ يا اَرْحَمَ الرّاحِمينَ
و خزانههاى علومت را بر ما باز كن به امید رحمتت اى مهربانترين مهربانان.
کتاب «فیزیولوژی پزشکی گانونگ» بهعنوان یکی از جامعترین و معتبرترین منابع در حوزه علوم پزشکی، همچنان مرجع کلیدی برای درک عملکرد پیچیده بدن انسان است. این اثر با تکیه بر تازهترین پژوهشها و توضیحات دقیق از سازوکارهای فیزیولوژیک، پلی میان علوم پایه پزشکی و کاربردهای بالینی ایجاد میکند و نقشی بیبدیل در آموزش، پژوهش و ارتقای دانش سلامت ایفا مینماید.
ترجمه دقیق و علمی این شاهکار توسط برند علمی آیندهنگاران مغز به مدیریت داریوش طاهری، دسترسی فارسیزبانان به مرزهای نوین دانش فیزیولوژی را ممکن ساخته و رسالتی علمی برای ارتقای آموزش پزشکی، فهم عمیقتر سازوکارهای بدن و توسعه روشهای نوین در حوزه سلامت فراهم آورده است.
» کتاب فیزیولوژی پزشکی گانونگ
» » فصل ۱۲: رفلکس و کنترل ارادی وضعیت و حرکت
در حال ویرایش
» Ganong’s Review of Medical Physiology
»» CHAPTER 12: Reflex and Voluntary Control of Posture & Movement
OBJECTIVES
After studying his chapter you should be able to:
• Describe the basic elements of a reflex pathway.
• Identify the components, function, and afferent nerve fibers of the muscle spindle.
• Explain the neuronal response initiated by striking the patellar tendon (patellar tendon or knee jerk reflex) that leads to contraction of a muscle.
• Explain how the activity of y-motor neurons alters the response to muscle stretch.
• Describe the role of Golgi tendon organs in the control of skeletal muscle.
• Define physiologic tremor, clonus, and muscle tone.
• Identify the components and function of the withdrawal reflex pathway.
• Define spinal shock and describe the initial and long-term changes in spinal reflexes that follow spinal cord injury.
• Describe how skilled movements are planned and carried out.
• Contrast the organization of the central pathways involved in the control of axial (posture) and distal (skilled movement, fine motor movements)muscles.
• Describe the clinical tests and findings that distinguish between upper and lower motor neuron disorders, including the Babinski sign and clonus.
• Identify the pathophysiology and characteristics of cerebral palsy, decerebrate rigidity, and decorticate rigidity.
• Identify the components of the basal ganglia and the pathways that interconnect them, along with the neurotransmitters in each pathway.
• Explain the pathophysiology and symptoms of Parkinson disease, Huntington disease, and other pathologies of the basal ganglia pathways.
• Discuss the functions of the cerebellum and the neurologic abnormalities produced by diseases of this part of the brain.
اهداف
پس از مطالعه این فصل، باید بتوانید:
• عناصر اساسی یک مسیر رفلکس را شرح دهید.
• اجزاء، عملکرد و رشتههای عصبی آوران دوک عضلانی را شناسایی کنید.
• پاسخ عصبی که با ضربه زدن به تاندون کشکک (تاندون کشکک یا رفلکس تکان دادن زانو) آغاز میشود را توضیح دهید که منجر به انقباض عضله میشود.
• توضیح دهید که چگونه فعالیت نورونهای حرکتی y پاسخ به کشش عضلانی را تغییر میدهد.
• نقش اندامهای تاندون گلژی را در کنترل ماهیچههای اسکلتی شرح دهید.
• ترمور فیزیولوژیک، کلونوس و تون عضلانی را تعریف کنید.
• اجزاء و عملکرد مسیر رفلکس خروج را شناسایی کنید.
• شوک نخاعی را تعریف کنید و تغییرات اولیه و طولانی مدت در رفلکسهای نخاعی که به دنبال آسیب نخاعی ایجاد میشود را شرح دهید.
• نحوه برنامه ریزی و اجرای حرکات ماهرانه را شرح دهید.
• سازماندهی مسیرهای مرکزی درگیر در کنترل عضلات محوری (حالت بدن) و دیستال (حرکت ماهرانه، حرکات حرکتی ظریف) را با هم مقایسه کنید.
• آزمایشهای بالینی و یافتههایی را که بین اختلالات نورون حرکتی فوقانی و تحتانی، از جمله علامت بابینسکی و کلونوس تمایز میدهند، توضیح دهید.
• شناسایی پاتوفیزیولوژی و ویژگیهای فلج مغزی، سفتی decerebrate، و سفتی decorticate.
• اجزای عقدههای قاعده ای و مسیرهایی که آنها را به هم متصل میکنند، همراه با انتقال دهندههای عصبی در هر مسیر را شناسایی کنید.
• پاتوفیزیولوژی و علائم بیماری پارکینسون، بیماریهانتینگتون و سایر آسیب شناسیهای مسیرهای عقدههای قاعده ای را توضیح دهید.
• در مورد عملکرد مخچه و ناهنجاریهای عصبی ناشی از بیماریهای این قسمت از مغز بحث کنید.

INTRODUCTION
Somatic motor function depends ultimately on the activity of the spinal motor neurons and homologous neurons in the motor nuclei of the cranial nerves. These neurons, the final common pathways to skeletal muscle, are bombarded by impulses from an immense array of descending pathways, other spinal neurons, and peripheral afferents. Some of these inputs end directly on a-motor neurons, but many exert their effects via interneurons or via y-motor neurons to the muscle spindles and back through the Ia afferent fibers to the spinal cord. It is the integrated activity of these multiple inputs from spinal, brainstem, midbrain, and cortical levels that regulates the posture of the body and makes coordinated movement possible.
مقدمه
عملکرد حرکتی جسمیدر نهایت به فعالیت نورونهای حرکتی ستون فقرات و نورونهای همولوگ در هستههای حرکتی اعصاب جمجمه بستگی دارد. این نورونها، آخرین مسیرهای مشترک به ماهیچههای اسکلتی، توسط تکانههایی از مجموعه عظیمیاز مسیرهای نزولی، سایر نورونهای نخاعی و آورانهای محیطی بمباران میشوند. برخی از این ورودیها مستقیماً به نورونهای a-motor ختم میشوند، اما بسیاری از آنها تأثیرات خود را از طریق نورونهای بیناعصاب یا از طریق نورونهای y-motor به دوکهای عضلانی و از طریق رشتههای آوران Ia به طناب نخاعی اعمال میکنند. این فعالیت یکپارچه این ورودیهای چندگانه از سطوح ستون فقرات، ساقه مغز، مغز میانی و قشر مغز است که وضعیت بدن را تنظیم میکند و حرکت هماهنگ را ممکن میکند.
The inputs converging on motor neurons have three major functions: to induce voluntary activity, to adjust body posture, and to make movements smooth and precise. The patterns of voluntary activity are planned within the brain, and the commands are sent to the muscles primarily via the corticospinal and corticobulbar systems. Posture is continually adjusted not only before but also during movement by information carried in descending brainstem pathways and peripheral afferents. Movement is smoothed and coordinated by the medial and intermediate portions of the cerebellum (spinocerebellum) and its connections. The basal ganglia and the lateral portions of the cerebellum (cerebrocerebellum) are part of a feedback circuit to the premotor and motor cortex that is concerned with planning and organizing voluntary movement. Chapter 11 introduced somatomotor control by describing the role of the vestibular system in control of balance and equilibrium. This chapter considers two types of motor output: reflex (involuntary) and voluntary. A subdivision of reflex responses includes some rhythmic movements such as swallowing, chewing, scratching, and walking, which are largely involuntary but are subject to voluntary adjustment and control.
ورودیهایی که روی نورونهای حرکتی همگرا میشوند، سه عملکرد عمده دارند: القای فعالیت ارادی، تنظیم وضعیت بدن، و صاف و دقیق کردن حرکات. الگوهای فعالیت ارادی در مغز برنامه ریزی میشود و دستورات عمدتاً از طریق سیستمهای قشر نخاعی و کورتیکوبولبار به عضلات ارسال میشود. وضعیت بدن به طور مداوم نه تنها قبل از حرکت بلکه در حین حرکت توسط اطلاعاتی که در مسیرهای نزولی ساقه مغز و آورانهای محیطی حمل میشود تنظیم میشود. حرکت توسط بخشهای میانی و میانی مخچه (اسپینوسربلوم) و اتصالات آن هموار و هماهنگ میشود. عقدههای قاعده ای و بخشهای جانبی مخچه (مخچه) بخشی از یک مدار بازخورد به قشر پیش حرکتی و حرکتی هستند که به برنامه ریزی و سازماندهی حرکت ارادی مربوط میشود. فصل 11 با تشریح نقش سیستم دهلیزی در کنترل تعادل و تعادل، کنترل جسمیحرکتی را معرفی کرد. این فصل دو نوع خروجی موتور را در نظر میگیرد: رفلکس (غیر ارادی) و ارادی. یک زیربخش از پاسخهای رفلکس شامل برخی حرکات موزون مانند بلع، جویدن، خراشیدن و راه رفتن است که عمدتاً غیرارادی هستند اما در معرض تنظیم و کنترل ارادی هستند.
GENERAL PROPERTIES OF REFLEXES
The basic unit of integrated reflex activity is the reflex arc that consists of a sense organ, an afferent neuron, one or more synapses within a central integrating station, an efferent neuron, and an effector. The afferent neurons enter via the dorsal roots or cranial nerves and have their cell bodies in dorsal root ganglia or homologous cranial nerve ganglia. The efferent fibers leave via the ventral roots or motor cranial nerves.
ویژگیهای عمومیبازتابها
واحد اصلی فعالیت رفلکس یکپارچه، قوس بازتابی است که از یک اندام حسی، یک نورون آوران، یک یا چند سیناپس در یک ایستگاه مرکزی یکپارچه، یک نورون وابران و یک عامل تشکیل شده است. نورونهای آوران از طریق ریشههای پشتی یا اعصاب جمجمه ای وارد میشوند و بدن سلولی خود را در گانگلیونهای ریشه پشتی یا عقدههای عصبی همولوگ جمجمه ای قرار میدهند. فیبرهای وابران از طریق ریشههای شکمییا اعصاب جمجمه حرکتی خارج میشوند.
Activity in the reflex arc starts in a sensory receptor with a receptor potential (see Chapter 8) whose magnitude is proportional to the strength of the stimulus (Figure 12-1). This generates all-or-none action potentials in the afferent nerve, the number of action potentials being proportional to the size of the receptor potential. In the central nervous system (CNS), the responses are again graded in terms of excitatory postsynaptic potentials (EPSPs) and inhibitory postsynaptic potentials (IPSPs) at the synaptic junctions (see Chapter 6). When the action potentials in the efferent motor nerve reach the effector, a graded response is induced. If the graded response is adequate to produce action potentials in the muscle, muscle contraction will ensue. The activity in the reflex arc is modified by the multiple inputs converging on the efferent neurons or at any synaptic station within the reflex arc.
فعالیت در قوس بازتابی در یک گیرنده حسی با پتانسیل گیرنده شروع میشود (به فصل 8 مراجعه کنید) که بزرگی آن متناسب با قدرت محرک است (شکل 12-1). این پتانسیلهای عمل همه یا هیچ در عصب آوران ایجاد میکند، تعداد پتانسیلهای عمل متناسب با اندازه پتانسیل گیرنده است. در سیستم عصبی مرکزی (CNS)، پاسخها دوباره بر حسب پتانسیلهای پس سیناپسی تحریکی (EPSPs) و پتانسیلهای پس سیناپسی مهاری (IPSPs) در اتصالات سیناپسی درجه بندی میشوند (به فصل 6 مراجعه کنید). هنگامیکه پتانسیلهای عمل در عصب حرکتی وابران به عامل میرسد، یک پاسخ درجه بندی شده القا میشود. اگر پاسخ درجه بندی شده برای تولید پتانسیل عمل در عضله کافی باشد، انقباض عضلانی به وجود میآید. فعالیت در قوس بازتابی توسط ورودیهای متعددی که روی نورونهای وابران یا در هر ایستگاه سیناپسی درون قوس بازتابی همگرا میشوند، اصلاح میشود.

FIGURE 12–1 The reflex arc. Note that at the receptor and in the CNS anonpropagated graded response occurs that is proportional to the magnitude of the stimulus. The response at the neuromuscular junction is also graded, though under normal conditions it is always large enough to produce a response in skeletal muscle. On the other hand, in the portions of the arc specialized for transmission (afferent and efferent nerve fibers, muscle membrane), the responses are all-or-none action potentials.
شکل 12-1 قوس بازتابی. توجه داشته باشید که در گیرنده و در CNS پاسخ درجه بندی نشده ای رخ میدهد که متناسب با بزرگی محرک است. پاسخ در اتصال عصبی عضلانی نیز درجه بندی میشود، اگرچه در شرایط عادی همیشه به اندازه کافی بزرگ است که پاسخی در عضله اسکلتی ایجاد کند. از سوی دیگر، در بخشهایی از قوس که مخصوص انتقال هستند (فیبرهای عصبی آوران و وابران، غشای عضلانی)، پاسخها پتانسیلهای عمل همه یا هیچ هستند.
Reflex activity is stereotyped and specific in that a particular stimulus elicits a particular response. The fact that reflex responses are stereotyped does not exclude the possibility of their being modified by experience. Reflexes are adaptable and can be modified to perform motor tasks and maintain balance. Descending inputs from higher brain regions play an important role in modulating and adapting spinal reflexes.
فعالیت بازتابی کلیشه ای و خاص است که یک محرک خاص پاسخ خاصی را برمیانگیزد. این واقعیت که پاسخهای بازتابی کلیشهای هستند، امکان تغییر آنها توسط تجربه را رد نمیکند. رفلکسها سازگار هستند و میتوان آنها را برای انجام وظایف حرکتی و حفظ تعادل تغییر داد. ورودیهای نزولی از مناطق بالاتر مغز نقش مهمیدر تعدیل و تطبیق رفلکسهای نخاعی دارند.
The a-motor neurons that supply the extrafusal fibers in skeletal muscles are the efferent side of many reflex arcs. All neural influences affecting muscular contraction ultimately funnel through them to the muscles, thus they are called the final common pathway. Numerous inputs converge on a-motor neurons; the surface of an average motor neuron and its dendrites accommodates about 10,000 synaptic knobs. At least five inputs go from the same spinal segment to a typical spinal motor neuron. In addition to these, there are excitatory and inhibitory inputs, generally relayed via interneurons, from other levels of the spinal cord and multiple long-descending tracts from the brain. All of these pathways converge on and determine the activity in the final common pathway.
نورونهای a-motor که فیبرهای خارج رحمیماهیچههای اسکلتی را تامین میکنند، سمت وابران بسیاری از کمانهای بازتابی هستند. تمام تأثیرات عصبی مؤثر بر انقباض عضلانی در نهایت از طریق آنها به عضلات منتقل میشود، بنابراین آنها را آخرین مسیر مشترک مینامند. ورودیهای متعدد روی نورونهای a-motor همگرا میشوند. سطح یک نورون حرکتی متوسط و دندریتهای آن حدود 10000 دستگیره سیناپسی را در خود جای میدهد. حداقل پنج ورودی از همان بخش ستون فقرات به یک نورون حرکتی نخاعی معمولی میرود. علاوه بر اینها، ورودیهای تحریککننده و بازدارندهای وجود دارد که عموماً از طریق نورونهای داخلی، از سطوح دیگر نخاع و چندین مسیر نزولی طولانی از مغز منتقل میشوند. همه این مسیرها در مسیر مشترک نهایی همگرا شده و فعالیت را تعیین میکنند.
MONOSYNAPTIC REFLEXES: THE STRETCH REFLEX
The simplest reflex arc is the one with a single synapse between the afferent and efferent neurons (i.e., monosynaptic reflexes). Reflex arcs in which interneurons are interposed between the afferent and efferent neurons are called polysynaptic reflexes. There can be anywhere from two to hundreds of synapses in a polysynaptic reflex arc.
رفلکسهای تک سیناپتیک: رفلکس کششی
سادهترین قوس بازتابی، قوسی است که دارای یک سیناپس بین نورونهای آوران و وابران است (یعنی رفلکسهای تک سیناپسی). کمانهای بازتابی که در آن نورونهای بین نورونهای آوران و وابران قرار میگیرند، رفلکسهای پلی سیناپسی نامیده میشوند. در یک قوس بازتابی پلی سیناپسی میتواند از دو تا صدها سیناپس وجود داشته باشد.
When a skeletal muscle with an intact nerve supply is stretched, it contracts. This response is called the stretch reflex or myotatic reflex. The stimulus that initiates this reflex is stretch of the muscle, and the response is contraction of the muscle being stretched. The sense organ is a small encapsulated spindle-like or fusiform-shaped structure called the muscle spindle, located within the fleshy part of the muscle. The impulses originating from the spindle are transmitted to the CNS by fast sensory fibers that pass directly to the motor neurons that supply the same muscle. The neurotransmitter at the central synapse is glutamate. The stretch reflex is the best known and studied monosynaptic reflex and is typified by the knee jerk reflex (Clinical Box 12-1).
هنگامیکه یک عضله اسکلتی با منبع عصبی دست نخورده کشیده میشود، منقبض میشود. این پاسخ را رفلکس کششی یا رفلکس میوتاتیک مینامند. محرکی که این رفلکس را آغاز میکند کشش عضله است و پاسخ آن انقباض عضله در حال کشیده شدن است. اندام حسی یک ساختار دوکیمانند یا دوکیشکل محصور شده به نام دوک عضلانی است که در قسمت گوشتی عضله قرار دارد. تکانههایی که از دوک منشا میگیرند توسط فیبرهای حسی سریع به CNS منتقل میشوند که مستقیماً به نورونهای حرکتی که همان عضله را تأمین میکنند، منتقل میشوند. انتقال دهنده عصبی در سیناپس مرکزی گلوتامات است. رفلکس کششی شناخته شده ترین و مطالعه شده ترین رفلکس تک سیناپسی است و با رفلکس حرکتی زانو مشخص میشود (باکس بالینی 12-1).
CLINICAL BOX 12 1
Knee Jerk Reflex
Tapping the patellar tendon elicits the knee jerk, a stretch reflex of the quadriceps femoris muscle, because the tap on the tendon stretches the muscle. A similar contraction is observed if the quadriceps is stretched manually. The knee jerk reflex is an example of a deep tendon reflex (DTR) in a neurologic exam and is graded on the following scale: 0 (absent), 1+ (hypoactive), 2+ (brisk, normal), 3+ (hyperactive without clonus), 4+ (hyperactive with mild clonus), and 5+ (hyperactive with sustained clonus). Absence of the knee jerk can signify an abnormality anywhere within the reflex arc, including the muscle spindle, the Ia afferent nerve fibers, or the motor neurons to the quadriceps muscle. The most common cause is a peripheral neuropathy from such things as diabetes, alcoholism, and toxins. A hyperactive reflex can signify an interruption of corticospinal and other descending pathways that suppress the activity in the reflex arc. DTR is not specific to an assessment of the knee jerk reflex; stretch reflexes can be elicited from most of the large muscles of the body. Tapping on the tendon of the triceps brachii, for example, causes an extensor response at the elbow as a result of reflex contraction of the triceps. Tapping on the Achilles tendon causes an ankle jerk due to reflex contraction of the gastrocnemius, and tapping on the side of the face causes a stretch reflex in the masseter. The spinal nerve involved in testing various DTRS is as follows: biceps (C5, C6 spinal nerve); triceps (C7 spinal nerve); patellar (L4 spinal nerve); and Achilles tendon (S1 spinal nerve).
جعبه بالینی 12. 1
رفلکس حرکتی زانو
ضربه زدن به تاندون کشکک باعث ایجاد تکان زانو میشود، یک رفلکس کششی عضله چهار سر ران، زیرا ضربه روی تاندون باعث کشیدگی عضله میشود. اگر عضله چهار سر ران به صورت دستی کشیده شود، انقباض مشابهی مشاهده میشود. رفلکس حرکتی زانو نمونه ای از رفلکس تاندون عمیق (DTR) در معاینه عصبی است و در مقیاس زیر درجه بندی میشود: 0 (غایب)، 1+ (کم فعال)، 2+ (تند، نرمال)، 3+ (بیش فعال بدون کلونوس)، 4+ (بیش فعال با کلونوس خفیف 5+ فعال)، و کلونوس فعال. فقدان تکان دادن زانو میتواند نشان دهنده یک ناهنجاری در هر جایی از قوس بازتابی باشد، از جمله دوک عضلانی، رشتههای عصبی آوران Ia یا نورونهای حرکتی عضله چهار سر ران. شایع ترین علت نوروپاتی محیطی ناشی از مواردی مانند دیابت، اعتیاد به الکل و سموم است. رفلکس بیش فعال میتواند به معنی قطع شدن مسیرهای قشر نخاعی و سایر مسیرهای نزولی باشد که فعالیت در قوس رفلکس را سرکوب میکند. DTR برای ارزیابی رفلکس تکان دادن زانو خاص نیست. رفلکسهای کششی را میتوان از اکثر عضلات بزرگ بدن استخراج کرد. ضربه زدن بر روی تاندون عضله سه سر بازویی، به عنوان مثال، در نتیجه انقباض رفلکس عضلات سه سر، باعث پاسخ اکستانسور در آرنج میشود. ضربه زدن به تاندون آشیل به دلیل انقباض رفلکس گاستروکنمیوس باعث تکان مچ پا میشود و ضربه زدن به پهلوی صورت باعث ایجاد رفلکس کششی در ماستر میشود. عصب نخاعی درگیر در آزمایش DTRSهای مختلف به شرح زیر است: عصب دوسر (C5، C6 عصب نخاعی). سه سر (اعصاب نخاعی C7)؛ کشکک (عصب نخاعی L4)؛ و تاندون آشیل (عصب نخاعی S1).
STRUCTURE OF MUSCLE SPINDLES
Each muscle spindle has three essential elements: (1) a group of specialized intrafusal muscle fibers with contractile polar ends and a noncontractile center, (2) large diameter myelinated afferent nerves (types Ia and II) originating in the central portion of the intrafusal fibers, and (3) small diameter myelinated efferent nerves supplying the polar contractile regions of the intrafusal fibers (Figure 12-2A). It is important to understand the relationship of these elements to each other and to the muscle itself to appreciate the role of this sense organ in signaling changes in the length of the muscle in which it is located. Changes in muscle length are associated with changes in joint angle; thus, muscle spindles provide information on position (ie, proprioception).
ساختار دوکهای عضلانی
هر دوک عضلانی سه عنصر اساسی دارد: (1) گروهی از فیبرهای عضلانی داخل دوکی تخصصی با انتهای قطبی انقباضی و یک مرکز غیر انقباضی، (2) اعصاب آوران میلیندار با قطر بزرگ (انواع Ia و II) که از قسمت مرکزی فیبرهای داخل دوکی منشأ میگیرند، و (3) اعصاب وابران میلیندار با قطر کوچک که نواحی انقباضی قطبی فیبرهای داخل دوکی را تغذیه میکنند (شکل 12-2A). درک رابطه این عناصر با یکدیگر و با خود عضله برای درک نقش این اندام حسی در سیگنالدهی تغییرات طول عضلهای که در آن قرار دارد، مهم است. تغییرات طول عضله با تغییرات زاویه مفصل مرتبط است. بنابراین، دوکهای عضلانی اطلاعاتی در مورد موقعیت (یعنی حس عمقی) ارائه میدهند.


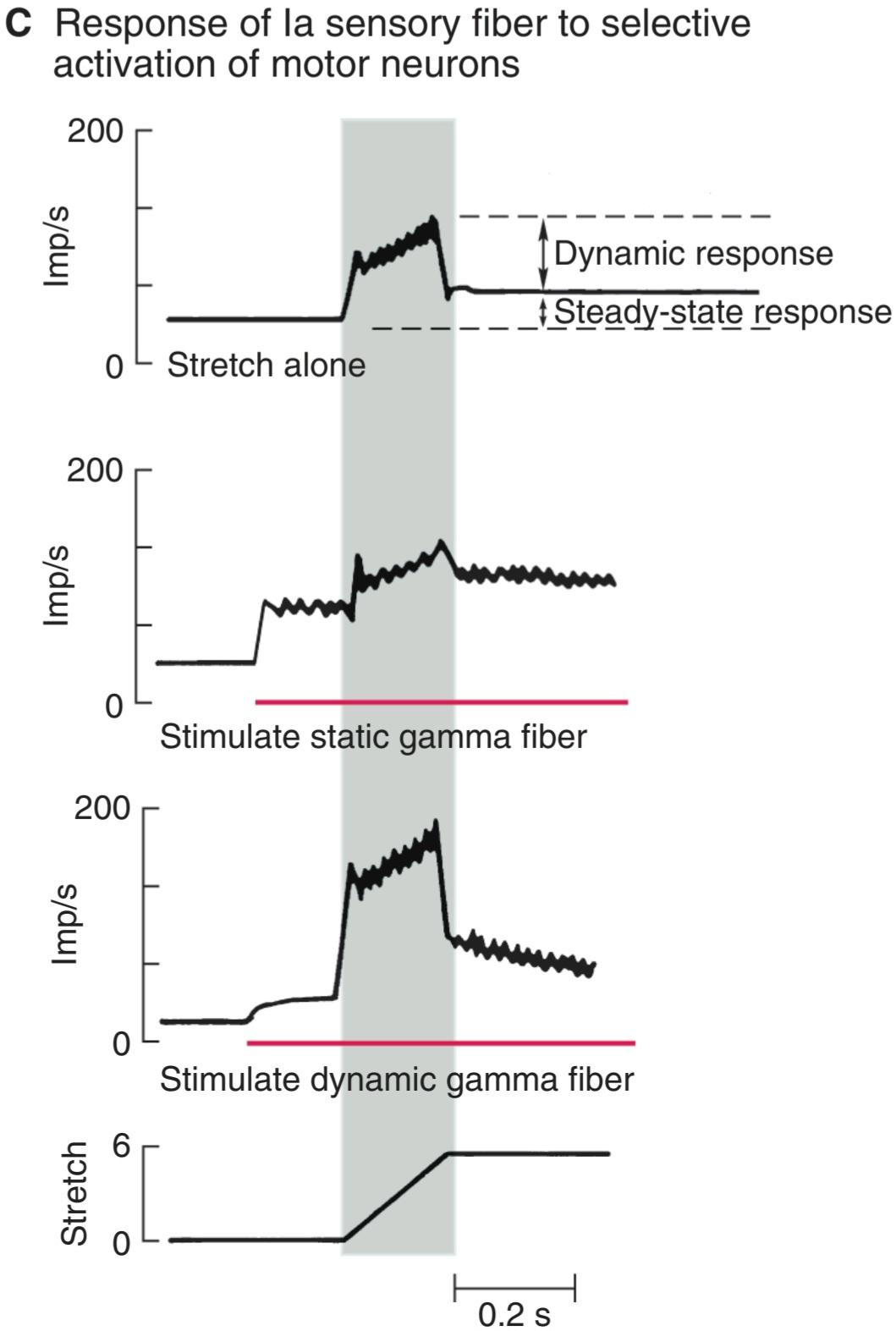
FIGURE 12-2 Mammalian muscle spindle. A) Diagrammatic representation of the main components of mammalian muscle spindle including intrafusal muscle fibers, afferent sensory fiber endings, and efferent motor fibers (y-motor neurons). B) Three types of intrafusal muscle fibers: dynamic nuclear bag, static nuclear bag, and nuclear chain fibers. A single la afferent fiber innervates all three types of fibers to form a primary sensory ending. A group II sensory fiber innervates nuclear chain and static bag fibers to form a secondary sensory ending. Dynamic y-motor neurons innervate dynamic bag fibers; static y-motor neurons innervate combinations of chain and static bag fibers. C) Comparison of discharge pattern of la afferent activity during stretch alone and during stimulation of static or dynamic y-motor neurons. Without y-stimulation, Ia fibers show a small dynamic response to muscle stretch and a modest increase in steady-state firing. When static y-motor neurons are activated, the steady-state response increases and the dynamic response decreases. When dynamic y-motor neurons are activated, the dynamic response is markedly increased but the steady-state response gradually returns to its original level. (Reproduced with permission from Gray H: Gray’s Anatomy: The Anatomical Basis of Clinical Practice, 40th ed. St. Louis, MO: Churchill Livingstone/Elsevier; 2009.)
شکل 12-2 دوک عضلانی پستانداران. الف) نمایش نموداری اجزای اصلی دوک عضلانی پستانداران از جمله فیبرهای عضلانی داخل فیوزال، انتهای فیبر حسی آوران و فیبرهای حرکتی وابران (نرونهای حرکتی y). ب) سه نوع الیاف عضلانی داخل لوله ای: کیسه هسته ای پویا، کیسه هسته ای استاتیک و الیاف زنجیره هسته ای. یک فیبر آوران لا هر سه نوع فیبر را عصب دهی میکند تا یک انتهای حسی اولیه را تشکیل دهد. یک فیبر حسی گروه دوم، زنجیره هسته ای و الیاف کیسه ایستا را عصب میکند تا یک انتهای حسی ثانویه را تشکیل دهد. نورونهای پویا y-motor الیاف کیسه پویا را عصب دهی میکنند. نورونهای استاتیک y-motor ترکیبی از رشتههای زنجیره ای و کیسه ای استاتیک را عصب دهی میکنند. ج) مقایسه الگوی تخلیه فعالیت لا آوران در طول کشش به تنهایی و در حین تحریک نورونهای حرکتی Y استاتیک یا پویا. بدون تحریک y، فیبرهای Ia یک واکنش دینامیکی کوچک به کشش عضلانی و افزایش متوسطی در شلیک حالت پایدار نشان میدهند. هنگامیکه نورونهای استاتیک y-motor فعال میشوند، پاسخ حالت پایدار افزایش مییابد و پاسخ دینامیکی کاهش مییابد. هنگامیکه نورونهای y-motor پویا فعال میشوند، پاسخ دینامیکی به طور قابل توجهی افزایش مییابد، اما پاسخ حالت پایدار به تدریج به سطح اولیه خود باز میگردد. (بازتولید شده با اجازه از Gray H: Gray’s Anatomy: The Anatomical Base of Clinical Practice, 40th ed. St. Louis, MO: Churchill Livingstone/Elsevier; 2009.)
The intrafusal fibers are positioned in parallel to the extrafusal fibers (the regular contractile units of the muscle) with the ends of the spindle capsule attached to the tendons at either end of the muscle. Intrafusal fibers do not contribute to the overall contractile force of the muscle, but rather serve a pure sensory function. There are two types of intrafusal fibers in mammalian muscle spindles. The first type contains many nuclei in a dilated central area and is called a nuclear bag fiber (Figure 12-2B). There are two subtypes of nuclear bag fibers: dynamic and static. The second intrafusal fiber type, the nuclear chain fiber, is thinner and shorter and lacks a definite bag. Typically, each muscle spindle contains two or three nuclear bag fibers and about five nuclear chain fibers.
فیبرهای اینترافوزال به موازات فیبرهای خارج رحمی(واحدهای انقباضی منظم عضله) قرار میگیرند و انتهای کپسول دوک به تاندونها در دو انتهای عضله متصل است. فیبرهای داخل رحمیبه نیروی انقباضی کلی عضله کمک نمیکنند، بلکه عملکرد حسی خالص را ایفا میکنند. دو نوع فیبر داخل رحمیدر دوک عضلانی پستانداران وجود دارد. نوع اول حاوی هستههای زیادی در ناحیه مرکزی متسع است و فیبر کیسه هسته ای نامیده میشود (شکل 12-2B). دو نوع از الیاف کیسه هسته ای وجود دارد: دینامیک و استاتیک. نوع دوم فیبر اینترافیوزال، فیبر زنجیره هسته ای، نازک تر و کوتاه تر است و کیسه مشخصی ندارد. به طور معمول، هر دوک عضلانی حاوی دو یا سه فیبر کیسه هسته ای و حدود پنج رشته زنجیره هسته ای است.
There are two kinds of sensory endings in each spindle, a single primary (group Ia) ending and up to eight secondary (group II) endings (Figure 12– 2B). The Ia afferent fiber wraps around the center of the dynamic and static nuclear bag fibers and nuclear chain fibers. Group II sensory fibers are located adjacent to the centers of the static nuclear bag and nuclear chain fibers; these fibers do not innervate the dynamic nuclear bag fibers. Ia afferents are very sensitive to the velocity of the change in muscle length during a stretch (dynamic response); thus, they provide information about the speed of movements and allow for quick corrective movements. The steady-state (tonic) activity of group Ia and II afferents provide information on steady-state length of the muscle (static response). The top trace in Figure 12-2C shows the dynamic and static components of activity in a la afferent during muscle stretch. Note that they discharge most rapidly while the muscle is being stretched (shaded area of graphs) and less rapidly during sustained stretch.
دو نوع انتهای حسی در هر دوک وجود دارد، یک انتهای اولیه منفرد (گروه Ia) و حداکثر هشت انتهای ثانویه (گروه II) (شکل 12-2B). فیبر آوران Ia دور مرکز الیاف کیسه هسته ای پویا و استاتیک و الیاف زنجیره هسته ای میپیچد. الیاف حسی گروه دوم در مجاورت مراکز کیسه هسته ای ساکن و الیاف زنجیره هسته ای قرار دارند. این الیاف الیاف کیسه هسته ای پویا را عصب دهی نمیکنند. آورانهای Ia به سرعت تغییر طول عضله در طول کشش بسیار حساس هستند (پاسخ پویا). بنابراین، آنها اطلاعاتی در مورد سرعت حرکات ارائه میدهند و امکان حرکات اصلاحی سریع را فراهم میکنند. فعالیت حالت پایدار (تونیک) آورانهای گروه Ia و II اطلاعاتی را در مورد طول حالت پایدار عضله (پاسخ استاتیک) ارائه میدهد. رد بالایی در شکل 12-2C مؤلفههای پویا و ایستا فعالیت در a la afferent را در طول کشش عضلانی نشان میدهد. توجه داشته باشید که زمانی که عضله در حال کشیده شدن است (منطقه سایه دار نمودارها) آنها سریعتر تخلیه میشوند و در طول کشش پایدار با سرعت کمتری تخلیه میشوند.
The spindles have a motor nerve supply of their own called y-motor neurons; they are 3-6 μm in diameter and constitute about 30% of the fibers in the ventral roots. There are two types of y-motor neurons: dynamic that supply dynamic nuclear bag fibers and static that supply static nuclear bag fibers and nuclear chain fibers. Activation of dynamic y-motor neurons increases the dynamic sensitivity of the group la endings. Activation of the static y-motor neurons increases the tonic level of activity in both group Ia and II endings, decreases the dynamic sensitivity of group la afferents, and can prevent silencing of la afferents during muscle stretch (Figure 12-2C).
دوکها منبع عصب حرکتی خود را دارند که نورونهای حرکتی y نامیده میشوند. قطر آنها 3-6 میکرومتر است و حدود 30 درصد از الیاف ریشههای شکمیرا تشکیل میدهند. دو نوع نورون y-motor وجود دارد: دینامیک که فیبرهای کیسه هسته ای پویا را تامین میکند و استاتیک که الیاف کیسه هسته ای ساکن را تامین میکند و الیاف زنجیره هسته ای. فعال شدن نورونهای y-motor پویا حساسیت دینامیکی انتهای گروه la را افزایش میدهد. فعال شدن نورونهای استاتیک y-motor سطح تونیک فعالیت را در انتهای هر دو گروه Ia و II افزایش میدهد، حساسیت دینامیکی آورانهای گروه la را کاهش میدهد و میتواند از خاموش شدن آورانهای la در طول کشش عضلانی جلوگیری کند (شکل 12-2C).
CENTRAL CONNECTIONS OF AFFERENT FIBERS
Ia fibers end directly on motor neurons supplying the extrafusal fibers of the same muscle (Figure 12-3). The reaction time is the interval between the application of a stimulus and the response. In humans, the reaction time for a stretch reflex is 19-24 ms. Weak stimulation of the sensory nerve from the muscle that stimulates only Ia fibers causes a contractile response with a similar latency. Because the conduction velocities of the afferent and efferent fiber types are known and the distance from the muscle to the spinal cord can be measured, it is possible to calculate how much of the reaction time was taken up by conduction to and from the spinal cord. When this value is subtracted from the reaction time, the remainder (central delay) is the time taken for the reflex activity to traverse the spinal cord. The central delay for the knee jerk reflex is 0.6-0.9 ms. Because the minimum synaptic delay is 0.5 ms, only one synapse could have been traversed.
اتصالات مرکزی الیاف آور
فیبرهای Ia مستقیماً به نورونهای حرکتی ختم میشوند که فیبرهای خارج رحمیهمان عضله را تأمین میکنند (شکل 12-3). زمان واکنش فاصله بین اعمال یک محرک و پاسخ است. در انسان، زمان واکنش برای رفلکس کششی 19-24 میلی ثانیه است. تحریک ضعیف عصب حسی از عضله ای که فقط فیبرهای Ia را تحریک میکند باعث پاسخ انقباضی با تاخیر مشابه میشود. از آنجایی که سرعتهای هدایت انواع فیبر آوران و وابران شناخته شده است و فاصله ماهیچه تا نخاع قابل اندازه گیری است، میتوان محاسبه کرد که رسانش به نخاع و از نخاع چه مقدار از زمان واکنش را گرفته است. هنگامیکه این مقدار از زمان واکنش کم میشود، باقیمانده (تاخیر مرکزی) زمان صرف شده برای فعالیت رفلکس برای عبور از طناب نخاعی است. تاخیر مرکزی برای رفلکس حرکتی زانو 0.6-0.9 میلی ثانیه است. از آنجایی که حداقل تأخیر سیناپسی 0.5 میلی ثانیه است، تنها یک سیناپس را میتوان عبور داد.

FIGURE 12-3 Diagram illustrating the pathways responsible for the stretch reflex and the inverse stretch reflex. Stretch stimulates the muscle spindle, which activates la fibers that excite the motor neuron. Stretch also stimulates the Golgi tendon organ, which activates Ib fibers that excite an interneuron that releases the inhibitory mediator glycine. With strong stretch, the resulting hyperpolarization of the motor neuron is so great that it stops discharging.
شکل 12-3 نمودار مسیرهای مسئول رفلکس کشش و رفلکس کشش معکوس را نشان میدهد. کشش دوک عضلانی را تحریک میکند، که فیبرهای لا را فعال میکند که نورون حرکتی را تحریک میکند. کشش همچنین اندام تاندون گلژی را تحریک میکند، که فیبرهای Ib را فعال میکند که نورون داخلی را تحریک میکند که واسطه مهارکننده گلیسین را آزاد میکند. با کشش قوی، هیپرپلاریزه شدن نورون حرکتی آنقدر زیاد است که تخلیه آن متوقف میشود.
FUNCTION OF MUSCLE SPINDLES
When the muscle spindle is stretched, its sensory endings are distorted and receptor potentials are generated. These in turn set up action potentials in the sensory fibers at a frequency proportional to the degree of stretching. Because the spindle is in parallel with the extrafusal fibers, when the muscle is passively stretched, the spindles are also stretched, referred to as “loading the spindle.” This initiates reflex contraction of the extrafusal fibers in the muscle. On the other hand, the spindle afferents characteristically stop firing when the muscle is made to contract by electrical stimulation of the a-motor neurons to the extrafusal fibers because the muscle shortens while the spindle is unloaded(Figure 12-4).
عملکرد دوکهای عضلانی
هنگامیکه دوک عضلانی کشیده میشود، انتهای حسی آن مخدوش میشود و پتانسیلهای گیرنده ایجاد میشود. اینها به نوبه خود پتانسیل عمل را در فیبرهای حسی در فرکانس متناسب با درجه کشش ایجاد میکنند. از آنجایی که دوک به موازات فیبرهای خارج رحمیاست، زمانی که عضله به صورت غیرفعال کشیده میشود، دوکها نیز کشیده میشوند که به آن “بارگذاری دوک” میگویند. این کار باعث انقباض رفلکس فیبرهای خارج رحمیدر عضله میشود. از سوی دیگر، زمانی که عضله با تحریک الکتریکی نورونهای a-motor به فیبرهای خارج رحمیمنقبض میشود، آورانهای دوک مشخصاً شلیک میکنند، زیرا عضله در حالی که دوک تخلیه میشود کوتاه میشود (شکل 12-4).
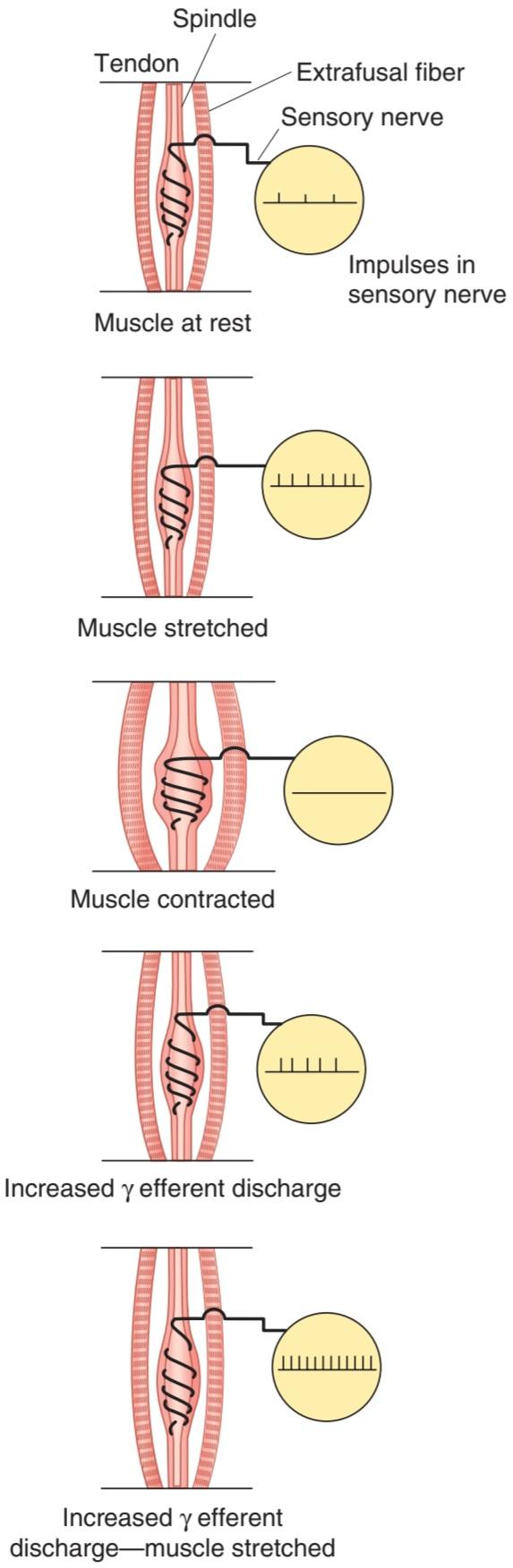
FIGURE 12-4 Effect of various conditions on muscle spindle discharge. When the whole muscle is stretched, the muscle spindle is also stretched and its sensory endings are activated at a frequency proportional to the degree of stretching (“loading the spindle”). Spindle afferents stop firing when the muscle contracts (“unloading the spindle”). Stimulation of y-motor neurons causes the contractile ends of the intrafusal fibers to shorten. This stretches the nuclear bag region, initiating impulses in sensory fibers. If the whole muscle is stretched during stimulation of the y-motor neurons, the rate of discharge in sensory fibers is further increased.
شکل 12-4 اثر شرایط مختلف بر ترشح دوک عضلانی. هنگامیکه کل عضله کشیده میشود، دوک عضلانی نیز کشیده میشود و انتهای حسی آن با فرکانس متناسب با میزان کشش (“بارگذاری دوک”) فعال میشود. آورانهای دوک با انقباض عضله (“تخلیه دوک”) شروع به کار نمیکنند. تحریک نورونهای y-motor باعث کوتاه شدن انتهای انقباضی فیبرهای داخل رحمیمیشود. این امر ناحیه کیسه هسته ای را کشیده و باعث ایجاد تکانه در الیاف حسی میشود. اگر کل عضله در طول تحریک نورونهای حرکتی y کشیده شود، میزان تخلیه در رشتههای حسی بیشتر میشود.
The muscle spindle and its reflex connections constitute a feedback device that operates to maintain muscle length. If the muscle is stretched, spindle discharge increases and reflex shortening is produced. If the muscle is shortened without a change in y-motor neuron discharge, spindle afferent activity decreases and the muscle relaxes.
دوک عضلانی و اتصالات رفلکس آن یک دستگاه بازخوردی را تشکیل میدهند که برای حفظ طول عضله عمل میکند. اگر عضله کشیده شود، ترشح دوک افزایش مییابد و کوتاه شدن رفلکس ایجاد میشود. اگر عضله بدون تغییر در تخلیه نورون y حرکتی کوتاه شود، فعالیت آوران دوک کاهش مییابد و عضله شل میشود.
Dynamic and static responses of muscle spindle afferents influence physiologic tremor. The response of the Ia sensory fiber endings to the dynamic (phasic) as well as the static events in the muscle is important because the prompt, marked phasic response helps dampen oscillations caused by conduction delays in the feedback loop regulating muscle length. Normally a small oscillation occurs in this feedback loop. This physiologic tremor has low amplitude (barely visible to the naked eye) and a frequency of approximately 10 Hz. Physiologic tremor is a normal phenomenon that affects everyone while maintaining posture or during movements. However, the tremor would be more prominent if it were not for the sensitivity of the spindle to velocity of stretch. It can become exaggerated in some situations such as when we are anxious or tired or because of drug toxicity. Numerous factors contribute to the genesis of physiologic tremor. It is dependent on both central (inferior olive) sources and peripheral factors including motor unit firing rates, reflexes, and mechanical
resonance.
پاسخهای پویا و استاتیک آورانهای دوک عضلانی بر لرزش فیزیولوژیک تأثیر میگذارد. پاسخ پایانههای فیبر حسی Ia به پویا (فازی) و همچنین رویدادهای ایستا در عضله مهم است زیرا پاسخ فازی سریع و مشخص به کاهش نوسانات ناشی از تاخیرهای هدایت در حلقه بازخورد تنظیم کننده طول عضله کمک میکند. به طور معمول یک نوسان کوچک در این حلقه بازخورد رخ میدهد. این لرزش فیزیولوژیک دارای دامنه کم (به سختی با چشم غیر مسلح قابل مشاهده) و فرکانس تقریباً 10 هرتز است. لرزش فیزیولوژیک یک پدیده طبیعی است که با حفظ وضعیت بدن یا در حین حرکات، همه افراد را تحت تاثیر قرار میدهد. با این حال، اگر حساسیت دوک به سرعت کشش نبود، لرزش برجستهتر بود. در برخی موقعیتها، مانند مضطرب یا خستگی یا به دلیل سمیت دارو، ممکن است اغراقآمیز شود. عوامل متعددی در پیدایش لرزش فیزیولوژیک نقش دارند. هم به منابع مرکزی (زیتون تحتانی) و هم به عوامل محیطی از جمله سرعت شلیک واحد حرکتی، رفلکسها و مکانیکی وابسته است.
رزونانس
RECIPROCAL INNERVATION
When a stretch reflex occurs, the opposing muscles relax due to reciprocal innervation. Impulses in the Ia fibers from the muscle spindles of the protagonist muscle cause postsynaptic inhibition of the motor neurons to the antagonists. A collateral from each Ia fiber passes in the spinal cord to an inhibitory interneuron that synapses on a motor neuron supplying the antagonist muscles. This example of postsynaptic inhibition is discussed in Chapter 6, and the pathway is illustrated in Figure 6-5.
عصب متقابل
هنگامیکه یک رفلکس کششی رخ میدهد، عضلات مقابل به دلیل عصب دهی متقابل شل میشوند. تکانههای موجود در رشتههای Ia از دوکهای عضلانی عضله پروتاگونیست باعث مهار پس سیناپسی نورونهای حرکتی به آنتاگونیستها میشود. وثیقهای از هر فیبر Ia در نخاع به یک نورون بازدارنده میرود که روی یک نورون حرکتی که عضلات آنتاگونیست را تأمین میکند سیناپس میکند. این مثال از بازداری پس سیناپسی در فصل 6 مورد بحث قرار گرفته است و مسیر در شکل 6-5 نشان داده شده است.
EFFECTS OF Y-MOTOR NEURON DISCHARGE
Activation of y-motor neurons produces a very different picture from that produced by activation of the a-motor neurons. Activation of y-motor neurons does not lead directly to detectable contraction of the muscles because intrafusal fibers are not strong enough or plentiful enough to cause shortening. However, their activation does cause the contractile ends of the intrafusal fibers to shorten and therefore stretches the nuclear bag portion of the spindles, deforming the endings, and initiating impulses in la fibers (Figure 12-4). This in turn can lead to reflex contraction of the muscle. Thus, muscles can be made to contract via activation of the a-motor neurons that innervate the extrafusal fibers or the y- motor neurons that initiate contraction indirectly via the stretch reflex.
اثرات تخلیه نورون Y-موتور
فعالسازی نورونهای حرکتی y تصویری بسیار متفاوت از آنچه در اثر فعالسازی نورونهای a-motor ایجاد میشود، ایجاد میکند. فعال شدن نورونهای حرکتی y مستقیماً منجر به انقباض قابل تشخیص ماهیچهها نمیشود، زیرا فیبرهای داخل رحمیبه اندازه کافی قوی یا فراوان نیستند که باعث کوتاه شدن آنها شوند. با این حال، فعال شدن آنها باعث کوتاه شدن انتهای انقباضی فیبرهای داخل لوله میشود و بنابراین قسمت کیسه هستهای دوکها کشیده میشود، انتهای آنها تغییر شکل میدهد و تکانههایی را در الیاف ایجاد میکند (شکل 12-4). این به نوبه خود میتواند منجر به انقباض رفلکس عضله شود. بنابراین، ماهیچهها را میتوان از طریق فعالسازی نورونهای a-motor که فیبرهای خارج از جریان را عصببندی میکنند یا نورونهای y-موتور که انقباض غیرمستقیم را از طریق رفلکس کشش آغاز میکنند، منقبض کرد.
If the whole muscle is stretched during stimulation of the y-motor neurons, the rate of discharge in the Ia fibers is further increased (Figure 12-4). Increased y-motor neuron activity thus increases spindle sensitivity during stretch.
اگر کل عضله در طول تحریک نورونهای حرکتی y کشیده شود، میزان تخلیه در رشتههای Ia بیشتر افزایش مییابد (شکل 12-4). افزایش فعالیت نورون y-motor در نتیجه حساسیت دوک را در طول کشش افزایش میدهد.
In response to descending excitatory input to spinal motor circuits, both a- and y-motor neurons are activated. Because of this “a-y coactivation,” intrafusal and extrafusal fibers shorten together, and spindle afferent activity can occur throughout the period of muscle contraction. In this way, the spindle remains capable of responding to stretch and reflexively adjusting a-motor neuron discharge.
در پاسخ به ورودی تحریکی نزولی به مدارهای حرکتی ستون فقرات، هر دو نورون a- و y-motor فعال میشوند. به دلیل این “همفعال سازی a-y”، فیبرهای داخل و خارج با هم کوتاه میشوند و فعالیت آوران دوک میتواند در طول دوره انقباض عضلانی رخ دهد. به این ترتیب، دوک میتواند به کشش پاسخ دهد و تخلیه نورون یک حرکتی را به صورت انعکاسی تنظیم کند.
CONTROL OF y-MOTOR NEURON DISCHARGE
The y-motor neurons are regulated by descending tracts originating in areas of the brain that also control a-motor neurons (described below). Via these pathways, the sensitivity of the muscle spindles and hence the threshold of the stretch reflexes in various parts of the body can be adjusted and shifted to meet the needs of postural control.
کنترل تخلیه نورون y-موتور
نورونهای y-motor توسط دستگاههای نزولی که در مناطقی از مغز منشا میگیرند تنظیم میشوند که نورونهای a-motor را نیز کنترل میکنند (در زیر توضیح داده شده است). از طریق این مسیرها، حساسیت دوکهای عضلانی و در نتیجه آستانه رفلکسهای کششی در قسمتهای مختلف بدن را میتوان تنظیم و تغییر داد تا نیازهای کنترل وضعیتی را برآورده کند.
Other factors also influence y-motor neuron activity. Anxiety causes an increased discharge, a fact that probably explains the hyperactive tendon reflexes sometimes seen in anxious patients. In addition, unexpected movement is associated with a greater efferent discharge. Stimulation of the skin, especially by noxious agents, increases y-motor neuron activity to ipsilateral flexor muscle spindles while decreasing that to extensors and produces the opposite pattern in the opposite limb. Trying to pull the hands apart when the flexed fingers are hooked together facilitates the knee jerk reflex (Jendrassik maneuver), and this may also be due to increase y-motor neuron discharge initiated by afferent impulses from the hands.
عوامل دیگر نیز بر فعالیت نورون y-motor تاثیر میگذارد. اضطراب باعث افزایش ترشح میشود، واقعیتی که احتمالاً رفلکسهای تاندون بیش فعال که گاهی در بیماران مضطرب دیده میشود را توضیح میدهد. علاوه بر این، حرکت غیرمنتظره با ترشحات وابران بیشتر همراه است. تحریک پوست، به ویژه توسط عوامل مضر، فعالیت نورونهای حرکتی y را به دوکهای عضله فلکسور همان طرف افزایش میدهد در حالی که آن را به اکستانسورها کاهش میدهد و الگوی مخالف را در اندام مقابل ایجاد میکند. تلاش برای باز کردن دستها زمانی که انگشتان خم شده به هم قلاب میشوند، رفلکس حرکتی زانو (مانور جندراسیک) را تسهیل میکند، و این ممکن است به دلیل افزایش تخلیه نورون y حرکتی باشد که توسط تکانههای آوران از دستها آغاز میشود.
INVERSE STRETCH REFLEX
Up to a point, the harder a muscle is stretched, the stronger is the reflex contraction. However, when the tension becomes great enough, contraction suddenly ceases and the muscle relaxes. This relaxation in response to strong stretch is called the inverse stretch reflex. The receptor for the inverse stretch reflex is in the Golgi tendon organ (Figure 12-5) that consists of a netlike collection of knobby nerve endings among the fascicles of a tendon. There are 3-25 muscle fibers per tendon organ. The fibers from the Golgi tendon organs are the Ib group of myelinated, rapidly conducting sensory nerve fibers. Activation of these Ib fibers leads to IPSPs in the motor neurons that supply the muscle from which the fibers arise. The Ib fibers end in the spinal cord on inhibitory interneurons that terminate directly on the motor neurons (Figure 12- 3). They also make excitatory connections with motor neurons supplying antagonists to the muscle.
رفلکس کششی معکوس
تا یک نقطه، هرچه عضله سخت تر کشیده شود، انقباض رفلکس قوی تر است. با این حال، زمانی که تنش به اندازه کافی زیاد شود، انقباض ناگهانی متوقف میشود و عضله شل میشود. این آرامش در پاسخ به کشش شدید، رفلکس کششی معکوس نامیده میشود. گیرنده رفلکس کشش معکوس در اندام تاندون گلژی است (شکل 12-5) که از مجموعه شبکه مانندی از انتهای عصب دستگیره ای در میان فاسیکلهای تاندون تشکیل شده است. 3-25 فیبر عضلانی در هر اندام تاندون وجود دارد. الیاف ارگانهای تاندون گلژی گروه Ib از رشتههای عصبی حسی میلین دار هستند که به سرعت رسانا میشوند. فعال شدن این فیبرهای Ib منجر به ایجاد IPSP در نورونهای حرکتی میشود که عضلهای را که فیبرها از آن بیرون میآیند، تامین میکنند. الیاف Ib به نخاع روی نورونهای بازدارنده ای ختم میشوند که مستقیماً به نورونهای حرکتی ختم میشوند (شکل 12-3). آنها همچنین با نورونهای حرکتی که آنتاگونیستها را به عضله میرسانند، ارتباط تحریکی برقرار میکنند.

FIGURE 12-5 Golgi tendon organ. This organ is the receptor for the inverse stretch reflex and consists of a netlike collection of knobby nerve endings among the fascicles of a tendon. The innervation is the Ib group of myelinated, rapidly conducting sensory nerve fibers. (Reproduced with permission from Gray H: Gray’s Anatomy: The Anatomical Basis of Clinical Practice, 40th ed. St. Louis,MO: Churchill Livingstone/Elsevier; 2009.)
شکل 12-5 اندام تاندون گلژی. این اندام گیرنده رفلکس کششی معکوس است و از مجموعه شبکهای از انتهای عصب دستگیرهای در میان فاسیکلهای تاندون تشکیل شده است. عصب دهی گروه Ib از رشتههای عصبی حسی میلین دار است که به سرعت هدایت میشوند. (تکثیر شده با اجازه از Gray H: Gray’s Anatomy: The Anatomical Base of Clinical Practice، نسخه 40 سنت لوئیس، MO: چرچیل لیوینگستون/الزویر؛ 2009.)
Because the Golgi tendon organs, unlike the spindles, are in series with the muscle fibers, they are stimulated by both passive stretch and active contraction of the muscle. The threshold of the Golgi tendon organs is low. The degree of stimulation by passive stretch is not great because the more elastic muscle fibers take up much of the stretch; therefore, it takes a strong stretch to produce relaxation. However, discharge is regularly produced by contraction of the muscle, and the Golgi tendon organ thus functions as a transducer in a feedback circuit that regulates muscle force in a manner analogous to the spindle feedback circuit that regulates muscle length.
از آنجایی که اندامهای تاندون گلژی، بر خلاف دوکها، با رشتههای عضلانی در ردیف قرار دارند، هم با کشش غیرفعال و هم با انقباض فعال عضله تحریک میشوند. آستانه اندامهای تاندون گلژی کم است. درجه تحریک توسط کشش غیرفعال زیاد نیست زیرا فیبرهای عضلانی الاستیک بیشتر بخش زیادی از کشش را به خود اختصاص میدهند. بنابراین، برای ایجاد آرامش، کشش قوی لازم است. با این حال، تخلیه به طور منظم با انقباض عضله تولید میشود، و بنابراین اندام تاندون گلژی به عنوان یک مبدل در یک مدار بازخوردی عمل میکند که نیروی عضلانی را به شیوه ای مشابه با مدار بازخورد دوکی که طول عضله را تنظیم میکند، تنظیم میکند.
The primary endings in the spindles and the Golgi tendon organs together regulate the velocity of the muscle contraction, muscle length, and muscle force. The interaction of spindle discharge, tendon organ discharge, and reciprocal innervation determines the firing rate of a-motor neurons (Clinical Box 12-2).
انتهای اولیه در دوکها و اندامهای تاندون گلژی با هم سرعت انقباض عضلانی، طول عضله و نیروی عضلانی را تنظیم میکنند. برهمکنش تخلیه دوک، تخلیه اندام تاندون و عصب دهی متقابل نرخ شلیک نورونهای a-motor را تعیین میکند (جعبه بالینی 12-2).
CLINICAL BOX 12.2
Clonus
Clonus is characteristic of states in which there is increased y-motor neuron activity. This neurologic sign is the occurrence of regular, repetitive, rhythmic contractions of a muscle subjected to sudden, maintained stretch. Only sustained clonus with five or more beats is considered abnormal. Ankle clonus is initiated by brisk, maintained dorsiflexion of the foot; the response is rhythmic plantar flexion at the ankle. The stretch reflex-inverse stretch reflex sequence may contribute to this response. However, it can occur on the basis of synchronized motor neuron discharge without Golgi tendon organ activation. The spindles of the tested muscle are hyperactive, and the burst of impulses from them activates all the motor neurons supplying the muscle at once. The consequent muscle contraction stops spindle discharge. However, the stretch has been maintained, and as soon as the muscle relaxes it is again stretched and the spindles stimulated. There are numerous causes of abnormal clonus including traumatic brain injury, brain tumors, strokes, and multiple sclerosis. Clonus may also occur in spinal cord injury that disrupts the descending cortical input to a spinal glycinergic inhibitory interneuron called the Renshaw cell. This cell receives excitatory input from a-motor neurons via axon collaterals (and in turn it inhibits the same a-motor neuron). In addition, cortical fibers activating ankle flexors synapse on Renshaw cells (as well as type la inhibitory interneurons) that inhibit the antagonistic ankle extensors. This circuitry prevents reflex stimulation of the extensors when flexors are active. Therefore, when the descending cortical fibers are damaged (upper motor neuron lesion), the inhibition of antagonists is absent. The result is repetitive, sequential contraction of ankle flexors and extensors (clonus). Clonus may be seen in patients with amyotrophic lateral sclerosis (ALS), stroke, multiple sclerosis, spinal cord damage, epilepsy, liver or kidney failure, and hepatic encephalopathy.
جعبه بالینی 12.2
کلونوس
کلونوس مشخصه حالتهایی است که در آن فعالیت نورون y-motor افزایش یافته است. این علامت عصبی بروز انقباضات منظم، مکرر و ریتمیک عضله ای است که تحت کشش ناگهانی و مداوم قرار میگیرد. فقط کلونوس پایدار با پنج ضربه یا بیشتر غیر طبیعی در نظر گرفته میشود. کلونوس مچ پا با دورسی فلکشن سریع پا شروع میشود. پاسخ، فلکشن ریتمیک کف پا در مچ پا است. توالی رفلکس کششی معکوس رفلکس کششی ممکن است به این پاسخ کمک کند. با این حال، میتواند بر اساس تخلیه همزمان نورون حرکتی بدون فعال شدن اندام تاندون گلژی رخ دهد. دوکهای عضله آزمایش شده بیش فعال هستند و انفجار تکانههای آنها همه نورونهای حرکتی تامین کننده عضله را به یکباره فعال میکند. انقباض عضلانی در نتیجه ترشح دوک را متوقف میکند. با این حال، کشش حفظ شده است و به محض شل شدن عضله دوباره کشیده میشود و دوکها تحریک میشوند. دلایل متعددی برای کلونوس غیرطبیعی وجود دارد که از جمله آنها میتوان به آسیب مغزی تروماتیک، تومورهای مغزی، سکته مغزی و ام اس اشاره کرد. کلونوس همچنین ممکن است در آسیب طناب نخاعی رخ دهد که ورودی قشر نزولی به یک نورون بازدارنده گلیسینرژیک ستون فقرات به نام سلول رنشا را مختل میکند. این سلول ورودی تحریکی را از نورونهای a-motor از طریق وثیقههای آکسون دریافت میکند (و به نوبه خود همان نورون a-motor را مهار میکند). علاوه بر این، فیبرهای قشری فعال کننده فلکسورهای مچ پا روی سلولهای رنشا (و همچنین نورونهای بازدارنده نوع la) سیناپس میشوند که اکستانسورهای آنتاگونیست مچ پا را مهار میکنند. این مدار از تحریک رفلکس اکستانسورها در هنگام فعال بودن فلکسورها جلوگیری میکند. بنابراین، هنگامیکه فیبرهای قشر نزولی آسیب میبینند (ضایعه نورون حرکتی فوقانی)، مهار آنتاگونیستها وجود ندارد. نتیجه انقباض مکرر و متوالی فلکسورها و اکستانسورهای مچ پا (کلونوس) است. کلونوس ممکن است در بیماران مبتلا به اسکلروز جانبی آمیوتروفیک (ALS)، سکته مغزی، مولتیپل اسکلروزیس، آسیب نخاعی، صرع، نارسایی کبد یا کلیه و انسفالوپاتی کبدی دیده شود.
THERAPEUTIQ HIGHLIGHTS
Treatment of clonus often centers on its underlying cause. For some individuals, physical therapy that includes stretching exercises can reduce episodes of clonus. Immunosuppressants (eg, azathioprine and corticosteroids), anticonvulsants (eg, primidone and levetiracetam), and tranquilizers (eg, clonazepam) are beneficial in the treatment of clonus. Botulinum toxin has also been used to block the release of acetylcholine in the muscle, which triggers the rhythmic muscle contractions that are characteristic of clonus.
نکات برجسته درمانی
درمان کلونوس اغلب بر علت اصلی آن متمرکز است. برای برخی افراد، فیزیوتراپی که شامل تمرینات کششی است میتواند دورههای کلونوس را کاهش دهد. داروهای سرکوب کننده ایمنی (مانند آزاتیوپرین و کورتیکواستروئیدها)، داروهای ضد تشنج (مانند پریمیدون و لوتیراستام) و آرام بخشها (مثلاً کلونازپام) در درمان کلونوس مفید هستند. سم بوتولینوم همچنین برای جلوگیری از آزاد شدن استیل کولین در عضله استفاده شده است که باعث انقباضات ریتمیک ماهیچه ای میشود که مشخصه کلونوس است.
MUSCLE TONE
The resistance of a muscle to stretch is often referred to as its tone or tonus. If the motor nerve to a muscle is severed, the muscle offers very little resistance and is said to be flaccid. A hypertonic (spastic) muscle is one in which the resistance to stretch is high because of hyperactive stretch reflexes. Somewhere between the states of flaccidity and spasticity is the ill-defined area of normal tone. The muscles are generally hypotonic when the rate of y-motor neuron discharge is low and hypertonic when it is high.
تون عضلانی
مقاومت یک عضله در برابر کشش اغلب به عنوان تن یا تن آن نامیده میشود. اگر عصب حرکتی عضله قطع شود، عضله مقاومت بسیار کمیاز خود نشان میدهد و گفته میشود که شل است. عضله هیپرتونیک (اسپاستیک) عضله ای است که در آن مقاومت در برابر کشش به دلیل رفلکسهای کششی بیش فعال است. جایی بین حالتهای شلی و اسپاستیسیته، ناحیه تون طبیعی نامشخص است. زمانی که میزان تخلیه نورون y حرکتی کم باشد، عضلات به طور کلی هیپوتونیک و زمانی که میزان آن زیاد است، هیپرتونیک هستند.
When the muscles are hypertonic, the sequence of moderate stretch → muscle contraction, strong stretch → muscle relaxation is clearly seen. Passive flexion of the elbow, for example, meets immediate resistance as a result of the stretch reflex in the triceps muscle. Further stretch activates the inverse stretch reflex. The resistance to flexion suddenly collapses, and the arm flexes. Continued passive flexion stretches the muscle again, and the sequence may be repeated. This sequence of resistance followed by a sudden decrease in resistance when a limb is moved passively is known as the clasp-knife effect because of its resemblance to the closing of a pocket knife.
هنگامیکه عضلات هیپرتونیک هستند، دنباله کشش متوسط ← انقباض عضلانی، کشش قوی ← شل شدن عضلات به وضوح دیده میشود. به عنوان مثال، خم شدن غیرفعال آرنج، در نتیجه رفلکس کشش در عضله سه سر، با مقاومت فوری روبرو میشود. کشش بیشتر رفلکس کشش معکوس را فعال میکند. مقاومت در برابر خم شدن ناگهانی فرو میریزد و بازو خم میشود. ادامه خم شدن غیرفعال دوباره عضله را کش میدهد و این توالی ممکن است تکرار شود. این توالی مقاومت و به دنبال آن کاهش ناگهانی مقاومت در هنگام حرکت غیرفعال یک اندام به دلیل شباهت آن به بسته شدن چاقوی جیبی به اثر چاقوی گیره معروف است.
WITHDRAWAL REFLEX
The withdrawal reflex is a typical polysynaptic reflex that occurs in response to a noxious stimulus to the skin or subcutaneous tissues and muscle. The response is flexor muscle contraction and inhibition of extensor muscles, so that the body part stimulated is flexed and withdrawn from the stimulus. When a strong stimulus is applied to a limb, the response includes not only flexion and withdrawal of that limb but also extension of the opposite limb. This crossed extensor response is properly part of the withdrawal reflex. Strong stimuli can generate activity in the interneuron pool that spreads to all four extremities.
رفلکس عقب نشینی
رفلکس خروج یک رفلکس پلی سیناپسی معمولی است که در پاسخ به یک محرک مضر به پوست یا بافتهای زیر جلدی و ماهیچهها رخ میدهد. پاسخ انقباض عضله فلکسور و مهار عضلات بازکننده است، به طوری که قسمت تحریک شده بدن خم شده و از محرک خارج میشود. هنگامیکه یک محرک قوی بر روی یک اندام اعمال میشود، پاسخ نه تنها شامل خم شدن و عقب نشینی آن اندام بلکه شامل امتداد اندام مقابل نیز میشود. این پاسخ اکستانسور متقاطع به درستی بخشی از رفلکس خروج است. محرکهای قوی میتوانند فعالیتی را در حوضچه بین نورونی ایجاد کنند که به هر چهار اندام گسترش مییابد.
IMPORTANCE OF THE WITHDRAWAL REFLEX
Flexor responses can be produced by innocuous stimulation of the skin or by stretch of the muscle, but strong flexor responses with withdrawal are initiated only by stimuli that are noxious or at least potentially harmful (ie, nociceptive stimuli). The withdrawal reflex serves a protective function as flexion of the stimulated limb gets it away from the source of irritation, and extension of the other limb supports the body.
اهمیت رفلکس عقب نشینی
پاسخهای فلکسور را میتوان با تحریک بی ضرر پوست یا با کشش عضله ایجاد کرد، اما پاسخهای خم کننده قوی با عقب نشینی تنها با محرکهایی شروع میشود که مضر یا حداقل بالقوه مضر هستند (یعنی محرکهای درد). رفلکس خروج یک عملکرد محافظتی دارد زیرا خم شدن اندام تحریک شده آن را از منبع تحریک دور میکند و گسترش اندام دیگر از بدن حمایت میکند.
A weak noxious stimulus to one foot evokes a minimal flexion response; stronger stimuli produce greater and greater flexion as the stimulus irradiates to more and more of the motor neuron pool supplying the muscles of the limb. Stronger stimuli also cause a more prolonged response. A weak stimulus causes one quick flexion movement; a strong stimulus causes prolonged flexion and sometimes a series of flexion movements. This prolonged response is due to prolonged, repeated firing of the motor neurons (after-discharge) that is due to continued bombardment of motor neurons by impulses arriving by complicated and circuitous polysynaptic paths.
یک محرک مضر ضعیف به یک پا، پاسخ خمشی حداقلی را برمیانگیزد. محرکهای قویتر، خمش بیشتر و بیشتر را ایجاد میکنند، زیرا محرک به حوضچه نورون حرکتی بیشتری که ماهیچههای اندام را تامین میکند تابش میکند. محرکهای قویتر نیز باعث پاسخ طولانیتر میشوند. یک محرک ضعیف باعث یک حرکت خم شدن سریع میشود. یک محرک قوی باعث خم شدن طولانی مدت و گاهی یک سری حرکات خمشی میشود. این پاسخ طولانی به دلیل شلیک طولانی مدت و مکرر نورونهای حرکتی (پس از تخلیه) است که به دلیل بمباران مداوم نورونهای حرکتی توسط تکانههایی است که از مسیرهای پیچیده و مداری پلی سیناپسی وارد میشوند.
As the strength of a noxious stimulus is increased, the reaction time is shortened. Spatial and temporal facilitation occur at synapses in the polysynaptic pathway. Stronger stimuli produce more action potentials per second in the active branches and cause more branches to become active; summation of the EPSPs to the threshold level for action potential generation occurs more rapidly.
با افزایش قدرت یک محرک مضر، زمان واکنش کوتاه میشود. تسهیل مکانی و زمانی در سیناپسها در مسیر پلی سیناپسی رخ میدهد. محرکهای قویتر پتانسیل عمل بیشتری در هر ثانیه در شاخههای فعال تولید میکنند و باعث میشوند شاخههای بیشتری فعال شوند. جمع EPSPها به سطح آستانه برای تولید پتانسیل عمل سریعتر رخ میدهد.
SPINAL INTEGRATION OF REFLEXES
The responses to spinal cord injury (SCI) illustrate the integration of reflexes at the spinal level. The deficits seen after SCI vary, of course, depending on the level of the injury. Clinical Box 12-3 provides information on long-term problems related to SCI and recent advancements in treatment options.
ادغام نخاعی رفلکسها
پاسخ به آسیب نخاعی (SCI) ادغام رفلکسها در سطح نخاع را نشان میدهد. کسریهایی که بعد از SCI مشاهده میشود، البته بسته به سطح آسیب متفاوت است. باکس بالینی 12-3 اطلاعاتی در مورد مشکلات طولانی مدت مربوط به SCI و پیشرفتهای اخیر در گزینههای درمانی ارائه میدهد.
CLINICAL BOX 12.3
Spinal Cord Injury
It has been estimated that the worldwide annual incidence of sustaining SCI is between 10 and 83 per million of the population. Leading causes are vehicular accidents, violence, and sports injuries. The mean age of patients who sustain SCI is 33 years old, and men outnumber women with a nearly 4:1 ratio. Approximately 52% of SCI cases result in quadriplegia and about 42% lead to paraplegia. With quadriplegia, the threshold for the withdrawal reflex is very low; even minor noxious stimuli may cause not only prolonged withdrawal of one extremity but marked flexion-extension patterns in the other three limbs. Stretch reflexes are also hyperactive. Afferent stimuli irradiate from one reflex center to another after SCI. When even a relatively minor noxious stimulus is applied to the skin, it may activate autonomic neurons and produce evacuation of the bladder and rectum, sweating, pallor, and blood pressure swings in addition to the withdrawal response. This distressing mass reflex can sometimes be used to give patients with paraplegia a degree of bladder and bowel control. They can be trained to initiate urination and defecation by stroking or pinching their thighs, thus producing an intentional mass reflex. If the cord section is incomplete, the flexor spasms initiated by noxious stimuli can be associated with bursts of pain that are particularly bothersome. They can be treated with considerable success with baclofen, a GABA receptor agonist that crosses the blood-brain barrier and facilitates inhibition.
جعبه بالینی 12.3
آسیب نخاعی
تخمین زده شده است که بروز سالانه پایداری SCI در سراسر جهان بین 10 تا 83 در هر میلیون نفر از جمعیت است. علل اصلی تصادفات خودرو، خشونت و آسیبهای ورزشی است. میانگین سنی بیماران مبتلا به SCI 33 سال است و تعداد مردان با نسبت تقریباً 4:1 از زنان بیشتر است. تقریباً 52 درصد از موارد SCI منجر به کوادری پلژی و حدود 42 درصد منجر به پاراپلژی میشود. با کوادری پلژی، آستانه رفلکس خروج بسیار پایین است. حتی محرکهای مضر جزئی ممکن است نه تنها باعث عقب نشینی طولانی مدت یک اندام شود، بلکه باعث ایجاد الگوهای خمش-کشش مشخص در سه اندام دیگر شود. رفلکسهای کششی نیز بیش فعال هستند. محرکهای آوران از یک مرکز رفلکس به مرکز دیگر پس از SCI تابش میکنند. هنگامیکه حتی یک محرک مضر نسبتاً جزئی روی پوست اعمال میشود، ممکن است نورونهای خودمختار را فعال کند و علاوه بر واکنش خروج، باعث تخلیه مثانه و رکتوم، تعریق، رنگ پریدگی و نوسانات فشار خون شود. گاهی اوقات میتوان از این رفلکس توده ناراحت کننده برای کنترل مثانه و روده به بیماران مبتلا به پاراپلژی استفاده کرد. آنها را میتوان آموزش داد تا با نوازش یا نیشگون گرفتن رانهای خود شروع به دفع ادرار و مدفوع کنند و در نتیجه یک رفلکس توده عمدی ایجاد کنند. اگر بخش طناب ناقص باشد، اسپاسم فلکسور که توسط محرکهای مضر شروع میشود، میتواند با انفجارهای درد همراه باشد که بهخصوص آزاردهنده است. آنها را میتوان با موفقیت قابل توجهی با باکلوفن، یک آگونیست گیرنده GABA که از سد خونی مغزی عبور میکند و مهار را تسهیل میکند، درمان کرد.
THERAPEUTIC HIGHLIGHTS
Treatment of SCI patients presents complex problems. Administration of corticosteroids such as methylprednisolone may have beneficial effects by fostering recovery and minimizing loss of function after SCI. They need to be given soon after the injury and then discontinued because of the well- established deleterious effects of long-term corticosteroid treatment. Their immediate value is likely due to reduction of the inflammatory response in the damaged tissue. Because SCI patients are immobile, a negative nitrogen balance develops and large amounts of body protein are catabolized. Their body weight compresses the circulation to the skin over bony prominences, causing formation of pressure ulcers. The ulcers heal poorly and are prone to infection because of body protein depletion. The tissues that are broken down include the protein matrix of bone and this, plus the immobilization, cause Ca2+ to be released in large amounts, leading to hypercalcemia, hypercalciuria, and formation of calcium stones in the urinary tract. The combination of stones and bladder paralysis cause urinary stasis, which predisposes to urinary tract infection, the most common complication of SCI. The search continues for ways to get axons of neurons in the spinal cord to regenerate. Administration of neurotrophins shows some promise in experimental animals, and so does implantation of embryonic stem cells at the site of injury. Another possibility being explored is bypassing the site of SCI with brain-computer interface devices. However, these novel approaches are a long way from routine clinical use.
نکات برجسته درمانی
درمان بیماران SCI مشکلات پیچیده ای را ارائه میدهد. تجویز کورتیکواستروئیدها مانند متیل پردنیزولون ممکن است اثرات مفیدی با تقویت بهبودی و به حداقل رساندن از دست دادن عملکرد پس از SCI داشته باشد. آنها باید بلافاصله پس از آسیب داده شوند و سپس به دلیل اثرات مضر ثابت شده درمان طولانی مدت کورتیکواستروئید، قطع شوند. ارزش فوری آنها احتمالاً به دلیل کاهش پاسخ التهابی در بافت آسیب دیده است. از آنجایی که بیماران SCI بی حرکت هستند، تعادل نیتروژن منفی ایجاد میشود و مقادیر زیادی پروتئین بدن کاتابولیز میشود. وزن بدن آنها گردش خون را به پوست روی برجستگیهای استخوانی فشرده میکند و باعث ایجاد زخمهای فشاری میشود. زخمها به خوبی بهبود نمییابند و به دلیل کاهش پروتئین بدن مستعد عفونت هستند. بافتهایی که تجزیه میشوند شامل ماتریکس پروتئین استخوان هستند و این به علاوه بیحرکتی باعث میشود که Ca2+ به مقدار زیاد آزاد شود که منجر به هیپرکلسمی، هیپرکلسیوری و تشکیل سنگهای کلسیمیدر دستگاه ادراری میشود. ترکیب سنگ و فلج مثانه باعث استاز ادراری میشود که مستعد عفونت دستگاه ادراری است که شایع ترین عارضه SCI است. جستجو برای یافتن راههایی برای بازیابی آکسونهای نورونهای نخاع ادامه دارد. تجویز نوروتروفینها در حیوانات آزمایشگاهی و همچنین کاشت سلولهای بنیادی جنینی در محل آسیب، امیدبخش است. احتمال دیگری که در حال بررسی است دور زدن محل SCI با دستگاههای رابط مغز و کامپیوتر است. با این حال، این رویکردهای جدید فاصله زیادی با بالینی معمول دارند استفاده کنید.
In all vertebrates, transection of the spinal cord is followed by a period of spinal shock during which all spinal reflex responses are profoundly depressed. Subsequently, reflex responses return and become hyperactive. The duration of spinal shock is proportional to the degree of encephalization of motor function in the various species. In frogs and rats it lasts for minutes; in dogs and cats it lasts for 1-2 h; in monkeys it lasts for days; and in humans it usually lasts for a minimum of 2 weeks.
در تمام مهره داران، قطع نخاع با یک دوره شوک نخاعی همراه است که در طی آن تمام واکنشهای رفلکس نخاعی به شدت تحت فشار قرار میگیرند. متعاقبا، پاسخهای رفلکس برمیگردند و بیش فعال میشوند. طول مدت شوک نخاعی متناسب با درجه انسفالیزاسیون عملکرد حرکتی در گونههای مختلف است. در قورباغهها و موشها چند دقیقه طول میکشد. در سگها و گربهها 1-2 ساعت طول میکشد. در میمونها روزها طول میکشد. و در انسان معمولاً حداقل 2 هفته طول میکشد.
Cessation of tonic bombardment of spinal neurons by excitatory impulses in descending pathways (see below) plays a role in development of spinal shock. In addition, spinal inhibitory interneurons that normally are themselves inhibited may be released from this descending inhibition to become disinhibited. This, in turn, would inhibit motor neurons. The recovery of reflex excitability may be due to the development of denervation hypersensitivity to the mediators released by the remaining spinal excitatory endings. Another contributing factor is sprouting of collaterals from existing neurons, with the formation of additional excitatory endings on interneurons and motor neurons.
توقف بمباران تونیک نورونهای نخاعی توسط تکانههای تحریکی در مسیرهای نزولی (نگاه کنید به زیر) در ایجاد شوک نخاعی نقش دارد. علاوه بر این، نورونهای بازدارنده نخاعی که معمولاً خودشان مهار میشوند، ممکن است از این مهار نزولی رها شوند تا مهار شوند. این به نوبه خود نورونهای حرکتی را مهار میکند. بازیابی تحریک پذیری رفلکس ممکن است به دلیل ایجاد حساسیت بیش از حد عصب کشی به واسطههای آزاد شده توسط پایانههای تحریک کننده ستون فقرات باقی مانده باشد. یکی دیگر از عوامل کمک کننده، جوانه زدن وثیقهها از نورونهای موجود، با تشکیل پایانههای تحریکی اضافی روی نورونهای بین عصبی و نورونهای حرکتی است.
The first reflex response to appear as spinal shock wears off in humans is often a slight contraction of the leg flexors and adductors in response to a noxious stimulus (ie, the withdrawal reflex). In some patients, the knee jerk reflex recovers first. The interval between cord transection and the return of reflex activity is about 2 weeks in the absence of any complications, but if complications are present it is much longer. Once the spinal reflexes begin to reappear after spinal shock, their threshold steadily drops.
اولین واکنش رفلکسی که با از بین رفتن شوک نخاعی در انسان ظاهر میشود، اغلب انقباض خفیف خمکنندهها و کشندههای پا در پاسخ به یک محرک مضر (یعنی رفلکس ترک) است. در برخی از بیماران، رفلکس تکان دادن زانو ابتدا بهبود مییابد. فاصله بین قطع بند ناف و بازگشت فعالیت رفلکس در صورت عدم وجود عارضه حدود 2 هفته است اما در صورت وجود عوارض بسیار بیشتر است. هنگامیکه رفلکسهای نخاعی پس از شوک نخاعی شروع به ظاهر شدن میکنند، آستانه آنها به طور پیوسته کاهش مییابد.
Circuits intrinsic to the spinal cord can produce walking movements when stimulated in a suitable manner even after spinal cord transection in cats and dogs. There are two locomotor pattern generators in the spinal cord: one in the cervical region and one in the lumbar region. However, this does not mean that spinal animals or humans can walk without stimulation; the pattern generator has to be turned on by tonic discharge of a discrete area in the midbrain, the mesencephalic locomotor region, and, of course, this is only possible in patients with incomplete spinal cord transection. Progress is being made in teaching humans with SCI to take a few steps by placing them, with support, on a treadmill.
مدارهای ذاتی نخاع میتوانند حرکات راه رفتن را در صورت تحریک به شیوه ای مناسب حتی پس از قطع نخاع در گربهها و سگها ایجاد کنند. دو ژنراتور الگوی حرکتی در نخاع وجود دارد: یکی در ناحیه گردن و دیگری در ناحیه کمر. با این حال، این بدان معنا نیست که حیوانات نخاعی یا انسان میتوانند بدون تحریک راه بروند. ژنراتور الگو باید با تخلیه تونیک یک ناحیه مجزا در مغز میانی، ناحیه حرکتی مزانسفالیک روشن شود و البته این فقط در بیمارانی که قطع نخاع ناقص دارند امکان پذیر است. پیشرفتهایی در آموزش به انسانهای مبتلا به SCI برای برداشتن چند گام با قرار دادن آنها روی تردمیل با پشتیبانی صورت گرفته است.
GENERAL PRINCIPLES OF CENTRAL ORGANIZATION OF MOTOR PATHWAYS
To voluntarily move a limb, the brain must plan a movement, arrange appropriate motion at many different joints at the same time, and adjust the motion by comparing plan with performance. The motor system “learns by doing” and performance improves with repetition. This involves synaptic plasticity. Damage to the cerebral cortex before or during childbirth or during the first 2-3 years of development can lead to cerebral palsy, a disorder that affects muscle tone, movement, and coordination (Clinical Box 12-4).
اصول کلی سازمان مرکزی راههای موتوری
برای حرکت ارادی یک اندام، مغز باید یک حرکت را برنامه ریزی کند، حرکت مناسب را در مفاصل مختلف به طور همزمان ترتیب دهد و با مقایسه برنامه با عملکرد، حرکت را تنظیم کند. سیستم حرکتی “با انجام دادن یاد میگیرد” و عملکرد با تکرار بهبود مییابد. این شامل پلاستیسیته سیناپسی است. آسیب به قشر مغز قبل یا در حین زایمان یا در طی 2-3 سال اول رشد میتواند منجر به فلج مغزی شود، اختلالی که بر تون، حرکت و هماهنگی عضلات تأثیر میگذارد (باکس 12-4).
CLINICAL BOX 12.4
Cerebral Palsy
Cerebral palsy (CP) is a term used to describe any one of several nonprogressive neurologic disorders that occur before or during childbirth or during early childhood. Prenatal factors, including exposure of the developing brain to hypoxia, infections, or toxins, may account for 70-80% of cases of CP. Typical symptoms of the disorder include spasticity, ataxia, deficits in fine motor control, and abnormal gait (crouched or “scissored gait”). Sensory deficits including loss of vision and hearing as well as learning difficulties and seizures often occur in children with CP. In developed countries, the prevalence of CP is 2-2.5 cases per 1000 live births; however, the incidence of CP in children who are born prematurely is much higher compared with children born at term. Based on differences in the resting tone in muscles and the limbs involved, CP is classified into different groups. The most prevalent type is spastic CP that is characterized by spasticity, hyperreflexia, clonus, and a positive Babinski sign. These are all signs of damage to the corticospinal tract (Clinical Box 12–5). Dyskinetic CP is characterized by abnormal involuntary movements (chorea and athetosis) and may reflect damage to extrapyramidal motor areas. It is not uncommon to have signs of both types of CP. The rarest type is hypotonic CP that presents with truncal and extremity hypotonia, hyperreflexia, and persistent primitive reflexes.
جعبه بالینی 12.4
فلج مغزی
فلج مغزی (CP) اصطلاحی است که برای توصیف هر یک از چندین اختلال عصبی غیر پیشرونده که قبل یا در حین زایمان یا در اوایل دوران کودکی رخ میدهد، استفاده میشود. عوامل پیش از تولد، از جمله قرار گرفتن مغز در حال رشد در معرض هیپوکسی، عفونتها یا سموم، ممکن است 70 تا 80 درصد از موارد CP را تشکیل دهند. علائم معمول این اختلال عبارتند از اسپاستیسیتی، آتاکسی، نقص در کنترل حرکتی ظریف و راه رفتن غیرطبیعی (راه رفتن خمیده یا “قیچی شده”). نقایص حسی از جمله از دست دادن بینایی و شنوایی و همچنین مشکلات یادگیری و تشنج اغلب در کودکان مبتلا به CP رخ میدهد. در کشورهای توسعه یافته، شیوع CP 2-2.5 مورد در هر 1000 تولد زنده است. با این حال، بروز CP در کودکانی که نارس به دنیا میآیند در مقایسه با کودکانی که در ترم متولد میشوند بسیار بیشتر است. بر اساس تفاوت در تون استراحت در عضلات و اندامهای درگیر، CP به گروههای مختلف طبقه بندی میشود. شایع ترین نوع CP اسپاستیک است که با اسپاستیسیتی،هایپررفلکسی، کلونوس و علامت بابینسکی مثبت مشخص میشود. اینها همه نشانههای آسیب به دستگاه قشر نخاعی هستند (باکس بالینی 12-5). CP دیسکینتیک با حرکات غیر ارادی غیر طبیعی (کره و آتتوز) مشخص میشود و ممکن است منعکس کننده آسیب به نواحی حرکتی خارج هرمیباشد. وجود علائم هر دو نوع CP غیر معمول نیست. نادرترین نوع CP هیپوتونیک است که با هیپوتونی تنه و اندام،هایپررفلکسی و رفلکسهای اولیه پایدار تظاهر میکند.
THERAPEUTIQ HIGHLIGHTS
There is no cure for CP. Treatment often includes physical and occupational therapy. Botulinum toxin injections into affected muscles have been used to reduce muscle spasticity, especially in the gastrocnemius muscle. Other drugs used to treat muscle spasticity in patients with CP include diazepam (a benzodiazepine that binds to the GABA receptor), baclofen (an agonist at presynaptic GABA receptors in the spinal cord), and dantrolene (a direct muscle relaxant). Various surgeries have been used to treat CP, including selective dorsal rhizotomy (section of the dorsal roots) and tenotomy (severing the tendon in the gastrocnemius muscles).
نکات برجسته درمانی
هیچ درمانی برای CP وجود ندارد. درمان اغلب شامل فیزیوتراپی و کاردرمانی است. تزریق سم بوتولینوم به عضلات آسیب دیده برای کاهش اسپاستیسیته عضلانی به ویژه در عضله گاستروکنمیوس استفاده شده است. سایر داروهای مورد استفاده برای درمان اسپاستیسیته عضلانی در بیماران مبتلا به CP عبارتند از: دیازپام (یک بنزودیازپین که به گیرنده GABA متصل میشود)، باکلوفن (آگونیست گیرندههای پیش سیناپسی GABA در نخاع)، و دانترولن (یک شل کننده مستقیم عضلانی). جراحیهای مختلفی برای درمان CP استفاده شده است، از جمله ریزوتومیانتخابی پشتی (برشی از ریشههای پشتی) و تنوتومی(بریدن تاندون در عضلات گاستروکنمیوس).
Figure 12-6 shows the general motor control scheme with the commands for voluntary movement originating in cortical association areas. The movements are planned in the cortex, the basal ganglia, and the lateral portions of the cerebellar hemispheres, as indicated by increased electrical activity before the movement. The basal ganglia and cerebellum funnel information to the premotor and motor cortex by way of the thalamus. Motor commands from the motor cortex are relayed predominantly via the corticospinal tracts to the spinal cord and the corresponding corticobulbar tracts to motor neurons in the brainstem. However, collaterals from these pathways and a few direct connections from the motor cortex end on brainstem nuclei that project to motor neurons in the brainstem and spinal cord. These pathways can also mediate voluntary movement. Movement sets up alterations in sensory input from the special senses and from muscles, tendons, joints, and the skin. This feedback information that adjusts and smooths movement is relayed directly to the motor cortex and spinocerebellum. The spinocerebellum then projects to the brainstem. The main brainstem pathways that are concerned with posture and coordination are the rubrospinal, reticulospinal, tectospinal, and vestibulospinal tracts.
شکل 12-6 طرح کلی کنترل حرکتی را با دستورات حرکت ارادی که از نواحی انجمن قشر مغز منشأ میگیرد نشان میدهد. حرکات در قشر، عقدههای قاعده ای و قسمتهای جانبی نیمکرههای مخچه برنامه ریزی میشوند که با افزایش فعالیت الکتریکی قبل از حرکت مشخص میشود. عقدههای قاعده ای و مخچه اطلاعات را از طریق تالاموس به قشر پیش حرکتی و حرکتی میرسانند. دستورات حرکتی از قشر حرکتی عمدتاً از طریق مجاری قشر نخاعی به نخاع و دستگاههای کورتیکوبولبار مربوطه به نورونهای حرکتی در ساقه مغز منتقل میشوند. با این حال، وثیقههای این مسیرها و چند اتصال مستقیم از قشر حرکتی به هستههای ساقه مغز ختم میشوند که به نورونهای حرکتی در ساقه مغز و نخاع میرسند. این مسیرها همچنین میتوانند حرکت ارادی را واسطه کنند. حرکت باعث ایجاد تغییراتی در ورودی حسی از حواس خاص و عضلات، تاندونها، مفاصل و پوست میشود. این اطلاعات بازخوردی که حرکت را تنظیم و هموار میکند، مستقیماً به قشر حرکتی و مخچه نخاعی منتقل میشود. سپس مخچه نخاعی به سمت ساقه مغز پیش میرود. مسیرهای اصلی ساقه مغز که به وضعیت و هماهنگی بدن مربوط میشوند، مسیرهای روبروسنخاعی، رتیکولو نخاعی، تکتو نخاعی و دهلیزی هستند.

FIGURE 12-6 Control of voluntary movement. Commands for voluntary movement originate in cortical association areas. The cortex, basal ganglia, and cerebellum work cooperatively to plan movements. Movement executed by the cortex is relayed via the corticospinal tracts and corticobulbar tracts to motor neurons. The cerebellum provides feedback to adjust and smoothen movement.
شکل 12-6 کنترل حرکت ارادی. دستورات برای حرکت داوطلبانه از نواحی انجمن قشری منشأ میگیرند. قشر قشر، عقدههای قاعده ای و مخچه به طور مشترک برای برنامه ریزی حرکات کار میکنند. حرکت اجرا شده توسط قشر از طریق مجاری قشر نخاعی و دستگاههای کورتیکوبولبار به نورونهای حرکتی منتقل میشود. مخچه بازخوردی را برای تنظیم و صاف کردن حرکت فراهم میکند.
MOTOR CORTEX & VOLUNTARY MOVEMENT
PRIMARY MOTOR CORTEX
The reader can refer to Figure 8-8 for the locations of the major cortical regions involved in motor control. The primary motor cortex (M1) is in the precentral gyrus of the frontal lobe, extending into the central sulcus. By means of stimulation experiments in patients undergoing craniotomy under local anesthesia, this region was mapped to show where various parts of the body are represented in the precentral gyrus. Figure 12-7 shows the motor homunculus with the feet at the top of the gyrus and the face at the bottom. The cortical representation of each body part is proportional in size to the skill with which the part is used in fine, voluntary movement. The areas involved in speech and hand movements are especially large in the cortex; use of the pharynx, lips, and tongue to form words and of the fingers and opposable thumbs to manipulate the environment are activities in which humans are especially skilled.
قشر حرکتی و حرکت داوطلبانه
قشر موتور اولیه
خواننده میتواند به شکل 8-8 برای مکانهای نواحی اصلی قشر درگیر در کنترل حرکتی مراجعه کند. قشر حرکتی اولیه (M1) در شکنج پیش مرکزی لوب فرونتال قرار دارد و تا شیار مرکزی گسترش مییابد. با استفاده از آزمایشهای تحریکی در بیمارانی که تحت بیحسی موضعی تحت کرانیوتومیقرار میگیرند، این منطقه برای نشان دادن مکانهای مختلف بدن در شکنج پیش مرکزی ترسیم شد. شکل 12-7 موتور همونکولوس را با پاها در بالای شکنج و صورت در پایین نشان میدهد. نمایش قشری هر قسمت از بدن متناسب با مهارتی است که از آن قسمت در حرکات خوب و ارادی استفاده میشود. نواحی درگیر در گفتار و حرکات دست به ویژه در قشر بزرگ است. استفاده از حلق، لبها و زبان برای شکل دادن کلمات و از انگشتان دست و شست مخالف برای دستکاری محیط، فعالیتهایی هستند که انسانها در آن مهارت ویژه ای دارند.

FIGURE 12-7 Motor homunculus. The figure represents, on a coronal section of the precentral gyrus, the location of the cortical representation of the various parts. The size of the various parts is proportional to the cortical area devoted to them. Compare with Figure 8-9 which shows the sensory homunculus. (Reproduced with permission from Penfield W, Rasmussen G: The Cerebral Cortex of Man. Macmillan, 1950.)
شکل 12-7 موتور هومونکولوس. شکل، بر روی یک بخش تاجی از شکنج پیش مرکزی، محل نمایش قشر قسمتهای مختلف را نشان میدهد. اندازه قسمتهای مختلف متناسب با ناحیه قشری است که به آنها اختصاص داده شده است. با شکل 8-9 مقایسه کنید که هومونکولوس حسی را نشان میدهد. (تکثیر شده با اجازه Penfield W, Rasmussen G: The Ceebral Cortex of Man. Macmillan, 1950.)
Modern brain imaging techniques such as positron emission tomography (PET) and functional magnetic resonance imaging (fMRI) can be used to map the cortex to identify motor areas. Figure 12-8 shows activation of the hand area of the motor cortex while repetitively squeezing a ball with either the right or left hand.
تکنیکهای مدرن تصویربرداری مغز مانند توموگرافی گسیل پوزیترون (PET) و تصویربرداری تشدید مغناطیسی عملکردی (fMRI) میتواند برای نقشهبرداری از قشر مغز برای شناسایی نواحی حرکتی استفاده شود. شکل 12-8 فعال شدن ناحیه دست قشر حرکتی را در حین فشردن مکرر توپ با دست راست یا چپ نشان میدهد.

FIGURE 12-8 Hand area of motor cortex demonstrated by functional magnetic resonance imaging (fMRI) in a 7-year-old boy. Changes in signal intensity, measured using a method called echoplanar MRI, result from changes in the flow, volume, and oxygenation of the blood. The child was instructed to repetitively squeeze a foam-rubber ball at the rate of two to four squeezes per second with the right or left hand. Changes in cortical activity with the ball in the right hand are shown in black. Changes in cortical activity with the ball in the left hand are shown in white. (Data from Novotny EJ, et al: Functional magnetic resonance imaging [fMRI] in pediatric epilepsy. Epilepsia 1994;35(Supp 8):36.)
شکل 12-8 ناحیه دستی قشر حرکتی که با تصویربرداری رزونانس مغناطیسی عملکردی (fMRI) در یک پسر 7 ساله نشان داده شده است. تغییرات در شدت سیگنال، که با استفاده از روشی به نام اکوپلانار ام آر آی اندازه گیری میشود، ناشی از تغییرات در جریان، حجم و اکسیژن رسانی خون است. به کودک دستور داده شد که به طور مکرر یک توپ فوم لاستیکی را با سرعت دو تا چهار فشار در ثانیه با دست راست یا چپ فشار دهد. تغییرات در فعالیت قشر مغز با توپ در دست راست به رنگ سیاه نشان داده شده است. تغییرات در فعالیت قشر مغز با توپ در دست چپ به رنگ سفید نشان داده شده است. (دادهها از Novotny EJ و همکاران: تصویربرداری تشدید مغناطیسی عملکردی [fMRI] در صرع کودکان. Epilepsia 1994; 35 (Supp 8):36.)
The cells in the cortical motor areas are arranged in a columnar organization. M1 neurons represent movements of groups of muscles for different tasks. Neurons in several cortical columns project to the same muscle; moreover, the cells in each column receive sensory input from the peripheral area in which they produce movement, providing the basis for feedback control of movement. Some of this input may be direct and some is relayed from other parts of the cortex.
سلولها در نواحی حرکتی قشر مغز در یک سازمان ستونی مرتب شده اند. نورونهای M1 حرکات گروههایی از ماهیچهها را برای کارهای مختلف نشان میدهند. نورونها در چندین ستون قشر مغز به یک عضله منعکس میشوند. علاوه بر این، سلولهای هر ستون ورودی حسی را از ناحیه محیطی که در آن حرکت تولید میکنند، دریافت میکنند و مبنایی را برای کنترل بازخورد حرکت فراهم میکنند. بخشی از این ورودی ممکن است مستقیم باشد و برخی از قسمتهای دیگر قشر مغز منتقل شود.
SUPPLEMENTARY MOTOR AREA
The supplementary motor area is on and above the superior bank of the cingulate sulcus on the medial side of the hemisphere. It projects to M1 and contains a map of the body; but it is less precise than in M1. The supplementary motor area is involved in organizing or planning motor sequences, while M1 executes the movements.
منطقه موتور تکمیلی
ناحیه حرکتی تکمیلی روی و بالای کرانه فوقانی شیار سینگولیت در سمت داخلی نیمکره قرار دارد. آن را به M1 پروژه میدهد و شامل یک نقشه از بدن. اما نسبت به M1 دقت کمتری دارد. ناحیه حرکتی تکمیلی در سازماندهی یا برنامه ریزی توالیهای حرکتی نقش دارد، در حالی که M1 حرکات را اجرا میکند.
When human subjects count to themselves without speaking, the motor cortex is quiescent, but when they speak the numbers aloud as they count, blood flow increases in M1 and the supplementary motor area. Thus, both regions are involved in voluntary movement when the movements being performed are complex and involve planning.
وقتی سوژههای انسانی بدون صحبت کردن برای خودشان حساب میکنند، قشر حرکتی ساکت است، اما وقتی اعداد را در حین شمارش با صدای بلند صحبت میکنند، جریان خون در M1 و ناحیه حرکتی مکمل افزایش مییابد. بنابراین، زمانی که حرکات در حال انجام پیچیده و مستلزم برنامه ریزی هستند، هر دو منطقه درگیر حرکت داوطلبانه هستند.
PREMOTOR CORTEX
The premotor cortex is located anterior to the precentral gyrus, on the lateral and medial cortical surface; it contains a somatotopic map. This region receives input from sensory regions of the parietal cortex and projects to M1, the spinal cord, and the brainstem reticular formation. This region is concerned with setting posture at the start of a planned movement and with getting the individual prepared to move. It is most involved in control of proximal limb muscles needed to orient the body for movement.
قشر قبل از حرکت
قشر پیش حرکتی در جلوی شکنج پیش مرکزی، در سطح قشر جانبی و میانی قرار دارد. این شامل یک نقشه جسمانی است. این ناحیه ورودی را از نواحی حسی قشر جداری دریافت میکند و به M1، نخاع و تشکیل مشبک ساقه مغز میپردازد. این منطقه با تنظیم وضعیت در شروع یک حرکت برنامه ریزی شده و آماده کردن فرد برای حرکت مرتبط است. بیشتر در کنترل عضلات پروگزیمال اندام مورد نیاز برای جهت دهی بدن برای حرکت نقش دارد.
POSTERIOR PARIETAL CORTEX
The somatic sensory area and related portions of the posterior parietal lobe project to the premotor cortex. Lesions of the somatic sensory area cause defects in motor performance that are characterized by inability to execute learned sequences of movements such as eating with a knife and fork. Some of the neurons are concerned with aiming the hands toward an object and manipulating it, whereas other neurons are concerned with hand-eye coordination. As described below, neurons in this posterior parietal cortex contribute to the descending pathways involved in motor control.
قشر جداری خلفی
ناحیه حسی جسمیو بخشهای مربوط به لوب جداری خلفی به قشر پیش حرکتی میرود. ضایعات ناحیه حسی جسمیباعث نقص در عملکرد حرکتی میشود که با ناتوانی در اجرای توالیهای آموخته شده از حرکات مانند غذا خوردن با چاقو و چنگال مشخص میشود. برخی از نورونها با هدف گرفتن دستها به سمت یک شی و دستکاری آن سروکار دارند، در حالی که سایر نورونها به هماهنگی چشم و دست میپردازند. همانطور که در زیر توضیح داده شد، نورونها در این قشر جداری خلفی به مسیرهای نزولی درگیر در کنترل حرکت کمک میکنند.
PLASTICITY
A striking discovery made possible by PET and fMRI is that the motor cortex shows the same kind of plasticity as already described for the sensory cortex in Chapter 8. For example, the finger areas of the contralateral motor cortex enlarge as a pattern of rapid finger movement is learned with the fingers of one hand; this change is detectable at 1 week and maximal at 4 weeks. Cortical areas of output to other muscles also increase in size when motor learning involves these muscles. When a small focal ischemic lesion is produced in the hand area of the motor cortex of monkeys, the hand area may reappear, with return of motor function, in an adjacent undamaged part of the cortex. Thus, the maps of the motor cortex are not immutable; they change with experience.
پلاستیسیته
یک کشف قابل توجه که توسط PET و fMRI ممکن شده است این است که قشر حرکتی همان شکل پذیری را نشان میدهد که قبلاً برای قشر حسی در فصل 8 توضیح داده شد. این تغییر در 1 هفته و حداکثر در 4 هفته قابل تشخیص است. زمانی که یادگیری حرکتی این ماهیچهها را درگیر میکند، نواحی خروجی قشر به سایر عضلات نیز افزایش مییابد. هنگامیکه یک ضایعه ایسکمیک کانونی کوچک در ناحیه دست قشر حرکتی میمونها ایجاد میشود، ناحیه دست ممکن است با بازگشت عملکرد حرکتی، در قسمت مجاور آسیب دیده قشر قشر ظاهر شود. بنابراین، نقشههای قشر حرکتی تغییرناپذیر نیستند. آنها با تجربه تغییر میکنند.
CONTROL OF AXIAL & DISTAL MUSCLES
Within the brainstem and spinal cord, pathways and neurons that are concerned with the control of skeletal muscles of the trunk (axial) and proximal portions of the limbs are located medially or ventrally, whereas pathways and neurons that are concerned with the control of skeletal muscles in the distal portions of the limbs are located laterally. The axial muscles are concerned with postural adjustments and gross movements, whereas the distal limb muscles mediate fine, skilled movements. Neurons in the medial portion of the ventral horn innervate proximal limb muscles, particularly the flexors, and lateral ventral horn neurons innervate distal limb muscles. Similarly, the ventral corticospinal tract and medial descending brainstem pathways (tectospinal, reticulospinal, and vestibulospinal tracts) are concerned with adjustments of proximal muscles and posture, and the lateral corticospinal and rubrospinal tracts are concerned with distal limb muscles and, particularly in the case of the lateral corticospinal tract, with skilled voluntary movements.
کنترل عضلات محوری و دیستال
در داخل ساقه مغز و نخاع، مسیرها و نورونهایی که مربوط به کنترل ماهیچههای اسکلتی تنه (محوری) و قسمتهای پروگزیمال اندامها هستند، به صورت داخلی یا شکمیقرار دارند، در حالی که مسیرها و نورونهایی که مربوط به کنترل عضلات اسکلتی در قسمتهای انتهایی اندامها هستند، در قسمت بعدی قرار دارند. ماهیچههای محوری با تنظیمات وضعیتی و حرکات درشت سروکار دارند، در حالی که عضلات اندام انتهایی، حرکات ظریف و ماهرانه را انجام میدهند. نورونها در بخش داخلی شاخ شکمیعضلات پروگزیمال اندام، به ویژه خم کنندهها را عصب دهی میکنند و نورونهای شاخ شکمیجانبی عضلات انتهای اندام را عصب دهی میکنند. به طور مشابه، مسیرهای قشر نخاعی شکمیو مسیرهای نزولی ساقه مغز میانی (تکتو نخاعی، شبکه نخاعی، و دهلیزی نخاعی) مربوط به تنظیم عضلات پروگزیمال و وضعیت بدن هستند، و دستگاههای قشر نخاعی و روبروسنال جانبی مربوط به اندامهای انتهایی اندامهای انتهایی، عضلات قشر نخاعی هستند حرکات داوطلبانه ماهرانه
CORTICOSPINAL & CORTICOBULBAR TRACTS
The somatotopic organization just described for the motor cortex continues throughout the pathways from the cortex to the motor neurons. The axons of neurons from the motor cortex that project to spinal motor neurons form the corticospinal tracts, a large bundle of about 1 million fibers. About 80% of these fibers cross the midline in the medullary pyramids to form the lateral corticospinal tract (Figure 12-9). The remaining 20% make up the ventral corticospinal tract, which does not cross the midline until it reaches the level of the spinal cord at which it terminates. Lateral corticospinal tract neurons make monosynaptic connections to motor neurons, especially those concerned with skilled movements. Many corticospinal tract neurons also synapse on spinal interneurons antecedent to motor neurons; this indirect pathway is important in coordinating groups of muscles.
دستگاههای قشر نخاعی و کورتیکوبولبار
سازمان سوماتوتوپیک که برای قشر حرکتی توضیح داده شد در سراسر مسیرها از قشر به نورونهای حرکتی ادامه دارد. آکسونهای نورونهای قشر حرکتی که به سمت نورونهای حرکتی نخاعی حرکت میکنند، مجاری قشر نخاعی را تشکیل میدهند، یک بسته بزرگ از حدود 1 میلیون فیبر. حدود 80% از این الیاف از خط میانی در هرمهای مدولاری عبور میکنند تا دستگاه قشر نخاعی جانبی را تشکیل دهند (شکل 12-9). 20 درصد باقیمانده دستگاه قشر نخاعی شکمیرا تشکیل میدهد که تا زمانی که به سطح نخاعی که در آن ختم میشود نرسد از خط وسط عبور نمیکند. نورونهای مجرای قشر نخاعی جانبی اتصالات تک سیناپسی را با نورونهای حرکتی، به ویژه آنهایی که با حرکات ماهرانه مرتبط هستند، ایجاد میکنند. بسیاری از نورونهای دستگاه قشر نخاعی نیز پیش از نورونهای حرکتی بر روی نورونهای داخلی نخاعی سیناپس میشوند. این مسیر غیرمستقیم در هماهنگی گروههای ماهیچه ای مهم است.

FIGURE 12-9 The corticospinal tracts. This tract originates in the precentral gyrus and passes through the internal capsule. Most fibers decussate in the pyramids and descend in the lateral white matter of the spinal cord to form the lateral division of the tract which can make monosynaptic connections with spinal motor neurons. The ventral division of the tract remains uncrossed until reaching the spinal cord where axons terminate on spinal interneurons antecedent to motor neurons.
شکل 12-9 مجاری قشر نخاعی. این مسیر از شکنج پیش مرکزی منشا گرفته و از کپسول داخلی عبور میکند. بیشتر الیاف در اهرام جدا میشوند و در ماده سفید جانبی نخاع فرود میآیند تا تقسیم جانبی دستگاه را تشکیل دهند که میتواند اتصالات مونوسیناپسی را با نورونهای حرکتی نخاعی ایجاد کند. بخش شکمیدستگاه تا رسیدن به نخاع، جایی که آکسونها به نورونهای بینالمللی نخاعی پیش از نورونهای حرکتی ختم میشوند، بدون تلاقی باقی میماند.
The trajectory from the cortex to the spinal cord passes through the corona radiata to the posterior limb of the internal capsule. Within the midbrain they traverse the cerebral peduncle and the basilar pons until they reach the medullary pyramids on their way to the spinal cord.
مسیر از قشر به نخاع از طریق تاج رادیاتا به اندام خلفی کپسول داخلی میگذرد. در داخل مغز میانی، آنها از ساقه مغزی و حفرههای قاعدهای عبور میکنند تا زمانی که در مسیر خود به سمت نخاع به اهرام مدولاری برسند.
The corticobulbar tract is composed of the fibers that pass from the motor cortex to motor neurons in the trigeminal, facial, and hypoglossal nuclei. Corticobulbar neurons end either directly on the cranial nerve nuclei or on their antecedent interneurons within the brainstem. Their axons traverse through the genu of the internal capsule, the cerebral peduncle (medial to corticospinal tract neurons), to descend with corticospinal tract fibers in the pons and medulla.
دستگاه کورتیکوبولبار از فیبرهایی تشکیل شده است که از قشر حرکتی به نورونهای حرکتی در هستههای سه قلو، صورت و هیپوگلاس میگذرد. نورونهای کورتیکوبولبار یا مستقیماً به هستههای عصبی جمجمه یا به نورونهای پیشین خود در ساقه مغز ختم میشوند. آکسونهای آنها از جنس کپسول داخلی، دمگل مغزی (نرونهای دستگاه داخلی به قشر نخاعی) عبور میکنند تا همراه با رشتههای دستگاه قشر نخاعی در پونز و مدولا پایین بیایند.
The motor system can be divided into lower and upper motor neurons. Lower motor neurons refer to the spinal and cranial motor neurons that directly innervate skeletal muscles. Upper motor neurons are those in the cortex and brainstem that activate the lower motor neurons. The pathophysiologic responses to damage to lower and upper motor neurons are very distinctive (Clinical Box 12-5).
سیستم حرکتی را میتوان به نورونهای حرکتی تحتانی و فوقانی تقسیم کرد. نورونهای حرکتی تحتانی به نورونهای حرکتی نخاعی و جمجمه ای اشاره دارند که مستقیماً عضلات اسکلتی را عصب دهی میکنند. نورونهای حرکتی بالایی آنهایی هستند که در قشر و ساقه مغز قرار دارند که نورونهای حرکتی تحتانی را فعال میکنند. پاسخهای پاتوفیزیولوژیک به آسیب به نورونهای حرکتی تحتانی و فوقانی بسیار متمایز است (باکس بالینی 12-5).
CLINICAL BOX 12.5
Lower versus Upper Motor Neuron Damage
Lower motor neurons are those whose axons terminate on skeletal muscles. Damage to these neurons is associated with flaccid paralysis, muscular atrophy, fasciculations (visible muscle twitches that appear as flickers under the skin), hypotonia (decreased muscle tone), and hyporeflexia or areflexia. An example of a disease that leads to lower motor neuron damage is ALS. “Amyotrophic” means “no muscle nourishment” and describes the atrophy that muscles undergo because of disuse. “Sclerosis” refers to the hardness felt when a pathologist examines the spinal cord on autopsy; the hardness is due to proliferation of astrocytes and scarring of the lateral columns of the spinal cord. ALS is a selective, progressive degeneration of a-motor neurons. This fatal disease is also known as Lou Gehrig disease in remembrance of a famous American baseball player who died of ALS. The worldwide annual incidence of ALS has been estimated to be 0.5-3 cases per 100,000 people. The disease has no racial, socioeconomic, or ethnic boundaries. The life expectancy of ALS patients is usually 3-5 years after diagnosis. ALS is most commonly diagnosed in middle age and affects men more often than women. Most cases of ALS are sporadic in origin; but 5-10% of the cases have a familial link. Possible causes include viruses, neurotoxins, heavy metals, DNA defects (especially in familial ALS), immune system abnormalities, and enzyme abnormalities. About 40% of the familial cases have a mutation in the gene for Cu/Zn superoxide dismutase (SOD-1) on chromosome 21. SOD is a free radical scavenger that reduces oxidative stress. A defective SOD-1 gene permits free radicals to accumulate and kill neurons. An increase in the excitability of deep cerebellar nuclei due to the inhibition of small- conductance calcium-activated potassium (SK) channels may contribute to the development of cerebellar ataxia.
جعبه بالینی 12.5
آسیب نورون موتور پایین در مقابل بالا
نورونهای حرکتی تحتانی آنهایی هستند که آکسون آنها به ماهیچههای اسکلتی ختم میشود. آسیب به این نورونها با فلج شل، آتروفی عضلانی، فاسیکولاسیون (انقباضات عضلانی قابل مشاهده که به صورت سوسو زدن زیر پوست ظاهر میشود)، هیپوتونی (کاهش تون عضلانی) و هیپورفلکسی یا آرفلکسی همراه است. نمونه ای از بیماری که منجر به آسیب نورون حرکتی پایین تر میشود ALS است. “آمیوتروفیک” به معنای “عدم تغذیه عضلانی” است و آتروفی را توصیف میکند که عضلات به دلیل استفاده نکردن دچار آن میشوند. “اسکلروز” به سختی اطلاق میشود که وقتی پاتولوژیست نخاع را در کالبد شکافی بررسی میکند. سختی به دلیل تکثیر آستروسیتها و زخم شدن ستونهای جانبی نخاع است. ALS یک انحطاط انتخابی و پیشرونده نورونهای حرکتی است. این بیماری کشنده به یاد یک بازیکن مشهور بیسبال آمریکایی که بر اثر بیماری ALS درگذشت، به بیماری لو گریگ نیز معروف است. بروز سالانه ALS در سراسر جهان 0.5-3 مورد در هر 100000 نفر تخمین زده شده است. این بیماری مرزهای نژادی، اجتماعی-اقتصادی یا قومیتی ندارد. امید به زندگی بیماران ALS معمولاً 3-5 سال پس از تشخیص است. ALS بیشتر در میانسالی تشخیص داده میشود و مردان را بیشتر از زنان مبتلا میکند. اکثر موارد ALS منشأ پراکنده دارند. اما 5 تا 10 درصد موارد دارای پیوند خانوادگی هستند. علل احتمالی عبارتند از ویروسها، نوروتوکسینها، فلزات سنگین، نقصهای DNA (بهویژه در ALS خانوادگی)، ناهنجاریهای سیستم ایمنی، و ناهنجاریهای آنزیمی. حدود 40 درصد از موارد خانوادگی دارای جهش در ژن سوپراکسید دیسموتاز مس/رو (SOD-1) در کروموزوم 21 هستند. SOD یک پاک کننده رادیکالهای آزاد است که استرس اکسیداتیو را کاهش میدهد. یک ژن معیوب SOD-1 به رادیکالهای آزاد اجازه میدهد تا نورونها را جمع کنند و از بین ببرند. افزایش تحریک پذیری هستههای عمیق مخچه به دلیل مهار کانالهای پتاسیم فعال شده با کلسیم (SK) با رسانایی کوچک ممکن است به ایجاد آتاکسی مخچه کمک کند.
Upper motor neurons typically refer to corticospinal tract neurons that innervate spinal motor neurons, but they can also include brainstem neurons that control spinal motor neurons. Damage to these neurons initially causes muscles to become weak and flaccid but eventually leads to spasticity, hypertonia (increased resistance to passive movement), hyperactive stretch reflexes, and abnormal plantar extensor reflex (positive Babinski sign). The Babinski sign is dorsiflexion of the great toe and fanning of the other toes when the lateral aspect of the sole of the foot is scratched. In adults, the normal response to this stimulation is plantar flexion in all the toes. The Babinski sign is of value in the localization of disease processes, but its physiologic significance is unknown. In infants whose corticospinal tracts are not well developed, dorsiflexion of the great toe and fanning of the other toes is the natural response to stimuli applied to the sole of the foot.
نورونهای حرکتی فوقانی معمولاً به نورونهای دستگاه قشر نخاعی اشاره میکنند که نورونهای حرکتی نخاعی را عصب دهی میکنند، اما آنها همچنین میتوانند شامل نورونهای ساقه مغز باشند که نورونهای حرکتی نخاعی را کنترل میکنند. آسیب به این نورونها در ابتدا باعث ضعیف شدن و شل شدن ماهیچهها میشود اما در نهایت منجر به اسپاستیسیتی، هیپرتونی (افزایش مقاومت در برابر حرکت غیرفعال)، رفلکسهای کششی بیش فعال و رفلکس غیر طبیعی اکستانسور کف پا (علامت بابینسکی مثبت) میشود. علامت بابینسکی خم شدن انگشت شست پا و فن کردن انگشتان دیگر در هنگام خراشیدگی قسمت جانبی کف پا است. در بزرگسالان، پاسخ طبیعی به این تحریک، خم شدن کف پا در تمام انگشتان پا است. علامت بابینسکی در محلی سازی فرآیندهای بیماری ارزش دارد، اما اهمیت فیزیولوژیکی آن ناشناخته است. در نوزادانی که مجاری قشر نخاعی آنها به خوبی رشد نکرده است، دورسی فلکشن انگشت شست پا و پنکه شدن انگشتان دیگر پاسخ طبیعی به محرکهایی است که روی کف پا اعمال میشود.
THERAPEUTIC HIGHLIGHTS
One of the few drugs shown to modestly slow the progression of ALS is riluzole, a drug that opens the SK channels and may be effective in preventing nerve damage caused by excessive release of the excitatory amino acid, glutamate. Spasticity associated with motor neuron disease can be reduced by the muscle relaxant baclofen (a derivative of GABA); in some cases, a subarachnoid infusion of baclofen is given via an implanted lumbar pump. Spasticity can also be treated with tizanidine, a centrally acting α2- adrenoceptor agonist; its effectiveness is due to increasing presynaptic inhibition of spinal motor neurons. Botulinum toxin is also approved for the treatment of spasticity; this toxin acts by binding to receptors on the cholinergic nerve terminals to decrease the release of acetylcholine, causing neuromuscular blockade.
نکات برجسته درمانی
یکی از معدود داروهایی که نشان داده شده است که پیشرفت ALS را به طور متوسط کاهش میدهد، ریلوزول است، دارویی که کانالهای SK را باز میکند و ممکن است در جلوگیری از آسیب عصبی ناشی از آزادسازی بیش از حد اسید آمینه تحریک کننده، گلوتامات، موثر باشد. اسپاستیسیته مرتبط با بیماری نورون حرکتی را میتوان با شل کننده عضلانی باکلوفن (مشتق شده از GABA) کاهش داد. در برخی موارد، انفوزیون ساب عنکبوتیه باکلوفن از طریق پمپ کمری کاشته شده انجام میشود. اسپاستیسیتی را میتوان با تیزانیدین، یک آگونیست گیرنده آلفا2- آدرنرژیک با اثر مرکزی نیز درمان کرد. اثربخشی آن به دلیل افزایش مهار پیش سیناپسی نورونهای حرکتی ستون فقرات است. سم بوتولینوم نیز برای درمان اسپاستیسیتی تایید شده است. این سم با اتصال به گیرندههای پایانههای عصبی کولینرژیک برای کاهش آزادسازی استیل کولین عمل میکند و باعث مسدود شدن عصبی عضلانی میشود.
ORIGINS OF CORTICOSPINAL & CORTICOBULBAR TRACTS
Corticospinal and corticobulbar tract neurons are pyramidal shaped and located in layer V of the cerebral cortex (see Chapter 14). The cortical areas from which these tracts originate were identified on the basis of electrical stimulation that produced prompt discrete movement. About 31% of the corticospinal tract neurons are from M1. The premotor cortex and supplementary motor cortex account for 29% of the corticospinal tract neurons. The other 40% of corticospinal tract neurons originate in the parietal lobe and primary somatosensory area in the postcentral gyrus. The corticospinal and corticobulbar system is the primary pathway for the initiation of skilled voluntary movement.
منشاء دستگاههای قشر نخاعی و قشر مغزی
نورونهای دستگاه قشر نخاعی و کورتیکوبولبار هرمیشکل هستند و در لایه V قشر مغز قرار دارند (به فصل 14 مراجعه کنید). نواحی قشر مغزی که این مجاری از آن منشا میگیرند بر اساس تحریک الکتریکی که حرکت گسسته سریع ایجاد میکند، شناسایی شدند. حدود 31 درصد از نورونهای دستگاه قشر نخاعی از M1 هستند. قشر پیش حرکتی و قشر حرکتی تکمیلی 29 درصد از نورونهای دستگاه قشر نخاعی را تشکیل میدهند. 40 درصد دیگر نورونهای دستگاه قشر نخاعی از لوب جداری و ناحیه حسی جسمیاولیه در شکنج پس مرکزی منشا میگیرند. سیستم قشر نخاعی و کورتیکوبولبار مسیر اصلی برای شروع حرکت ارادی ماهر است.
BRAINSTEM PATHWAYS INVOLVED IN POSTURE & VOLUNTARY MOVEMENT
As mentioned above, spinal motor neurons are organized so that those innervating the most proximal muscles are located most medially and those innervating the more distal muscles are located more laterally. This organization is also reflected in descending brainstem pathways (Figure 12-10).
مسیرهای ساقه مغز درگیر در وضعیت وضعیت بدن و حرکات داوطلبانه
همانطور که در بالا ذکر شد، نورونهای حرکتی ستون فقرات به گونه ای سازماندهی شده اند که آنهایی که بیشترین عضلات را عصب دهی میکنند در قسمت داخلی قرار دارند و آنهایی که عضلات دیستال تر را عصب دهی میکنند بیشتر در طرفین قرار دارند. این سازمان در مسیرهای نزولی ساقه مغز نیز منعکس میشود (شکل 12-10).

FIGURE 12-10 Medial and lateral descending brainstem pathways involved in motor control. A) Medial pathways (reticulospinal, vestibulospinal, and tectospinal) terminate in ventromedial area of spinal gray matter and control axial and proximal muscles. B) Lateral pathway (rubrospinal) terminates in dorsolateral area of spinal gray matter and controls distal muscles. (Reproduced with permission from Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. New York, NY: McGraw-Hill; 2000.)
شکل 12-10 مسیرهای نزولی داخلی و جانبی ساقه مغز درگیر در کنترل حرکتی. الف) مسیرهای داخلی (شبکه نخاعی، دهلیزی و نخاعی) به ناحیه شکمیماده خاکستری ستون فقرات ختم میشود و عضلات محوری و پروگزیمال را کنترل میکند. ب) مسیر جانبی (rubrospinal) به ناحیه پشتی جانبی ماده خاکستری ستون فقرات ختم میشود و عضلات دیستال را کنترل میکند. (تکثیر شده با اجازه از Kandel ER، Schwartz JH، Jessell TM [ویراستاران]: Principles of Neural Science، ویرایش چهارم نیویورک، نیویورک: McGraw-Hill؛ 2000.)
MEDIAL BRAINSTEM PATHWAYS
The medial brainstem pathways, which work in concert with the ventral corticospinal tract, are the pontine and medullary reticulospinal, vestibulospinal, and tectospinal tracts. These pathways descend in the ipsilateral ventral columns of the spinal cord and terminate predominantly on interneurons and long propriospinal neurons in the ventromedial part of the ventral horn to control axial and proximal muscles. few medial pathway neurons synapse directly on motor neurons controlling axial muscles.
مسیرهای میانی ساقه مغز
مسیرهای ساقه مغز میانی که در هماهنگی با دستگاه قشر نخاعی شکمیکار میکنند، مسیرهای شبکه ای نخاعی پونتین و مدولاری، دهلیزی نخاعی و تکتو نخاعی هستند. این مسیرها در ستونهای شکمیهمان طرف نخاع فرود میآیند و عمدتاً به نورونهای بیناعصاب و نورونهای بلند عمقی نخاعی در قسمت شکمیمیانی شاخ شکمیختم میشوند تا عضلات محوری و پروگزیمال را کنترل کنند. تعداد کمیاز نورونهای مسیر داخلی مستقیماً روی نورونهای حرکتی کنترل کننده عضلات محوری سیناپس میشوند.
The medial and lateral vestibulospinal tracts are involved in vestibular function and are described in Chapter 11. The medial tract originates in the medial and inferior vestibular nuclei and projects bilaterally to cervical spinal motor neurons that control neck musculature. The lateral tract originates in the lateral vestibular nuclei and projects ipsilaterally to neurons at all spinal levels. It activates motor neurons to antigravity muscles (eg, proximal limb extensors) to control posture and balance.
مجاری دهلیزی نخاعی داخلی و جانبی در عملکرد دهلیزی نقش دارند و در فصل 11 توضیح داده شده است. مسیر داخلی از هستههای دهلیزی داخلی و تحتانی سرچشمه میگیرد و به صورت دو طرفه به نورونهای حرکتی ستون فقرات گردنی که عضلات گردن را کنترل میکنند، میرود. مسیر جانبی از هستههای دهلیزی جانبی منشا میگیرد و به صورت همان طرف به نورونها در تمام سطوح ستون فقرات میرسد. این نورونهای حرکتی را به عضلات ضد جاذبه (به عنوان مثال، بازکننده اندامهای پروگزیمال) برای کنترل وضعیت و تعادل فعال میکند.
The pontine and medullary reticulospinal tracts project to all spinal levels. They are involved in the maintenance of posture and in modulating muscle tone, especially via an input to y-motor neurons. Pontine reticulospinal neurons are primarily excitatory and medullary reticulospinal neurons are primarily inhibitory. The tectospinal tract originates in the superior colliculus of the midbrain and projects to the contralateral cervical spinal cord to control head and eye movements.
مجاری شبکهای نخاعی پونتین و مدولاری به تمام سطوح نخاعی پیش میروند. آنها در حفظ وضعیت بدن و تعدیل تون عضلانی، به ویژه از طریق ورودی به نورونهای حرکتی y، نقش دارند. نورونهای شبکهای نخاعی پونتین عمدتاً تحریککننده هستند و نورونهای شبکهای نخاعی مدولاری عمدتاً مهارکننده هستند. مسیر تکتو نخاعی از کولیکولوس فوقانی مغز میانی سرچشمه میگیرد و برای کنترل حرکات سر و چشم به سمت نخاع گردنی مقابل میرود.
LATERAL BRAINSTEM PATHWAY
The main control of distal muscles arises from the lateral corticospinal tract, but neurons within the red nucleus of the midbrain cross the midline and project to interneurons in the dorsolateral part of the spinal ventral horn to also influence motor neurons that control distal limb muscles. This rubrospinal tract excites flexor motor neurons and inhibits extensor motor neurons. This pathway is not very prominent in healthy humans, but it may play a role in the posture typical of decorticate rigidity (see below).
مسیر جانبی ساقه مغز
کنترل اصلی عضلات دیستال از دستگاه کورتیکو نخاعی جانبی ناشی میشود، اما نورونهای درون هسته قرمز مغز میانی از خط وسط عبور میکنند و به سمت نورونهای درونی در قسمت پشتی جانبی شاخ شکمینخاعی حرکت میکنند تا بر نورونهای حرکتی که ماهیچههای انتهایی اندام را کنترل میکنند نیز تأثیر بگذارند. این دستگاه روبروسنخاعی نورونهای حرکتی فلکسور را تحریک میکند و نورونهای حرکتی اکستانسور را مهار میکند. این مسیر در انسانهای سالم خیلی برجسته نیست، اما ممکن است در وضعیت حالت معمولی سفتی تزئینی نقش داشته باشد (به زیر مراجعه کنید).
POSTURE-REGULATING SYSTEMS
When damage occurs somewhere along the neural axis, the activities integrated below the injury are released from the control of higher brain centers and often are accentuated. Release of this type, long a cardinal principle in neurology, may be due in some situations to disruption of an inhibitory control by higher neural regions.
سیستمهای تنظیم کننده وضعیت بدن
هنگامیکه آسیب در جایی در امتداد محور عصبی رخ میدهد، فعالیتهای ادغام شده در زیر آسیب از کنترل مراکز بالاتر مغز رها شده و اغلب برجسته میشوند. انتشار این نوع، که مدتها یک اصل اساسی در نورولوژی است، ممکن است در برخی شرایط به دلیل اختلال در کنترل مهاری توسط نواحی عصبی بالاتر باشد.
DECEREBRATION
A complete transection of the brainstem between the superior and inferior colliculi permits the brainstem pathways to function independent of their input from higher brain structures. This midcollicular decerebration is diagramed in Figure 12-11 by the dashed line labeled A. This lesion interrupts all inputs from the cortex (corticospinal and corticobulbar tracts) and red nucleus (rubrospinal tract), primarily to distal muscles of the extremities. The excitatory and inhibitory reticulospinal pathways (primarily to postural extensor muscles) remain intact. The dominance of drive from ascending sensory pathways to the excitatory reticulospinal pathway leads to hyperactivity in extensor muscles in all four extremities (decerebrate rigidity). This resembles what ensues after uncal herniation due to a supratentorial lesion. Uncal herniation can occur in patients with large tumors or a hemorrhage in the cerebral hemisphere. Figure12-12A shows the posture typical of such a patient. Clinical Box 12-6 describes complications related to uncal herniation.
DECEBRATION
قطع کامل ساقه مغز بین کولیکولهای فوقانی و تحتانی به مسیرهای ساقه مغز اجازه میدهد تا مستقل از ورودی آنها از ساختارهای بالاتر مغز عمل کنند. این دسربراسیون میانی کولیکولار در شکل 12-11 با خط چین نشان داده شده با A نشان داده شده است. این ضایعه تمام ورودیهای قشر مغز (دستگاههای قشر نخاعی و کورتیکوبولبار) و هسته قرمز (دستگاه روبروسنخاعی)، عمدتاً به عضلات دیستال اندامها را قطع میکند. مسیرهای تحریکی و مهاری شبکه نخاعی (عمدتاً به عضلات بازکننده وضعیتی) دست نخورده باقی میمانند. تسلط درایو از مسیرهای حسی صعودی به مسیر رتیکولوس نخاعی تحریکی منجر به بیش فعالی در عضلات بازکننده در هر چهار اندام میشود (سفتی decerebrate). این شبیه چیزی است که پس از فتق انکال ناشی از یک ضایعه فوق تنتوریال ایجاد میشود. فتق آنکال میتواند در بیماران مبتلا به تومورهای بزرگ یا خونریزی در نیمکره مغز رخ دهد. شکل 12-12A وضعیت معمول چنین بیمار را نشان میدهد. جعبه بالینی 12-6 عوارض مربوط به فتق ناقص را شرح میدهد.
Decerebrate rigidity is spasticity due to facilitation of the myotatic stretch reflex. That is, the excitatory input from the reticulospinal pathway activates y- motor neurons which indirectly activate a-motor neurons (via la spindle afferent activity). This is called the gamma loop. This was demonstrated by cutting dorsal roots to a limb in midcollicular decerebrate cats (dashed line labeled B in Figure 12-11) that immediately eliminated the hyperactivity of extensor muscles.
سفتی دسربرات اسپاستیسیته ناشی از تسهیل رفلکس کشش میوتاتیک است. یعنی ورودی تحریکی از مسیر شبکه نخاعی نورونهای حرکتی y را فعال میکند که به طور غیرمستقیم نورونهای a-motor را فعال میکند (از طریق فعالیت آوران دوک نخاعی). به این حلقه گاما میگویند. این با بریدن ریشههای پشتی به یک اندام در گربههای دسربرات میانی کولیکولار (خط چین با برچسب B در شکل 12-11) نشان داده شد که بلافاصله بیش فعالی عضلات بازکننده را از بین برد.
Decerebrate rigidity can also lead to direct activation of a-motor neurons. If the anterior lobe of the cerebellum is removed in a decerebrate animal (dashed line labeled C in Figure 12-11), extensor muscle hyperactivity is exaggerated (decerebellate rigidity). This lesion eliminates cortical inhibition of the cerebellar fastigial nucleus and secondarily increases excitation to vestibular nuclei. Subsequent dorsal root section does not reverse the rigidity due to activation of a-motor neurons independent of the gamma loop.
سفتی ضعیف همچنین میتواند منجر به فعال شدن مستقیم نورونهای a-motor شود. اگر لوب قدامیمخچه در یک حیوان دسربره برداشته شود (خط چین با برچسب C در شکل 12-11)، بیش فعالی عضله اکستانسور اغراق آمیز است (سفتی دسربلات). این ضایعه مهار قشری هسته فاستژیال مخچه را از بین میبرد و به طور ثانویه تحریک به هستههای دهلیزی را افزایش میدهد. بخش ریشه پشتی بعدی به دلیل فعال شدن نورونهای a-motor مستقل از حلقه گاما، سفتی را معکوس نمیکند.

FIGURE 12-11 A circuit drawing representing lesions produced in experimental animals to replicate decerebrate and decorticate deficits seen in humans. Bilateral transections are indicated by dashed lines A, B, C, and D. Decerebration is at a midcollicular level (A), decortication is rostral to the superior colliculus, dorsal roots sectioned for one extremity (B), and damage to the anterior lobe of cerebellum (C). The objective was to identify anatomic substrates responsible for decerebrate or decorticate rigidity/posturing seen in humans with lesions that either isolate the forebrain from the brainstem or separate rostral from caudal brainstem and spinal cord. (Reproduced with permission from Haines DE (ed): Fundamental Neuroscience for Basic and Clinical Applications, 3rd ed. St. Louis, MO: Elsevier; 2006.)
شکل 12-11 یک طرح مداری نشان دهنده ضایعات ایجاد شده در حیوانات آزمایشگاهی برای تکرار نقصهای decerebrate و decorticate که در انسان مشاهده میشود. برشهای دو طرفه با خطوط بریده A، B، C و D نشان داده میشوند. دسربراسیون در سطح میانی کولیکولار (A)، دکورتیکاسیون منقاری به کولیکولاس فوقانی، ریشههای پشتی برش برای یک اندام (B) و آسیب به لوب قدامیمخچه (C) است. هدف شناسایی لایههای آناتومیک مسئول سفتی/استقرار دسربره یا دکورتیکهای بود که در انسانها با ضایعاتی دیده میشود که یا جلوی مغز را از ساقه مغز جدا میکنند یا منقاری را از ساقه مغز و نخاعی جدا میکنند. (تکثیر شده با مجوز Haines DE (ed): Fundamental Neuroscience for Basic and Clinical Applications, 3rd ed. St. Louis, MO: Elsevier; 2006.)

FIGURE 12-12 Decerebrate and decorticate postures. A) Damage to lower midbrain and upper pons causes decerebrate posturing in which lower extremities are extended with toes pointed inward and upper extremities extended with fingers flexed and forearms pronate. Neck and head are extended. B) Damage to upper midbrain may cause decorticate posturing in which upper limbs are flexed, lower limbs are extended with toes pointed slightly inward, and head is extended. (Modified with permission from Kandel ER, Schwartz JH, Jessell TM (eds): Principles of Neural Science, 4th ed. New York, NY: McGraw- Hill; 2000.)
شکل 12-12 وضعیتها را دسربره و تزئین کنید. الف) آسیب به مغز میانی تحتانی و ران فوقانی باعث ایجاد وضعیت نامناسب میشود که در آن اندام تحتانی با انگشتان پا به سمت داخل کشیده شده و اندام فوقانی با انگشتان خم شده و ساعد به صورت پرونیت کشیده میشوند. گردن و سر کشیده شده اند. ب) آسیب به مغز میانی فوقانی ممکن است باعث ایجاد حالت دکورتیک شود که در آن اندامهای فوقانی خم شده اند، اندام تحتانی با انگشتان پا کمیبه سمت داخل کشیده میشوند و سر کشیده میشود. (اصلاح شده با اجازه Kandel ER، Schwartz JH، Jessell TM (eds): Principles of Neural Science، نسخه چهارم نیویورک، نیویورک: مک گراو-هیل؛ 2000.)
CLINICAL BOX 12-6
Uncal Herniation
Space-occupying lesions from large tumors, hemorrhages, strokes, or abscesses in the cerebral hemisphere can drive the uncus of the temporal lobe over the edge of the cerebellar tentorium, compressing the ipsilateral cranial nerve III (uncal herniation). Before the herniation these patients experience a decreased level of consciousness, lethargy, poorly reactive pupils, deviation of the eye to a “down and out” position, hyperactive reflexes, and a bilateral Babinski sign (due to compression of the ipsilateral corticospinal tract). After the brain herniates, the patients are decerebrate and comatose, have fixed and dilated pupils, and eye movements are absent. Once damage extends to the midbrain, a Cheyne-Stokes respiratory pattern develops, characterized by a pattern of waxing-and-waning depth of respiration with interposed periods of apnea. Eventually, medullary function is lost, breathing ceases, and recovery is unlikely. Hemispheric masses closer to the midline compress the thalamic reticular formation and can cause coma before eye findings develop (central herniation). As the mass enlarges, midbrain function is affected, the pupils dilate, and a decerebrate posture ensues. With progressive herniation, pontine vestibular and then medullary respiratory functions are lost.
جعبه بالینی 12.6
فتق آنکال
ضایعات فضایی ناشی از تومورهای بزرگ، خونریزیها، سکتهها، یا آبسهها در نیمکره مغزی میتوانند بر روی لبه تنتوریوم مخچه رانده شوند و عصب جمجمه ای همان طرف را فشرده کنند (فتق ناقص). قبل از فتق، این بیماران کاهش سطح هوشیاری، بی حالی، مردمکهای با واکنش ضعیف، انحراف چشم به حالت “پایین و بیرون”، رفلکسهای بیش فعال و علامت بابینسکی دو طرفه (به دلیل فشرده شدن دستگاه قشر نخاعی همان طرف) را تجربه میکنند. پس از فتق مغزی، بیماران دچار دسربره و کما، مردمکهای ثابت و گشاد شده و حرکات چشم وجود ندارد. هنگامیکه آسیب به مغز میانی گسترش مییابد، یک الگوی تنفسی Cheyne-Stokes ایجاد میشود که با یک الگوی اپیلاسیون و کاهش عمق تنفس همراه با دورههای بینابینی آپنه مشخص میشود. در نهایت، عملکرد مدولاری از بین میرود، تنفس متوقف میشود و بهبودی بعید است. تودههای نیمکره نزدیک به خط وسط تشکیل شبکه تالاموس را فشرده میکنند و میتوانند قبل از ایجاد یافتههای چشمیباعث کما شوند (فتق مرکزی). با بزرگ شدن توده، عملکرد مغز میانی تحت تأثیر قرار میگیرد، مردمکها گشاد میشوند و وضعیت نامناسبی ایجاد میشود. با فتق پیشرونده، عملکرد تنفسی دهلیزی پونتین و سپس مدولاری از بین میرود.
DECORTICATION
Injury to the cerebral cortex (decortication; dashed line labeled D in Figure 12– 11) produces decorticate rigidity that is characterized by flexion of the upper extremities at the elbow and extensor hyperactivity in the lower extremities (Figure 12-12B). The flexion is explained by rubrospinal excitation of flexor muscles in the upper extremities; the hyperextension of lower extremities is due to the same changes that occur after midcollicular decerebration.
تزئینات
آسیب به قشر مغز (تقسیم، خط بریده شده با برچسب D در شکل 12-11) باعث سفتی دکورتکس میشود که با خم شدن اندامهای فوقانی در آرنج و بیش فعالی اکستانسور در اندام تحتانی مشخص میشود (شکل 12-12B). خم شدن با تحریک روبروسنخاعی عضلات فلکسور در اندام فوقانی توضیح داده میشود. اکستنشن اندام تحتانی به دلیل همان تغییراتی است که پس از دسربراسیون میانی کولیکولار رخ میدهد.
Decorticate rigidity is seen on the hemiplegic side after hemorrhages or thromboses in the internal capsule. The small arteries in the internal capsule are especially prone to rupture or thrombotic obstruction, so this type of decorticate rigidity is fairly common. Sixty percent of intracerebral hemorrhages occur in the internal capsule, 10% in the cerebral cortex, 10% in the pons, 10% in the thalamus, and 10% in the cerebellum.
سفتی تزئینی در سمت همیپلژیک پس از خونریزی یا ترومبوز در کپسول داخلی دیده میشود. شریانهای کوچک در کپسول داخلی به ویژه مستعد پارگی یا انسداد ترومبوتیک هستند، بنابراین این نوع سفتی تزئینی نسبتاً رایج است. شصت درصد خونریزیهای داخل مغزی در کپسول داخلی، 10 درصد در قشر مغز، 10 درصد در پونز، 10 درصد در تالاموس و 10 درصد در مخچه رخ میدهد.
BASAL GANGLIA
ORGANIZATION OF THE BASAL GANGLIA
The term basal ganglia (or basal nuclei) is applied to five interactive structures on each side of the brain (Figure 12-13). These are the caudate nucleus, putamen, and globus pallidus (three large nuclear masses underlying the cortical mantle), the subthalamic nucleus, and substantia nigra. The caudate nucleus and putamen collectively form the striatum; the putamen and globus pallidus collectively form the lenticular nucleus.
بازال گانگلیا
سازمان گانگلیا پایه
اصطلاح عقدههای پایه (یا هستههای پایه) به پنج ساختار تعاملی در هر طرف مغز به کار میرود (شکل 12-13). اینها هسته دمی، پوتامن و گلوبوس پالیدوس (سه توده هسته ای بزرگ زیر گوشته قشری)، هسته زیر تالاموس و ماده سیاه هستند. هسته دمیو پوتامن مجموعاً مخطط را تشکیل میدهند. پوتامن و گلوبوس پالیدوس در مجموع هسته عدسی را تشکیل میدهند.

FIGURE 12-13 The basal ganglia. The basal ganglia are composed of the caudate nucleus, putamen, and globus pallidus and the functionally related subthalamic nucleus and substantia nigra. The frontal (coronal) section shows the location of the basal ganglia in relation to surrounding structures.
شکل 12-13 عقدههای قاعده ای. عقدههای قاعده ای از هسته دمی، پوتامن، و گلوبوس پالیدوس و هسته زیر تالاموس و ماده سیاه از نظر عملکردی تشکیل شده اند. بخش فرونتال (کرونال) موقعیت گانگلیونهای پایه را در رابطه با ساختارهای اطراف نشان میدهد.
The globus pallidus is divided into external and internal segments (GPe and GPi); both regions contain inhibitory GABAergic neurons. The substantia nigra is divided into pars compacta that uses dopamine as a neurotransmitter and pars reticulata that uses GABA as a neurotransmitter. About 95% of striatal neurons are medium spiny neurons that use GABA as a neurotransmitter. The remaining striatal neurons are aspiny interneurons that differ in terms of size and neurotransmitters: large (acetylcholine), medium (somatostatin), and small (GABA).
گلوبوس پالیدوس به بخشهای خارجی و داخلی (GPe و GPi) تقسیم میشود. هر دو ناحیه حاوی نورونهای GABAergic مهاری هستند. ماده سیاه به pars compacta که از دوپامین به عنوان یک انتقال دهنده عصبی استفاده میکند و pars reticulata که از GABA به عنوان یک انتقال دهنده عصبی استفاده میکند، تقسیم میشود. حدود 95 درصد از نورونهای جسم مخطط نورونهای خاردار متوسطی هستند که از GABA به عنوان یک انتقال دهنده عصبی استفاده میکنند. نورونهای جسم مخطط باقیمانده، نورونهای آسپینی هستند که از نظر اندازه و انتقال دهندههای عصبی متفاوت هستند: بزرگ (استیل کولین)، متوسط (سوماتوستاتین) و کوچک (GABA).
Figure 12-14 shows the major connections to and from and within the basal ganglia along with the neurotransmitters within these pathways. There are two main inputs to the basal ganglia; they are both excitatory (glutamate), and they both terminate in the striatum. They are from a wide region of the cerebral cortex (corticostriate pathway) and from intralaminar nuclei of the thalamus (thalamostriatal pathway). The two major outputs of the basal ganglia are from GPi and substantia nigra pars reticulata. Both are inhibitory (GABAergic) and both project to the thalamus. From the thalamus, there is an excitatory (glutamatergic) projection to the prefrontal and premotor cortex. This completes the cortical-basal ganglia-thalamic-cortical loop.
شکل 12-14 اتصالات عمده به و از و درون عقدههای پایه همراه با انتقال دهندههای عصبی در این مسیرها را نشان میدهد. دو ورودی اصلی به عقدههای پایه وجود دارد. آنها هر دو تحریک کننده هستند (گلوتامات)، و هر دو به مخطط ختم میشوند. آنها از ناحیه وسیعی از قشر مغز (مسیر کورتیکوستریات) و از هستههای داخل لایه ای تالاموس (مسیر تالاموستریاتال) هستند. دو خروجی اصلی عقدههای پایه از GPi و ماده سیاه پارس رتیکولاتا هستند. هر دو مهار کننده هستند (GABAergic) و هر دو به تالاموس برمیخیزند. از تالاموس، یک برآمدگی تحریکی (گلوتاماترژیک) به قشر جلوی پیشانی و پیش حرکتی وجود دارد. این حلقه گانگلیون قشری-پایه-تالاموس-قشری کامل میشود.
The connections within the basal ganglia include a dopaminergic nigrostriatal projection from the substantia nigra pars compacta to the striatum and a GABAergic projection from the striatum to substantia nigra pars reticulata. There is an inhibitory projection from the striatum to both GPe and GPi. The subthalamic nucleus receives an inhibitory input from GPe, and in turn the subthalamic nucleus has an excitatory (glutamatergic) projection to both GPe and GPi.
اتصالات درون عقدههای پایه شامل یک برآمدگی دوپامینرژیک سیاه استریاتال از جسم سیاه پارس فشرده به جسم مخطط و یک برآمدگی GABAergic از جسم مخطط به ماده سیاه پارس مشبک است. یک طرح مهاری از جسم مخطط به GPe و GPi وجود دارد. هسته ساب تالاموس یک ورودی بازدارنده از GPe دریافت میکند و به نوبه خود هسته زیر تالاموس دارای یک برآمدگی تحریکی (گلوتاماترژیک) به GPe و GPi است.
FIGURE 12–14 Diagrammatic representation of the principal connections of the basal ganglia. Solid lines indicate excitatory pathways, dashed lines indicate inhibitory pathways. The transmitters are indicated in the pathways, where they are known. DA, dopamine; Glu, glutamate. Acetylcholine is the transmitter produced by interneurons in the striatum. ES, external segment; IS, internal segment; PPN, pedunculopontine nuclei; SNPC, substantia nigra, pars compacta; SNPR, substantia nigra, pars reticulata. The subthalamic nucleus also projects to the pars compacta of the substantia nigra; this pathway has been omitted for clarity.
شکل 12-14 نمایش نموداری اتصالات اصلی عقدههای پایه. خطوط توپر مسیرهای تحریکی را نشان میدهند، خطوط نقطه چین مسیرهای بازدارنده را نشان میدهند. فرستندهها در مسیرهایی که شناخته شده اند نشان داده شده اند. DA، دوپامین؛ گلو، گلوتامات. استیل کولین فرستنده ای است که توسط نورونهای داخلی مخطط تولید میشود. ES، بخش خارجی؛ IS، بخش داخلی؛ PPN، هسته پدانکولوپونتین. SNPC، substantia nigra، pars compacta; SNPR، ماده سیاه، pars reticulata. هسته ساب تالاموس نیز به سمت pars compacta جسم سیاه پیش میرود. این مسیر برای وضوح حذف شده است.
FUNCTION
Neurons in the basal ganglia, like those in the lateral portions of the cerebellar hemispheres, discharge before movements begin. The basal ganglia are involved in the planning and programming of movement or, more broadly, in the processes by which an abstract thought is converted into voluntary action (Figure 12-6). They influence the motor cortex via the thalamus, and the corticospinal pathways provide the final common pathway to motor neurons. In addition, GPi projects to nuclei in the brainstem, and from there to motor neurons in the brainstem and spinal cord. The basal ganglia, particularly the caudate nuclei, also play a role in some cognitive processes. Possibly because of the interconnections of this nucleus with the frontal portions of the neocortex, lesions of the caudate nuclei disrupt performance on tests involving object reversal and delayed alternation. In addition, lesions of the head of the left but not the right caudate nucleus and nearby white matter in humans are associated with a dysarthric form of aphasia that resembles Wernicke aphasia (see Chapter 15).
تابع
نورونها در عقدههای قاعدهای، مانند نورونهایی که در قسمتهای جانبی نیمکرههای مخچه هستند، قبل از شروع حرکات ترشح میشوند. عقدههای پایه در برنامهریزی و برنامهریزی حرکت یا بهطور گستردهتر، در فرآیندهایی که توسط آن یک فکر انتزاعی به عمل ارادی تبدیل میشود، درگیر هستند (شکل 12-6). آنها از طریق تالاموس بر قشر حرکتی تأثیر میگذارند و مسیرهای قشر نخاعی آخرین مسیر مشترک را برای نورونهای حرکتی فراهم میکنند. علاوه بر این، GPi به هستههای ساقه مغز و از آنجا به نورونهای حرکتی در ساقه مغز و نخاع میپردازد. عقدههای قاعده ای، به ویژه هستههای دمی، نیز در برخی از فرآیندهای شناختی نقش دارند. احتمالاً به دلیل اتصالات متقابل این هسته با بخشهای پیشانی نئوکورتکس، ضایعات هستههای دمیعملکرد در آزمایشهای مربوط به معکوس شدن جسم و تناوب تاخیری را مختل میکنند. بعلاوه، ضایعات سر هسته دمیسمت چپ، اما نه هسته دمیراست و ماده سفید مجاور در انسان، با شکل دیزارتریک آفازی که شبیه آفازی Wernicke است، همراه است (به فصل 15 مراجعه کنید).
DISEASES OF THE BASAL GANGLIA IN HUMANS
Three distinct biochemical pathways in the basal ganglia normally operate in a balanced manner: (1) the nigrostriatal dopaminergic system, (2) the intrastriatal cholinergic system, and (3) the GABAergic system that projects from the striatum to the globus pallidus and substantia nigra. When one or more of these pathways become dysfunctional, characteristic motor abnormalities occur. Diseases of the basal ganglia lead to two general types of disorders: hyperkinetic and hypokinetic. The hyperkinetic conditions are those in which movement is excessive and abnormal, including chorea, athetosis, and ballism. Hypokinetic abnormalities include akinesia and bradykinesia.
بیماریهای گانگلیای بازال در انسان
سه مسیر بیوشیمیایی متمایز در عقدههای قاعده ای به طور معمول به صورت متعادل عمل میکنند: (1) سیستم دوپامینرژیک سیاه و سفید، (2) سیستم کولینرژیک داخل استریاتال و (3) سیستم GABAergic که از جسم مخطط به سمت گلوبوس پالیدوس و ماده سیاه پیش میرود. هنگامیکه یک یا چند مورد از این مسیرها ناکارآمد میشود، ناهنجاریهای حرکتی مشخصه رخ میدهد. بیماریهای عقدههای قاعده ای منجر به دو نوع اختلال کلی میشود:هایپرکینتیک و هیپوکینتیک. شرایط هیپرکینتیک شرایطی است که در آن حرکت بیش از حد و غیر طبیعی است، از جمله کوریا، آتتوز و بالستیک. ناهنجاریهای هیپوکینتیک شامل آکینزی و برادی کینزی است.
Chorea is characterized by rapid, involuntary “dancing” movements. Athetosis is characterized by continuous, slow writhing movements. Choreiform and athetotic movements have been likened to the start of voluntary movements occurring in an involuntary, disorganized way. In ballism, involuntary flailing, intense, and violent movements occur. Akinesia is difficulty in initiating movement and decreased spontaneous movement. Bradykinesia is slowness of movement.
Chorea با حرکات سریع و غیر ارادی “رقص” مشخص میشود. آتتوز با حرکات پیوسته و آهسته انقباض مشخص میشود. حرکات کوریفورم و آتتوتیک به شروع حرکات ارادی تشبیه شده اند که به صورت غیرارادی و بی نظم اتفاق میافتد. در بالیسم، لرزیدن غیرارادی، حرکات شدید و خشن رخ میدهد. آکینزی مشکل در شروع حرکت و کاهش حرکت خود به خودی است. برادیکینزی کندی حرکت است.
In addition to Parkinson disease (described below), there are several other disorders that involve a malfunction within the basal ganglia. A few of these are described in Clinical Box 12-7. Huntington disease is one of an increasing number of human genetic diseases affecting the nervous system that are characterized by trinucleotide repeat expansion. Most of these involve cytosine-adenine-guanine (CAG) repeats (Table 12-1), but one involves cytosine-guanine-guanine (CGG) repeats and another involves CTG repeats (T refers to thymine). Increased numbers of a 12-nucleotide repeat are also associated with a rare form of epilepsy.
علاوه بر بیماری پارکینسون (که در زیر توضیح داده شده است)، چندین اختلال دیگر وجود دارد که شامل اختلال در عملکرد گانگلیون پایه است. تعدادی از این موارد در جعبه بالینی 12-7 توضیح داده شده است. بیماریهانتینگتون یکی از تعداد فزاینده بیماریهای ژنتیکی انسان است که سیستم عصبی را تحت تأثیر قرار میدهد که با گسترش تکراری سه نوکلئوتیدی مشخص میشود. بیشتر این موارد شامل تکرارهای سیتوزین-آدنین-گوانین (CAG) است (جدول 12-1)، اما یکی شامل تکرارهای سیتوزین-گوانین-گوانین (CGG) و دیگری شامل تکرارهای CTG (T به تیمین اشاره دارد). افزایش تعداد تکرارهای 12 نوکلئوتیدی نیز با شکل نادری از صرع همراه است.
TABLE 12-1 Examples of trinucleotide repeat diseases.
جدول 12-1 نمونههایی از بیماریهای تکراری تری نوکلئوتیدی.
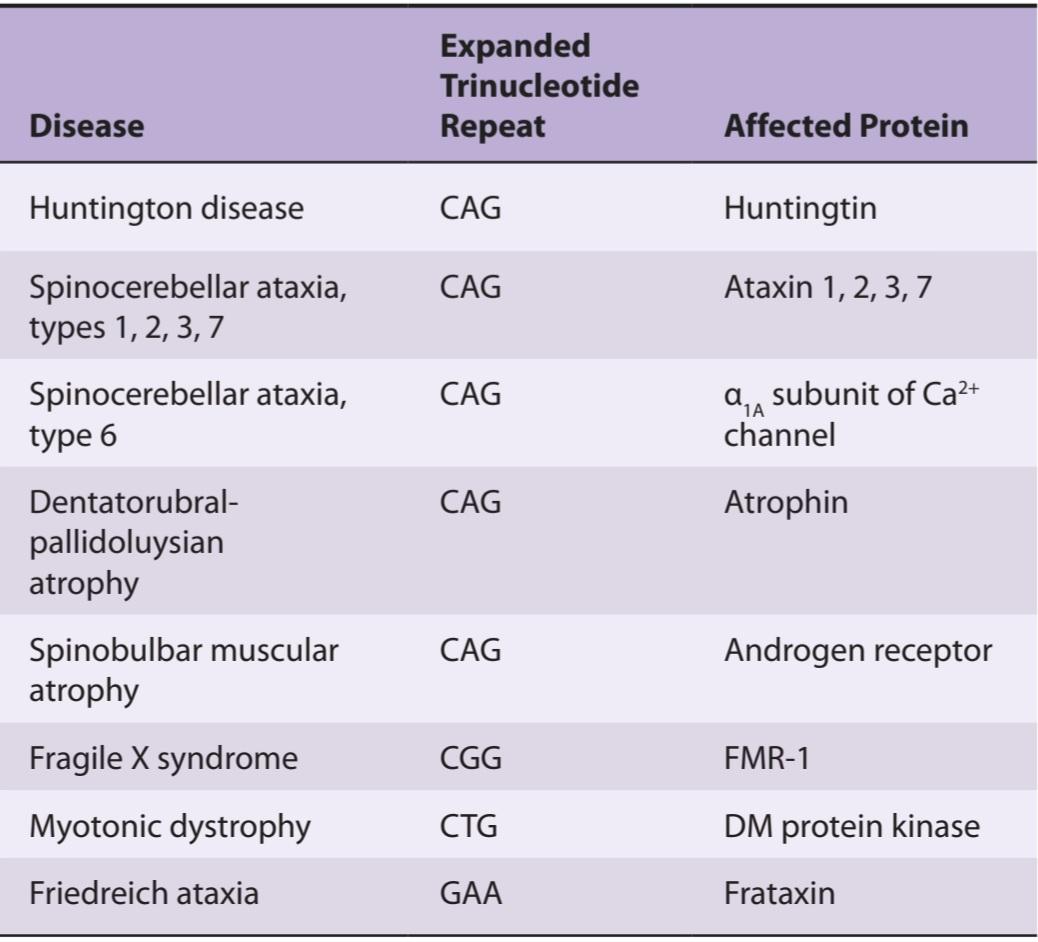
CLINICAL BOX 12. 7
Basal Ganglia Diseases
The initial detectable damage in Huntington disease is to medium spiny neurons in the striatum. The loss of this GABAergic pathway to the GPe releases inhibition, permitting the hyperkinetic features of the disease to develop. An early sign is a jerky trajectory of the hand when reaching to touch a spot, especially toward the end of the reach. Later, hyperkinetic choreiform movements appear and gradually increase until they incapacitate the patient. Speech becomes slurred and then incomprehensible, and a progressive dementia is followed by death, usually within 10-15 years after the onset of symptoms. Huntington disease affects 5 out of 100,000 people worldwide. It is inherited as an autosomal dominant disorder, and its onset is usually between the ages of 30 and 50. The abnormal gene responsible for the disease is located near the end of the short arm of chromosome 4. It normally contains 11-34 CAG repeats, each coding for glutamine. In patients with Huntington disease, this number is increased to 42-86 or more copies; and the greater the number of repeats, the earlier the age of onset and the more rapid the progression of the disease. The gene codes for huntingtin, a protein that plays a role in axonal trafficking, regulation of gene transcription, and cell survival. Poorly soluble protein aggregates, which are toxic, form in cell nuclei and elsewhere. However, the correlation between aggregates and symptoms is less than perfect. The loss of the function of huntingtin that occurs is proportional to the size of the CAG insert. In animal models of the disease, intrastriatal grafting of fetal striatal tissue improves cognitive performance. In addition, tissue caspase-1 activity is increased in the brains of humans with the disease. Moreover, in mice in which the gene for this apoptosis-regulating enzyme has been knocked out, progression of the disease is slowed.
جعبه بالینی 12.7
بیماریهای گانگلیون بازال
آسیب قابل تشخیص اولیه در بیماریهانتینگتون به نورونهای خاردار متوسط در جسم مخطط است. از دست دادن این مسیر GABAergic به GPe باعث بازداری میشود و به ویژگیهای هیپرکینتیک بیماری اجازه میدهد تا ایجاد شود. علامت اولیه یک مسیر پرشتاب دست هنگام رسیدن به لمس نقطه ای است، به خصوص در انتهای دست. بعداً حرکات کوریفرمهایپرکینتیک ظاهر میشود و به تدریج افزایش مییابد تا زمانی که بیمار را ناتوان میکند. گفتار نامفهوم و نامفهوم میشود و به دنبال آن زوال عقل پیشرونده، معمولاً در عرض 10-15 سال پس از شروع علائم، مرگ را به دنبال دارد. بیماریهانتینگتون از هر 100000 نفر در سراسر جهان 5 نفر را مبتلا میکند. این بیماری به عنوان یک اختلال اتوزومال غالب به ارث میرسد و شروع آن معمولا بین 30 تا 50 سالگی است. ژن غیر طبیعی مسئول این بیماری در نزدیکی انتهای بازوی کوتاه کروموزوم 4 قرار دارد. به طور معمول شامل 11-34 تکرار CAG است که هر کدام برای گلوتامین کد میکنند. در بیماران مبتلا به بیماریهانتینگتون، این تعداد به 42-86 نسخه یا بیشتر افزایش مییابد. و هر چه تعداد دفعات تکرار بیشتر باشد سن شروع زودتر و پیشرفت بیماری سریعتر میشود. کدهای ژنهانتینگتین، پروتئینی که در قاچاق آکسون، تنظیم رونویسی ژن و بقای سلولی نقش دارد. تودههای پروتئینی با محلول ضعیف، که سمیهستند، در هستههای سلولی و جاهای دیگر تشکیل میشوند. با این حال، همبستگی بین تجمعات و علائم کمتر از حد کامل است. از دست دادن عملکردهانتینگتین که رخ میدهد متناسب با اندازه درج CAG است. در مدلهای حیوانی این بیماری، پیوند داخل مخطط بافت مخطط جنین عملکرد شناختی را بهبود میبخشد. علاوه بر این، فعالیت کاسپاز-1 بافتی در مغز انسانهای مبتلا به این بیماری افزایش مییابد. علاوه بر این، در موشهایی که ژن این آنزیم تنظیم کننده آپوپتوز در آنها از بین رفته است، پیشرفت بیماری کند میشود.
Wilson disease (or hepatolenticular degeneration) is a rare disorder of copper metabolism that has an onset between 6 and 25 years of age, affecting about four times as many females as males. Wilson disease affects about 30,000 people worldwide. It is a genetic autosomal recessive disorder due to a mutation on the long arm of chromosome 13q. It affects the copper- transporting ATPase gene (ATP7B) in the liver, leading to an accumulation of copper in the liver and resultant progressive liver damage. About 1% of the population carries a single abnormal copy of this gene but does not develop any symptoms. The disease may develop in a child who inherits the gene from both parents. In affected individuals, copper accumulates in the periphery of the cornea in the eye accounting for the characteristic yellow Kayser-Fleischer rings. The dominant neuronal pathology is degeneration of the putamen, a part of the lenticular nucleus. Motor disturbances include “wing-beating” tremor or asterixis, dysarthria, unsteady gait, and rigidity. Tardive dyskinesia is a disease that involves the basal ganglia, but it is caused by medical treatment of another disorder with neuroleptic drugs such as phenothiazines or haloperidol. Therefore, tardive dyskinesia is iatrogenic in origin. Long-term use of these drugs may produce biochemical abnormalities in the striatum. The motor disturbances include either temporary or permanent uncontrolled involuntary movements of the face and tongue and cogwheel rigidity. The neuroleptic drugs act via blockade of dopaminergic transmission. Prolonged drug use leads to hypersensitivity of D3 dopaminergic receptors and an imbalance in nigrostriatal influences on motor control.
بیماری ویلسون (یا دژنراسیون کبدی) یک اختلال نادر در متابولیسم مس است که بین 6 تا 25 سالگی شروع میشود و تقریباً چهار برابر مردان در زنان تأثیر میگذارد. بیماری ویلسون حدود 30000 نفر را در سراسر جهان تحت تاثیر قرار میدهد. این یک اختلال اتوزومیمغلوب ژنتیکی است که به دلیل جهش در بازوی بلند کروموزوم 13q به وجود میآید. این ژن ATPase انتقال دهنده مس (ATP7B) در کبد را تحت تاثیر قرار میدهد و منجر به تجمع مس در کبد و در نتیجه آسیب تدریجی کبد میشود. حدود 1٪ از جمعیت یک نسخه غیر طبیعی از این ژن را دارند اما هیچ علامتی ندارند. این بیماری ممکن است در کودکی ایجاد شود که این ژن را از هر دو والدین به ارث برده است. در افراد مبتلا، مس در حاشیه قرنیه چشم جمع میشود و حلقههای زرد رنگ مشخصه Kayser-Fleischer را تشکیل میدهد. آسیب شناسی عصبی غالب، دژنراسیون پوتامن، بخشی از هسته عدسی است. اختلالات حرکتی شامل لرزش یا آستریکسیس “بال زدن”، دیس آرتری، راه رفتن ناپایدار و سفتی است. دیسکینزی دیررس یک بیماری است که گانگلیونهای پایه را درگیر میکند، اما به دلیل درمان پزشکی اختلال دیگری با داروهای نورولپتیک مانند فنوتیازینها یاهالوپریدول ایجاد میشود. بنابراین، دیسکینزی دیررس منشأ ایتروژنیک دارد. استفاده طولانی مدت از این داروها ممکن است باعث ایجاد ناهنجاریهای بیوشیمیایی در جسم مخطط شود. اختلالات حرکتی شامل حرکات غیرارادی موقت یا دائمیغیرارادی صورت و زبان و سفتی چرخ دنده است. داروهای اعصاب از طریق مسدود کردن انتقال دوپامینرژیک عمل میکنند. استفاده طولانی مدت از دارو منجر به حساسیت بیش از حد گیرندههای دوپامینرژیک D3 و عدم تعادل در تأثیرات سیاه و سفید بر روی کنترل حرکتی میشود.
THERAPEUTIC HIGHLIGHTS
Treatment for Huntington disease is directed at treating the symptoms and maintaining quality of life because there is no cure. In general, drugs used to treat the symptoms of this disease have side effects such as fatigue, nausea, and restlessness. In August 2008, the US Food and Drug Administration approved the use of tetrabenazine to reduce choreiform movements that characterize the disease. This drug binds reversibly to vesicular monoamine transporters (VMATs) and thus inhibits the uptake of monoamines into synaptic vesicles. It also acts as a dopamine receptor antagonist. Tetrabenazine is the first drug to receive approval for individuals with Huntington disease. It is also used to treat other hyperkinetic movement disorders such as tardive dyskinesia. Chelating agents (eg, penicillamine and trienthine) are used to reduce the copper in the body in individuals with Wilson disease. Tardive dyskinesia has proven to be difficult to treat. Treatment in patients with psychiatric disorders is often directed at prescribing a neuroleptic with less likelihood of causing the disorder. Clozapine is an atypical neuroleptic drug that has been an effective substitute for traditional neuroleptic drugs but with less risk for development of tardive dyskinesia.
نکات برجسته درمانی
درمان بیماریهانتینگتون در جهت درمان علائم و حفظ کیفیت زندگی است زیرا هیچ درمانی وجود ندارد. به طور کلی داروهایی که برای درمان علائم این بیماری استفاده میشوند عوارضی مانند خستگی، حالت تهوع و بی قراری دارند. در آگوست 2008، سازمان غذا و داروی ایالات متحده استفاده از تترابنازین را برای کاهش حرکات کوریفرم که مشخصه بیماری است، تایید کرد. این دارو به طور برگشت پذیر به انتقال دهندههای مونوآمین تاولی (VMATs) متصل میشود و بنابراین جذب مونوآمینها را به وزیکولهای سیناپسی مهار میکند. همچنین به عنوان یک آنتاگونیست گیرنده دوپامین عمل میکند. تترابنازین اولین دارویی است که برای افراد مبتلا به بیماریهانتینگتون تایید شده است. همچنین برای درمان سایر اختلالات حرکتی هیپرکینتیک مانند دیسکینزی دیررس استفاده میشود. عوامل کیلات (مانند پنیسیلامین و تریانتین) برای کاهش مس در بدن در افراد مبتلا به بیماری ویلسون استفاده میشود. ثابت شده است که درمان دیررس دیسکینزی دشوار است. درمان در بیماران مبتلا به اختلالات روانپزشکی اغلب با تجویز داروی نورولپتیک با احتمال کمتری برای ایجاد اختلال انجام میشود. کلوزاپین یک داروی نورولپتیک غیر معمول است که جایگزین موثری برای داروهای اعصاب سنتی بوده اما خطر کمتری برای ایجاد دیسکینزی دیررس دارد.
PARKINSON DISEASE
Parkinson disease has both hypokinetic and hyperkinetic features. It was originally described in 1817 by James Parkinson and is named for him. Parkinson disease is the first disease identified as being due to a deficiency in a specific neurotransmitter (Clinical Box 12-8). It results from the degeneration of dopaminergic neurons in the substantia nigra pars compacta. The fibers to the putamen (part of the striatum) are most severely affected.
بیماری پارکینسون
بیماری پارکینسون هر دو ویژگی هیپوکینتیک و هیپرکینتیک دارد. در ابتدا در سال 1817 توسط جیمز پارکینسون توصیف شد و به نام او نامگذاری شد. بیماری پارکینسون اولین بیماری است که به دلیل کمبود یک انتقال دهنده عصبی خاص شناسایی شده است (باکس بالینی 12-8). این ناشی از تخریب نورونهای دوپامینرژیک در جسم سیاه پارس فشرده است. فیبرهای پوتامن (بخشی از جسم مخطط) به شدت تحت تأثیر قرار میگیرند.
The hypokinetic features of Parkinson disease are akinesia and bradykinesia, and the hyperkinetic features are cogwheel rigidity and tremor at rest. The absence of motor activity and the difficulty in initiating voluntary movements are striking. There is a decrease in the normal, unconscious movements such as swinging of the arms during walking, the panorama of facial expressions related to the emotional content of thought and speech, and the multiple “fidgety” actions and gestures that occur in all of us. The rigidity is different from spasticity because motor neuron discharge increases to both the agonist and antagonist muscles. Passive motion of an extremity meets with a plastic, deadfeeling resistance that has been likened to bending a lead pipe and is therefore called lead pipe rigidity. Sometimes a series of “catches” takes place during passive motion (cogwheel rigidity), but the sudden loss of resistance seen in a spastic extremity is absent. The tremor, which is present at rest and disappears with activity, is due to regular, alternating 8-Hz contractions of antagonistic muscles.
ویژگیهای هیپوکینتیک بیماری پارکینسون آکینزی و برادیکینزی است و ویژگیهای هیپوکینتیک سفتی چرخ دنده و لرزش در حالت استراحت است. فقدان فعالیت حرکتی و مشکل در شروع حرکات ارادی قابل توجه است. کاهش در حرکات عادی و ناخودآگاه مانند تاب خوردن بازوها در حین راه رفتن، پانورامای حالات چهره مرتبط با محتوای عاطفی فکر و گفتار، و اعمال و حرکات متعدد «بیتابی» که در همه ما رخ میدهد، کاهش مییابد. سفتی با اسپاستیسیته متفاوت است، زیرا ترشحات نورون حرکتی به عضلات آگونیست و آنتاگونیست افزایش مییابد. حرکت غیرفعال یک اندام با یک مقاومت پلاستیکی و مرده روبرو میشود که به خمش لوله سربی تشبیه شده است و بنابراین صلبیت لوله سربی نامیده میشود. گاهی اوقات یک سری “گرفتن” در حین حرکت غیرفعال (سفتی چرخ دنده) اتفاق میافتد، اما کاهش ناگهانی مقاومت که در اندام اسپاستیک دیده میشود وجود ندارد. لرزش که در حالت استراحت وجود دارد و با فعالیت ناپدید میشود، به دلیل انقباضات منظم و متناوب 8 هرتز عضلات متضاد است.
Figure 12-15 shows the current view of the pathogenesis of the movement disorders in Parkinson disease. In healthy individuals, basal ganglia output is inhibitory via GABAergic nerve fibers. The dopaminergic neurons that project from the substantia nigra to the putamen normally have two effects. They stimulate the D1 dopamine receptors, which inhibit GPi via direct GABAergic receptors; and they inhibit D2 receptors, which also inhibit the GPe. In addition, the inhibition reduces the excitatory discharge from the subthalamic nucleus to the GPi. This balance between inhibition and excitation somehow maintains normal motor function. In Parkinson disease, the dopaminergic input to the putamen is lost. This results in decreased inhibition and increased excitation from the subthalamic nucleus to the GPi. The overall increase in inhibitory output to the thalamus and brainstem disorganizes movement.
شکل 12-15 نمای فعلی پاتوژنز اختلالات حرکتی در بیماری پارکینسون را نشان میدهد. در افراد سالم، برون ده گانگلیون پایه از طریق رشتههای عصبی GABAergic مهار میشود. نورونهای دوپامینرژیک که از جسم سیاه به پوتامن بیرون میروند معمولاً دو اثر دارند. آنها گیرندههای دوپامین D1 را تحریک میکنند که GPi را از طریق گیرندههای مستقیم GABAergic مهار میکند. و گیرندههای D2 را مهار میکنند که GPe را نیز مهار میکند. علاوه بر این، مهار باعث کاهش ترشحات تحریکی از هسته ساب تالاموس به GPi میشود. این تعادل بین مهار و تحریک به نحوی عملکرد حرکتی طبیعی را حفظ میکند. در بیماری پارکینسون، ورودی دوپامینرژیک به پوتامن از بین میرود. این منجر به کاهش مهار و افزایش تحریک از هسته ساب تالاموس به GPi میشود. افزایش کلی برون ده مهاری به تالاموس و ساقه مغز حرکت را به هم ریخته است.
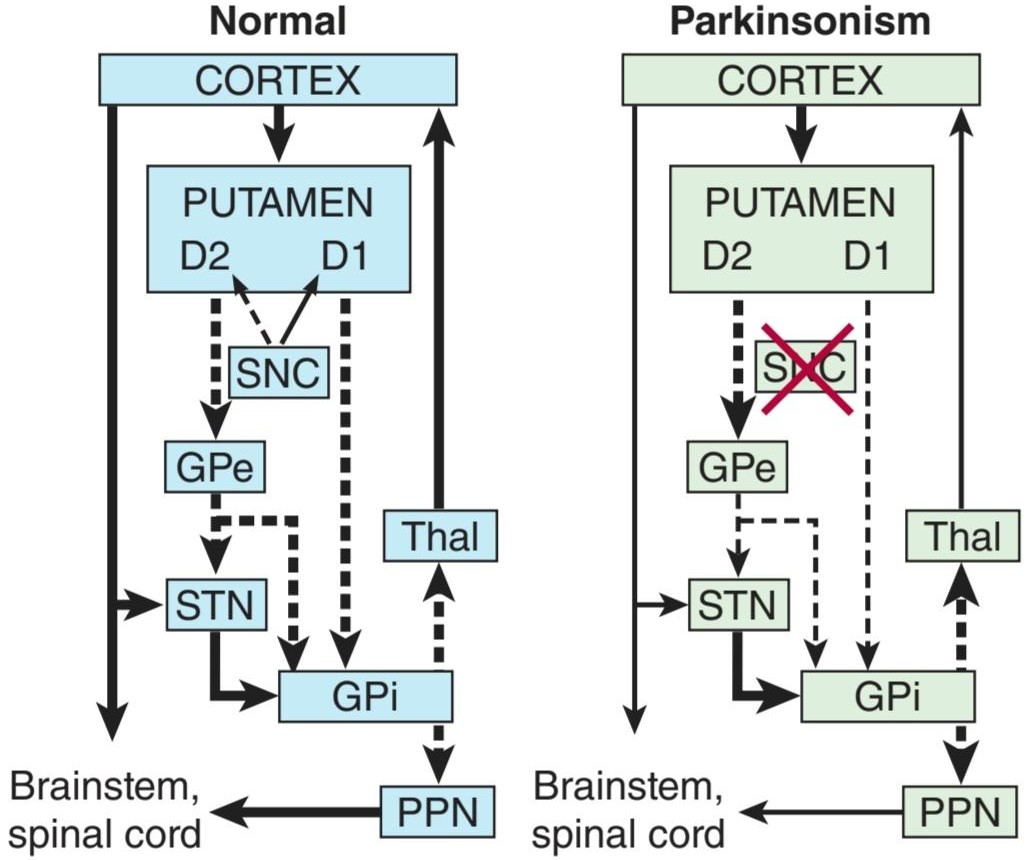
FIGURE 12-15 Probable basal ganglia-thalamocortical circuitry in Parkinson disease. Solid arrows indicate excitatory outputs and dashed arrows inhibitory outputs. The strength of each output is indicated by the width of the arrow. GPe, external segment of the globus pallidus; GPi, internal segment of the globus pallidus; PPN, pedunculopontine nuclei; SNC, pars compacta of the substantia nigra; STN, subthalamic nucleus; Thal, thalamus. See text for details. (Modified with permission from Grafton SC, DeLong M: Tracing the brain circuitry with functional imaging, Nat Med 1997 Jun; 3(6):602–603.)
شکل 12-15 مدار عقدههای قاعده ای-تالاموکورتیکال احتمالی در بیماری پارکینسون. فلشهای توپر خروجیهای تحریکی و فلشهای چین دار خروجیهای بازدارنده را نشان میدهند. قدرت هر خروجی با عرض فلش نشان داده میشود. GPe، بخش خارجی گلوبوس پالیدوس. GPi، بخش داخلی گلوبوس پالیدوس. PPN، هسته پدانکولوپونتین. SNC، pars compacta از جسم سیاه; STN، هسته ساب تالاموس؛ ثال، تالاموس. برای جزئیات به متن مراجعه کنید (اصلاح شده با مجوز Grafton SC، DeLong M: Tracing the مغز مدار با تصویربرداری عملکردی، Nat Med 1997 ژوئن؛ 3(6):602-603.)
Familial cases of Parkinson disease occur, but these are uncommon. The genes for at least five proteins can be mutated. These proteins appear to be involved in ubiquitination. Two of the proteins, a-synuclein and parkin, interact and are found in Lewy bodies. The Lewy bodies are inclusion bodies in neurons that occur in all forms of Parkinson disease.
موارد خانوادگی بیماری پارکینسون رخ میدهد، اما این موارد غیر معمول هستند. ژنهای حداقل پنج پروتئین میتوانند جهش یافته باشند. به نظر میرسد این پروتئینها در ubiquitination نقش دارند. دو تا از پروتئینها، a-synuclein و پارکین، برهم کنش دارند و در اجسام Lewy یافت میشوند. اجسام لوی اجسام گنجانده شده در نورونها هستند که در همه اشکال بیماری پارکینسون رخ میدهند.
An important consideration in Parkinson disease is the balance between the excitatory discharge of cholinergic interneurons and the inhibitory dopaminergic input in the striatum. Some improvement is produced by decreasing the cholinergic influence with anticholinergic drugs. More dramatic improvement is produced by administration of L-dopa. Unlike dopamine, this dopamine precursor crosses the blood-brain barrier and helps repair the dopamine deficiency. However, the degeneration of these neurons continues and in 5-7 years the beneficial effects of L-dopa disappear.
یک نکته مهم در بیماری پارکینسون، تعادل بین ترشحات تحریکی نورونهای کولینرژیک و ورودی دوپامینرژیک مهاری در جسم مخطط است. مقداری بهبود با کاهش اثر کولینرژیک با داروهای آنتی کولینرژیک ایجاد میشود. بهبود چشمگیرتری با تجویز L-dopa ایجاد میشود. برخلاف دوپامین، این پیش ساز دوپامین از سد خونی مغزی عبور میکند و به ترمیم کمبود دوپامین کمک میکند. با این حال، انحطاط این نورونها ادامه مییابد و در 5-7 سال اثرات مفید L-dopa از بین میرود.
CLINICAL BOX 12.8
Parkinson Disease
There are 7-10 million people worldwide in whom Parkinson disease has been diagnosed. The disease is 1.5 times more prevalent in men than in women. Each year in the United States, nearly 60,000 new cases are diagnosed. Parkinsonism occurs in sporadic idiopathic form in many middle- aged and elderly individuals and is one of the most common neurodegenerative diseases. It is estimated to occur in 1-2% of individuals over age 65. Dopaminergic neurons and dopamine receptors are steadily lost with age in the basal ganglia in healthy individuals, and an acceleration of these losses apparently precipitates parkinsonism. Symptoms appear when 60-80% of the nigrostriatal dopaminergic neurons degenerate.
جعبه بالینی12.8
بیماری پارکینسون
7 تا 10 میلیون نفر در سراسر جهان وجود دارند که بیماری پارکینسون در آنها تشخیص داده شده است. این بیماری در مردان 1.5 برابر بیشتر از زنان است. هر سال در ایالات متحده، نزدیک به 60000 مورد جدید تشخیص داده میشود. پارکینسونیسم به شکل ایدیوپاتیک پراکنده در بسیاری از افراد میانسال و مسن رخ میدهد و یکی از شایع ترین بیماریهای نورودژنراتیو است. تخمین زده میشود که در 1-2٪ از افراد بالای 65 سال رخ دهد. نورونهای دوپامینرژیک و گیرندههای دوپامین به طور پیوسته با افزایش سن در گانگلیونهای پایه در افراد سالم از بین میروند، و تسریع این تلفات ظاهراً پارکینسونیسم را تشدید میکند. علائم زمانی ظاهر میشوند که 60 تا 80 درصد از نورونهای دوپامینرژیک سیاه پوست دچار انحطاط میشوند.
Parkinsonism is also seen as a complication of treatment with the phenothiazine group of antipsychotic drugs and other drugs that block dopaminergic D2 receptors. It can be produced in rapid and dramatic form by injection of 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine (MPTP). This effect was discovered by chance when a drug dealer in northern California supplied some of his clients with a homemade preparation of synthetic heroin that contained MPTP. MPTP is a prodrug that is metabolized in astrocytes by the enzyme monoamine oxidase (MOA-B) to produce a potent oxidant, 1- methyl-4-phenylpyridinium (MPP+). MPP+ is taken up by the dopamine transporter into dopaminergic neurons in the substantia nigra, which it destroys without affecting other dopaminergic neurons to any appreciable degree.
پارکینسونیسم همچنین به عنوان یک عارضه درمان با گروه فنوتیازین از داروهای ضد روان پریشی و سایر داروهایی که گیرندههای دوپامینرژیک D2 را مسدود میکنند، دیده میشود. میتوان آن را به شکل سریع و چشمگیر با تزریق 1-متیل-4-فنیل-1،2،5،6-تتراهیدروپیریدین (MPTP) تولید کرد. این اثر به طور تصادفی زمانی کشف شد که یک فروشنده مواد مخدر در شمال کالیفرنیا به برخی از مشتریان خود یک آماده سازی خانگی از هروئین مصنوعی که حاوی MPTP بود، عرضه کرد. MPTP یک پیش دارویی است که در آستروسیتها توسط آنزیم مونوآمین اکسیداز (MOA-B) متابولیزه میشود تا یک اکسید کننده قوی به نام 1- متیل-4- فنیل پیریدینیوم (+MPP) تولید کند. +MPP توسط ناقل دوپامین به نورونهای دوپامینرژیک در جسم سیاه جذب میشود، که بدون تأثیر بر سایر نورونهای دوپامینرژیک به میزان قابلتوجهی از بین میرود.
THERAPEUTIC HIGHLIGHTS
There is no cure for Parkinson disease, and drug therapies are designed to treat the symptoms. Sinemet, a combination of levodopa (L-dopa) and carbidopa, is the most commonly used drug for the treatment of Parkinson disease. The addition of carbidopa to L-dopa increases its effectiveness and prevents the conversion of L-dopa to dopamine in the periphery and thus reduces some of the adverse side effects of L-dopa (nausea, vomiting, and cardiac rhythm disturbances). Dopamine agonists, including apomorphine, bromocriptine, pramipexole, and ropinirole, have also proven effective in some patients with Parkinson disease. Taken in combination with L-dopa, cathechol-O methyltransferase (COMT) inhibitors (eg, entacapone) are another class of drugs used to treat this disease. They act by blocking the breakdown of L-dopa, allowing more of it to reach the brain to increase the level of dopamine. MAO-B inhibitors (eg, selegiline) also prevent the breakdown of dopamine. They can be given soon after diagnosis and delay the need for L-dopa.
نکات برجسته درمانی
هیچ درمانی برای بیماری پارکینسون وجود ندارد و درمانهای دارویی برای درمان علائم طراحی شده اند. Sinemet، ترکیبی از لوودوپا (L-dopa) و کاربیدوپا، رایج ترین داروی مورد استفاده برای درمان بیماری پارکینسون است. افزودن کاربیدوپا به ال-دوپا اثربخشی آن را افزایش میدهد و از تبدیل ال-دوپا به دوپامین در محیط اطراف جلوگیری میکند و در نتیجه برخی از عوارض جانبی نامطلوب ال-دوپا (تهوع، استفراغ و اختلالات ریتم قلبی) را کاهش میدهد. آگونیستهای دوپامین، از جمله آپومورفین، بروموکریپتین، پرامیپکسول و روپینیرول نیز در برخی از بیماران مبتلا به پارکینسون مؤثر بودهاند. در ترکیب با L-dopa، مهارکنندههای کاتکول-O متیل ترانسفراز (COMT) (مثلاً انتاکاپون) دسته دیگری از داروهای مورد استفاده برای درمان این بیماری هستند. آنها با جلوگیری از تجزیه ال-دوپا عمل میکنند و اجازه میدهند مقدار بیشتری از آن به مغز برسد تا سطح دوپامین را افزایش دهد. مهارکنندههای MAO-B (به عنوان مثال، سلژیلین) همچنین از تجزیه دوپامین جلوگیری میکنند. آنها میتوانند بلافاصله پس از تشخیص داده شوند و نیاز به L-dopa را به تاخیر بیندازند.
The US Food and Drug Administration has approved the use of deep brain stimulation (DBS) for the treatment of Parkinson disease. DBS reduces the amount of L-dopa patients need and thus reduces its adverse side effects (eg, involuntary movements called dyskinesias). DBS has been associated with reduction of tremors, slowness of movements, and gait problems in some patients. Surgical treatments in general are reserved for those who have exhausted drug therapies or who have not responded favorably to them. Lesions in GPi (pallidotomy) or in the subthalamic nucleus (thalamotomy) have been performed to help restore the output balance of the basal ganglia toward normal (see Figure 12-15). Another surgical approach is to implant dopamine-secreting tissue in or near the basal ganglia. Transplants of the patient’s own adrenal medullary tissue or carotid body works for a while, apparently by functioning as a sort of dopamine minipump, but long-term results have been disappointing. Results with transplantation of fetal striatal tissue have been better, and transplanted cells have been shown to not only survive but also make appropriate connections in the host’s basal ganglia. However, some patients with transplants develop dyskinesias due to excessive levels of dopamine.
سازمان غذا و داروی آمریکا استفاده از تحریک عمقی مغز (DBS) را برای درمان بیماری پارکینسون تایید کرده است. DBS میزان نیاز بیماران L-dopa را کاهش میدهد و در نتیجه عوارض جانبی نامطلوب آن را کاهش میدهد (مانند حرکات غیرارادی به نام دیسکینزی). DBS با کاهش لرزش، کندی حرکات و مشکلات راه رفتن در برخی بیماران همراه است. درمانهای جراحی به طور کلی برای کسانی است که درمانهای دارویی را تمام کرده اند یا به آنها پاسخ مثبت نداده اند. ضایعات در GPi (پالیدوتومی) یا در هسته ساب تالاموس (تالاموتومی) برای کمک به بازگرداندن تعادل خروجی عقدههای پایه به سمت نرمال انجام شده است (شکل 12-15 را ببینید). یکی دیگر از روشهای جراحی کاشت بافت ترشح کننده دوپامین در داخل یا نزدیک عقدههای پایه است. پیوند بافت مدولاری آدرنال خود بیمار یا بدن کاروتید برای مدتی کار میکند، ظاهراً به عنوان نوعی پمپ دوپامین عمل میکند، اما نتایج دراز مدت ناامید کننده بوده است. نتایج پیوند بافت مخطط جنین بهتر بوده است و سلولهای پیوندی نشان داده شده است که نه تنها زنده میمانند بلکه اتصالات مناسبی را در عقدههای پایه میزبان ایجاد میکنند. با این حال، برخی از بیماران پیوندی به دلیل سطوح بیش از حد دوپامین دچار دیسکینزی میشوند.
CEREBELLUM
The cerebellum sits astride the main sensory and motor systems in the brainstem (Figure 12-16). The medial vermis and lateral cerebellar hemispheres are more extensively folded and fissured than the cerebral cortex. The cerebellum weighs only 10% as much as the cerebral cortex, but its surface area is about 75% of that of the cerebral cortex. Anatomically, the cerebellum is divided into three parts by two transverse fissures. The posterolateral fissure separates the medial nodulus and the lateral flocculus on either side from the rest of the cerebellum, and the primary fissure divides the remainder into an anterior and a posterior lobe. Lesser fissures divide the vermis into smaller sections, so that it contains 10 primary lobules numbered I-X from superior to inferior.
مخچه
مخچه بر روی سیستمهای حسی و حرکتی اصلی در ساقه مغز قرار دارد (شکل 12-16). ورم میانی و نیمکرههای مخچه جانبی نسبت به قشر مخ چین خوردهتر و شکاف دارند. وزن مخچه تنها 10 درصد به اندازه قشر مخ است، اما مساحت سطح آن حدود 75 درصد از سطح قشر مخ است. از نظر تشریحی، مخچه توسط دو شکاف عرضی به سه قسمت تقسیم میشود. شقاق خلفی جانبی گره داخلی و لخته جانبی دو طرف را از بقیه مخچه جدا میکند و شقاق اولیه باقی مانده را به یک لوب قدامیو یک لوب خلفی تقسیم میکند. شکافهای کوچکتر ورمیس را به بخشهای کوچکتری تقسیم میکنند، به طوری که شامل 10 لوبول اولیه با شماره I-X از بالا به پایین است.

FIGURE 12-16 A midsagittal section through the cerebellum. The medial vermis and lateral cerebellar hemispheres have many narrow, ridge-like folds called folia. Although not shown, the cerebellum is connected to the brainstem by three pairs of peduncles (superior, middle, and inferior). (Reproduced with permission from Waxman SG: Clinical Neuroanatomy, 26th ed. New York, NY: McGraw-Hill; 2010.)
شکل 12-16 یک بخش میانی ساژیتال از مخچه. ورم میانی و نیمکرههای مخچه جانبی دارای چینهای باریک و برآمدگی زیادی هستند که به آنها فولیا میگویند. اگرچه مخچه نشان داده نشده است، اما با سه جفت دمگل (بالا، میانی و تحتانی) به ساقه مغز متصل است. (بازتولید شده با اجازه Waxman SG: Clinical Neuroanatomy, 26th. New York, NY: McGraw-Hill; 2010.)
The cerebellum is connected to the brainstem by three pairs of peduncles that are located above and around the fourth ventricle. The superior cerebellar peduncle includes fibers from deep cerebellar nuclei that project to the brainstem, red nucleus, and thalamus. The middle cerebellar peduncle contains only afferent fibers from the contralateral pontine nuclei, and the inferior cerebellar peduncle a mixture of afferent fibers from the brainstem and spinal cord and efferent fibers to the vestibular nuclei.
مخچه توسط سه جفت دمگل که در بالا و اطراف بطن چهارم قرار دارند به ساقه مغز متصل میشود. ساقه مخچه فوقانی شامل الیافی از هستههای عمیق مخچه است که به ساقه مغز، هسته قرمز و تالاموس میرسد. دمگل مخچه میانی فقط شامل الیاف آوران از هستههای پونتین طرف مقابل است و دمگل مخچه تحتانی مخلوطی از الیاف آوران از ساقه مغز و نخاع و الیاف وابران به هستههای دهلیزی است.
The cerebellum has an external cerebellar cortex separated by white matter from the deep cerebellar nuclei. The middle and inferior cerebellar peduncles carry afferent fibers into the cerebellum where they are called mossy and climbing fibers. These fibers emit collaterals to the deep nuclei and pass to the cortex. There are four deep nuclei: the dentate, the globose, the emboliform, and the fastigial nuclei. The globose and the emboliform nuclei are sometimes lumped together as the interpositus nucleus.
مخچه دارای یک قشر مخچه خارجی است که توسط ماده سفید از هستههای عمیق مخچه جدا شده است. دمگلهای مخچه میانی و تحتانی الیاف آوران را به مخچه حمل میکنند که به آنها الیاف خزه ای و بالا رونده میگویند. این الیاف وثیقههایی را به هستههای عمیق ساطع میکنند و به قشر مغز منتقل میشوند. چهار هسته عمیق وجود دارد: هسته دندانه دار، کروی، آمبولی فرم و هسته فاستیژال. هستههای کروی و آمبولیفرم گاهی بهعنوان هسته بینپوزیتوس به هم متصل میشوند.
ORGANIZATION OF THE CEREBELLUM
The cerebellar cortex has three layers: an external molecular layer, a Purkinje cell layer that is only one cell thick, and an internal granular layer. There are five types of neurons in the cortex: Purkinje, granule, basket, stellate, and Golgi cells (Figure 12-17). The Purkinje cells are among the largest neurons in the CNS. They have extensive dendritic arbors that extend throughout the molecular layer. Their axons, which are the only output from the cerebellar cortex, project to the deep cerebellar nuclei, especially the dentate nucleus, where they form inhibitory synapses. They also make inhibitory connections with neurons in the vestibular nuclei.
سازمان مخچه
قشر مخچه دارای سه لایه است: یک لایه مولکولی خارجی، یک لایه سلولی پورکنژ که تنها یک سلول ضخامت دارد و یک لایه دانه ای داخلی. پنج نوع نورون در قشر مغز وجود دارد: سلولهای پورکنژ، گرانول، سبد، ستاره ای و گلژی (شکل 12-17). سلولهای پورکنژ یکی از بزرگترین نورونها در CNS هستند. آنها دارای درختچههای دندریتی گسترده ای هستند که در سراسر لایه مولکولی گسترش یافته اند. آکسونهای آنها که تنها خروجی از قشر مخچه هستند، به هستههای عمیق مخچه، به ویژه هسته دندانه دار، که در آنجا سیناپسهای بازدارنده را تشکیل میدهند، پیش میروند. آنها همچنین با نورونهای هسته دهلیزی ارتباط مهاری برقرار میکنند.
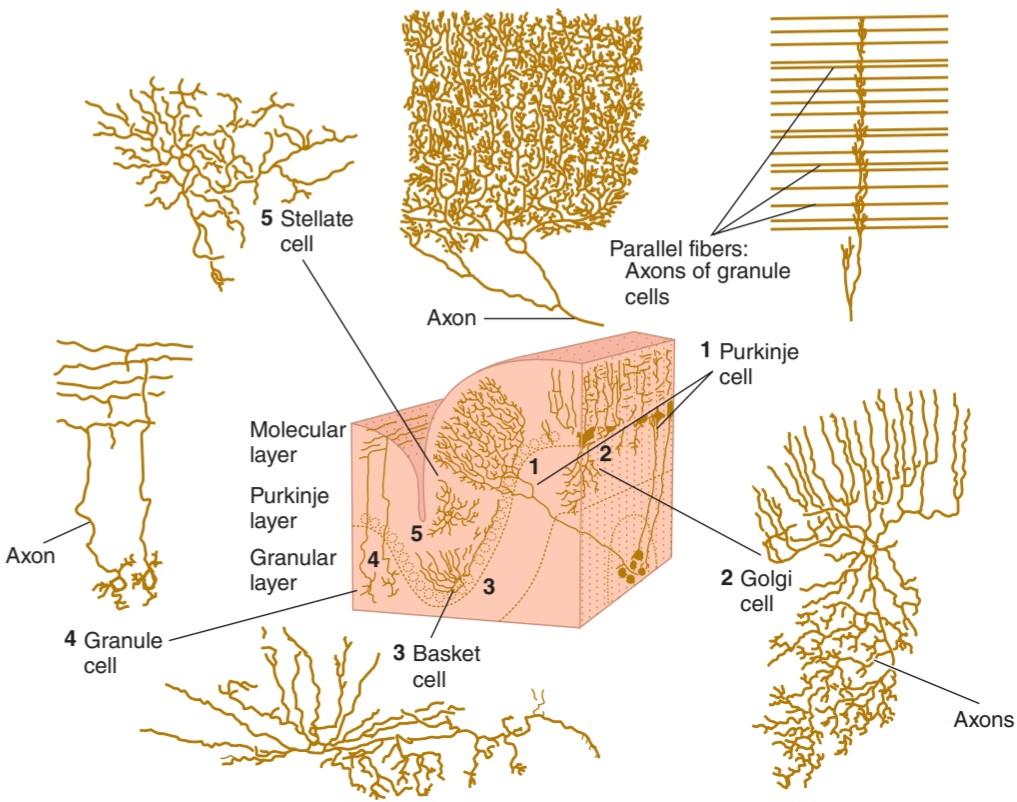
FIGURE 12-17 Location and structure of five neuronal types in the cerebellar cortex. Drawings are based on Golgi-stained preparations. Purkinje cells (1) have processes aligned in one plane; their axons are the only output from the cerebellum. Axons of granule cells (4) traverse and make connections with Purkinje cell processes in molecular layer. Golgi (2), basket (3), and stellate (5) cells have characteristic positions, shapes, branching patterns, and synaptic connections. (For 1 and 2, Reproduced with permission of Ramon y Cajal S: Histologie du Systeme Nerveux II., C.S.I.C. Madrid; For 3-5, Reproduced with permission of Palay SL, Chan-Palay V: Cerebellar Cortex. Berlin: Springer- Verlag, 1975.)
شکل 12-17 محل و ساختار پنج نوع عصبی در قشر مخچه. نقاشیها بر اساس آماده سازیهای رنگ آمیزی گلژی است. سلولهای پورکنژ (1) دارای فرآیندهایی هستند که در یک صفحه قرار دارند. آکسون آنها تنها خروجی از مخچه است. آکسونهای سلولهای گرانول (4) از فرآیندهای سلول پورکنژ در لایه مولکولی عبور کرده و با آنها ارتباط برقرار میکنند. سلولهای گلژی (2)، سبد (3) و ستاره ای (5) دارای موقعیتها، شکلها، الگوهای انشعاب و اتصالات سیناپسی هستند. (برای 1 و 2، تکثیر شده با اجازه Ramon y Cajal S: Histologie du Systeme Nerveux II., C.S.I.C. Madrid؛ برای 3-5، تکثیر شده با اجازه Palay SL، Chan-Palay V: Cerebellar Cortex. برلین: Springer- Verlag، 195).
The cerebellar granule cells, whose cell bodies are in the granular layer, receive excitatory input from the mossy fibers and innervate the Purkinje cells (Figure 12-18). Each sends an axon to the molecular layer, where the axon bifurcates to form a T. The branches of the T are straight and run long distances; thus, they are called parallel fibers. The dendritic trees of the Purkinje cells are markedly flattened and oriented at right angles to the parallel fibers. The parallel fibers form excitatory synapses on the dendrites of many Purkinje cells, and the parallel fibers and Purkinje dendritic trees form a grid of remarkably regular proportions.
سلولهای گرانول مخچه، که بدنه سلولی آنها در لایه دانه ای قرار دارد، ورودی تحریکی را از الیاف خزه دریافت میکند و سلولهای پورکنژ را عصب دهی میکند (شکل 12-18). هر کدام یک آکسون را به لایه مولکولی میفرستند، جایی که آکسون دوشاخه میشود و T را تشکیل میدهد. بنابراین، آنها را الیاف موازی مینامند. درختان دندریتیک سلولهای پورکنژ به طور قابل توجهی مسطح و در زوایای قائم به رشتههای موازی قرار دارند. الیاف موازی سیناپسهای تحریکی را روی دندریتهای بسیاری از سلولهای پورکنژ تشکیل میدهند و الیاف موازی و درختان دندریتی پورکنژ شبکهای با نسبتهای منظمیرا تشکیل میدهند.
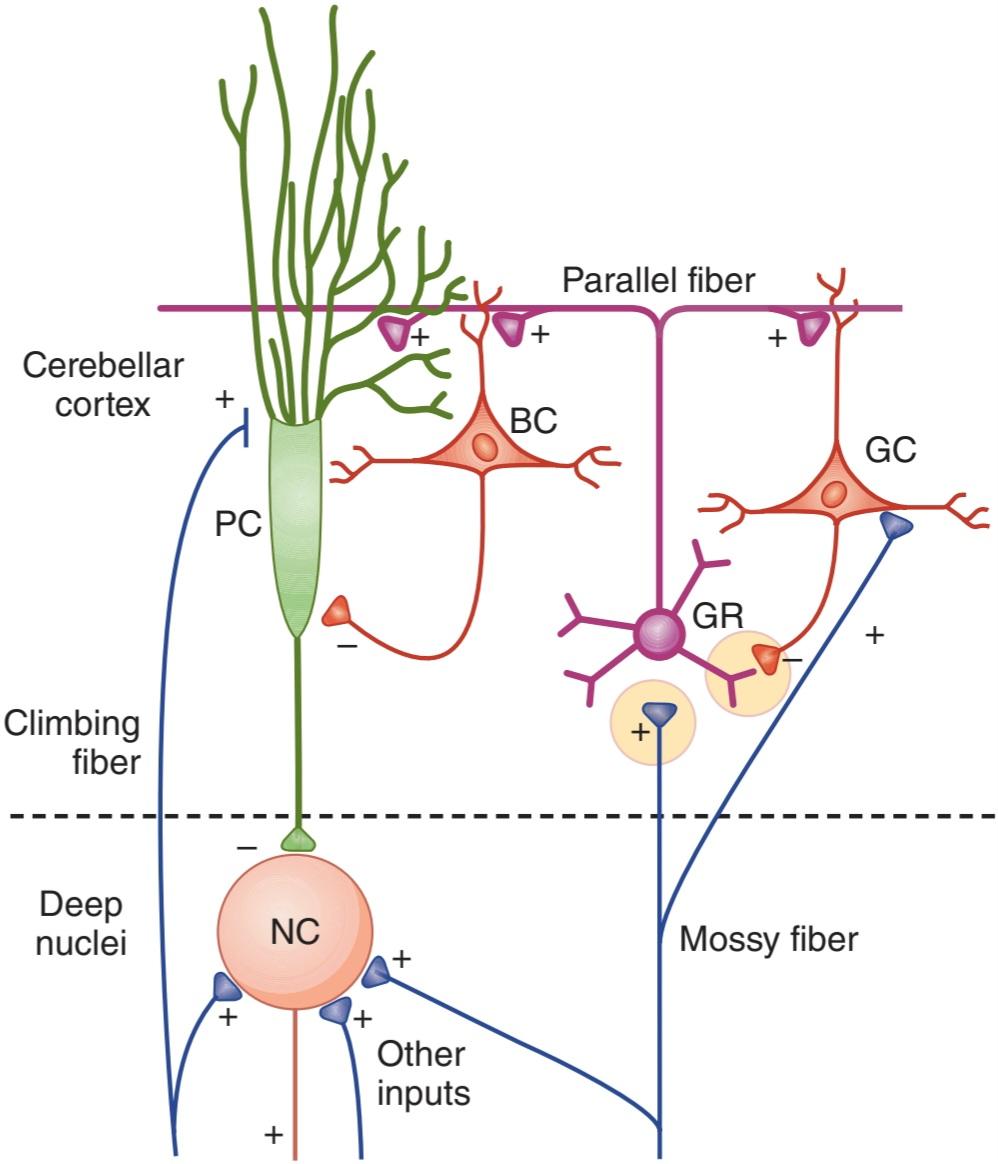
FIGURE 12-18 Diagram of neural connections in the cerebellum. Plus (+) and minus (-) signs indicate whether endings are excitatory or inhibitory. BC, basket cell; GC, Golgi cell; GR, granule cell; NC, cell in deep nucleus; PC, Purkinje cell. Note that PCs and BCs are inhibitory. The connections of the stellate cells, which are not shown, are similar to those of the basket cells, except that they end for the most part on Purkinje cell dendrites.
شکل 12-18 نمودار اتصالات عصبی در مخچه. علائم به علاوه (+) و منهای (-) نشان میدهد که انتهای آن تحریک کننده یا بازدارنده است. قبل از میلاد، سلول سبد; GC، سلول گلژی. GR، سلول گرانول. NC، سلول در هسته عمیق. کامپیوتر، سلول پورکنژ. توجه داشته باشید که رایانههای شخصی و BCها بازدارنده هستند. اتصالات سلولهای ستارهای که نشان داده نشدهاند، شبیه به سلولهای سبد هستند، با این تفاوت که بیشتر آنها به دندریتهای سلول پورکنژ ختم میشوند.
The other three types of neurons in the cerebellar cortex are inhibitory interneurons. Basket cells (Figure 12-17) are located in the molecular layer. They receive excitatory input from the parallel fibers and each projects to many Purkinje cells (Figure 12–18). Their axons form a basket around the cell body and axon hillock of each Purkinje cell they innervate. Stellate cells are similar to the basket cells but are located in the more superficial molecular layer. Golgi cells are located in the granular layer. Their dendrites project into the molecular layer and receive excitatory input from the parallel fibers (Figure 12-18). Their cell bodies receive excitatory input via collaterals from the incoming mossy fibers. Their axons project to the dendrites of the granule cells where they form an inhibitory synapse.
سه نوع نورون دیگر در قشر مخچه، نورونهای بازدارنده هستند. سلولهای سبد (شکل 12-17) در لایه مولکولی قرار دارند. آنها ورودی تحریکی را از فیبرهای موازی دریافت میکنند و هر کدام به سلولهای پورکنژ زیادی میپردازند (شکل 12-18). آکسون آنها سبدی را در اطراف بدن سلولی و تپه آکسون هر سلول پورکنژی که آنها را عصب میکنند تشکیل میدهد. سلولهای ستاره ای شبیه به سلولهای سبد هستند اما در لایه مولکولی سطحی تر قرار دارند. سلولهای گلژی در لایه دانه ای قرار دارند. دندریتهای آنها به لایه مولکولی فرو میروند و ورودی تحریکی را از الیاف موازی دریافت میکنند (شکل 12-18). بدن سلولی آنها ورودی تحریکی را از طریق وثیقههای فیبرهای خزه ای دریافتی دریافت میکند. آکسونهای آنها به دندریتهای سلولهای گرانول پیش میروند و در آنجا یک سیناپس مهاری تشکیل میدهند.
As already mentioned, the two excitatory main inputs to the cerebellar cortex are climbing fibers and mossy fibers (Figure 12-18). The climbing fibers come from a single source, the inferior olivary nuclei. Each projects to the primary dendrites of a Purkinje cell, around which it entwines like a climbing plant. Proprioceptive input to the inferior olivary nuclei comes from all over the body.
همانطور که قبلاً ذکر شد، دو ورودی اصلی تحریک کننده به قشر مخچه فیبرهای بالا رونده و الیاف خزه ای هستند (شکل 12-18). الیاف کوهنوردی از یک منبع منفرد میآیند، هستههای زیتونی تحتانی. هر کدام به سمت دندریتهای اولیه سلول پورکنژ حرکت میکند که مانند یک گیاه بالارونده در اطراف آن در هم میپیچد. ورودی حس عمقی به هستههای زیتون تحتانی از سراسر بدن میآید.
On the other hand, the mossy fibers provide direct proprioceptive input from all parts of the body plus input from the cerebral cortex via the pontine nuclei to the cerebellar cortex. They end on the dendrites of granule cells in complex synaptic groupings called glomeruli. The glomeruli also contain the inhibitory endings of the Golgi cells mentioned above.
از سوی دیگر، الیاف خزه ای ورودی مستقیم حس عمقی را از تمام قسمتهای بدن به اضافه ورودی از قشر مخ از طریق هستههای پونتین به قشر مخچه ارائه میدهند. آنها به دندریت سلولهای گرانول در گروههای سیناپسی پیچیده ای به نام گلومرول ختم میشوند. گلومرولها همچنین حاوی انتهای بازدارنده سلولهای گلژی هستند که در بالا ذکر شد.
The fundamental circuits of the cerebellar cortex are relatively simple (Figure 12-18). Climbing fiber inputs exert a strong excitatory effect on single Purkinje cells, whereas mossy fiber inputs exert a weak excitatory effect on many Purkinje cells via the granule cells. The basket and stellate cells are also excited by granule cells via their parallel fibers; and the basket and stellate cells, in turn, inhibit the Purkinje cells (feed-forward inhibition). Golgi cells are excited by the mossy fiber collaterals and parallel fibers, and they inhibit transmission from mossy fibers to granule cells. The neurotransmitter released by the stellate, basket, Golgi, and Purkinje cells is GABA, whereas the granule cells release glutamate. GABA acts via GABAA receptors, but the combinations of subunits in these receptors vary from one cell type to the next. The granule cell is unique in that it appears to be the only type of neuron in the CNS that has a GABAд receptor containing the a6 subunit.
مدارهای اساسی قشر مخچه نسبتا ساده هستند (شکل 12-18). ورودیهای فیبر صعودی یک اثر تحریکی قوی روی سلولهای منفرد پورکنژ اعمال میکنند، در حالی که ورودیهای فیبر خزه ای از طریق سلولهای گرانول روی بسیاری از سلولهای پورکنژ اثر تحریکی ضعیفی دارند. سلولهای سبد و ستارهای نیز توسط سلولهای گرانول از طریق الیاف موازی خود تحریک میشوند. و سلولهای سبد و ستارهای به نوبه خود سلولهای پورکنژ را مهار میکنند (ممانعت از پیشخور). سلولهای گلژی توسط وثیقههای فیبر خزه ای و الیاف موازی برانگیخته میشوند و از انتقال از الیاف خزه ای به سلولهای گرانول جلوگیری میکنند. انتقال دهنده عصبی آزاد شده توسط سلولهای ستاره ای، سبد، گلژی و پورکنژ GABA است، در حالی که سلولهای گرانول گلوتامات آزاد میکنند. گابا از طریق گیرندههای GABAA عمل میکند، اما ترکیب زیر واحدها در این گیرندهها از یک نوع سلول به نوع دیگر متفاوت است. سلول گرانول از این نظر منحصر به فرد است که به نظر میرسد تنها نوع نورون در CNS است که دارای گیرنده GABAд حاوی زیرواحد a6 است.
The output of the Purkinje cells is in turn inhibitory to the deep cerebellar nuclei. As noted above, these nuclei also receive excitatory inputs via collaterals from the mossy and climbing fibers. It is interesting, in view of their inhibitory Purkinje cell input, that the output of the deep cerebellar nuclei to the brainstem and thalamus is always excitatory. Thus, almost all the cerebellar circuitry seems to be concerned solely with modulating or timing the excitatory output of the deep cerebellar nuclei to the brainstem and thalamus. The primary afferent systems that converge to form the mossy fiber or climbing fiber input to the cerebellum are summarized in Table 12-2.
خروجی سلولهای پورکنژ به نوبه خود مهار کننده هستههای عمیق مخچه است. همانطور که در بالا ذکر شد، این هستهها همچنین ورودیهای تحریکی را از طریق وثیقه از الیاف خزه ای و صعودی دریافت میکنند. با توجه به ورودی مهاری سلول پورکنژ جالب است که خروجی هستههای عمیق مخچه به ساقه مغز و تالاموس همیشه تحریک کننده است. بنابراین، تقریباً تمام مدارهای مخچه صرفاً به تعدیل یا زمانبندی خروجی تحریکی هستههای عمیق مخچه به ساقه مغز و تالاموس مربوط میشوند. سیستمهای آوران اولیه که برای تشکیل فیبر خزهای یا ورودی فیبر بالارونده به مخچه همگرا میشوند در جدول 12-2 خلاصه شدهاند.
TABLE 12-2 Function of principal afferent systems to the cerebellum.
جدول 12-2 عملکرد سیستمهای آوران اصلی به مخچه.
aThe olivocerebellar pathway projects to the cerebellar cortex via climbing fibers; the rest of the listed paths project via mossy fibers. Several other pathways transmit impulses from nuclei in the brainstem to the cerebellar cortex and to the deep nuclei, including a serotonergic input from the raphe nuclei to the granular and molecular layers and a noradrenergic input from the locus coeruleus to all three layers.
aمسیر olivocerebellar از طریق الیاف بالا رونده به قشر مخچه پیش میرود؛ بقیه مسیرهای ذکر شده از طریق الیاف خزه ای پیش میروند. چندین مسیر دیگر تکانهها را از هستههای ساقه مغز به قشر مخچه و به هستههای عمیق منتقل میکنند، از جمله یک لایه سروتونرژیک و یک هسته مولکولار به لایه مغزی به سمت لایه مغزی. ورودی نورآدرنرژیک از لوکوس سرولئوس به هر سه لایه.
FUNCTIONAL DIVISIONS
From a functional point of view, the cerebellum is divided into three parts (Figure 12-19). The nodulus in the vermis and the flanking flocculus in the hemisphere on each side form the vestibulocerebellum (or flocculonodular lobe). This lobe is phylogenetically the oldest part of the cerebellum and has vestibular connections concerned with equilibrium and eye movements. The rest of the vermis and the adjacent medial portions of the hemispheres form the spinocerebellum, the region that receives proprioceptive input from the body as well as a copy of the “motor plan” from the motor cortex. By comparing plan with performance, it smooths and coordinates movements that are ongoing. The vermis projects to the brainstem area concerned with control of axial and proximal limb muscles (medial brainstem pathways, and the hemispheres of the spinocerebellum project to the brainstem areas concerned with control of distal limb muscles (lateral brainstem pathways). The lateral portions of the cerebellar hemispheres (cerebrocerebellum) are the newest from a phylogenetic point of view, reaching their greatest development in humans. They interact with the motor cortex in planning and programming movements.
بخشهای عملکردی
از نقطه نظر عملکردی، مخچه به سه قسمت تقسیم میشود (شکل 12-19). گره در ورمیس و لخته کناری در نیمکره در هر طرف، دهلیزی مخچه (یا لوب فلوکولونودولار) را تشکیل میدهند. این لوب از نظر فیلوژنتیکی قدیمیترین قسمت مخچه است و دارای اتصالات دهلیزی است که به تعادل و حرکات چشم مربوط میشود. بقیه ورمیس و بخشهای داخلی مجاور نیمکرهها مخچه نخاعی را تشکیل میدهند، ناحیهای که ورودی حس عمقی را از بدن و همچنین یک کپی از “طرح حرکتی” از قشر حرکتی دریافت میکند. با مقایسه پلان با عملکرد، حرکاتی را که در حال انجام هستند هموار و هماهنگ میکند. ورمیس به ناحیه ساقه مغز مربوط به کنترل عضلات محوری و پروگزیمال اندام (مسیرهای ساقه مغز میانی، و نیمکرههای مخچه نخاعی به نواحی ساقه مغز مربوط به کنترل عضلات انتهایی اندام (مسیرهای ساقه مغز جانبی) میرود. آنها در برنامه ریزی و برنامه ریزی حرکات با قشر حرکتی تعامل دارند.

FIGURE 12-19 Three functional divisions of the cerebellum. The nodulus in the vermis and the flanking flocculus in the hemisphere on each side form the vestibulocerebellum which has vestibular connections and is concerned with equilibrium and eye movements. The rest of the vermis and the adjacent medial portions of the hemispheres form the spinocerebellum, the region that receives proprioceptive input from the body as well as a copy of the “motor plan” from the motor cortex. The lateral portions of the cerebellar hemispheres are called the cerebrocerebellum which interacts with the motor cortex in planning and programming movements. (Modified with permission from Kandel ER, Schwartz JH, Jessell TM (eds): Principles of Neural Science, 4th ed. New York,NY: McGraw-Hill; 2000.)
شکل 12-19 سه بخش عملکردی مخچه. گره در ورمیس و لختههای کناری در نیمکره در هر طرف، دهلیزی مخچه را تشکیل میدهند که دارای اتصالات دهلیزی است و به تعادل و حرکات چشم مربوط میشود. بقیه ورمیس و بخشهای داخلی مجاور نیمکرهها مخچه نخاعی را تشکیل میدهند، ناحیهای که ورودی حس عمقی را از بدن و همچنین یک کپی از “طرح حرکتی” از قشر حرکتی دریافت میکند. قسمتهای جانبی نیمکرههای مخچه را مخچه مینامند که در برنامه ریزی و برنامه ریزی حرکات با قشر حرکتی تعامل دارد. (اصلاح شده با اجازه Kandel ER، Schwartz JH، Jessell TM (eds): Principles of Neural Science، ویرایش چهارم نیویورک، نیویورک: McGraw-Hill؛ 2000.)
Most of the vestibulocerebellar output passes directly to the brainstem; but the rest of the cerebellar cortex projects to the deep nuclei that then project to the brainstem. The deep nuclei provide the only output for the spinocerebellum and the cerebrocerebellum. The medial portion of the spinocerebellum projects to the fastigial nuclei and from there to the brainstem. The adjacent hemispheric portions of the spinocerebellum project to the emboliform and globose nuclei and from there to the brainstem. The cerebrocerebellum projects to the dentate nucleus and from there either directly or indirectly to the ventrolateral nucleus of the thalamus.
بیشتر خروجی دهلیزی به طور مستقیم به ساقه مغز میرسد. اما بقیه قشر مخچه به هستههای عمیق میرسند و سپس به ساقه مغز میرسند. هستههای عمیق تنها خروجی را برای مخچه نخاعی و مخچه مغز فراهم میکنند. قسمت میانی مخچه نخاعی به سمت هستههای فاستیژال و از آنجا به ساقه مغز میرود. بخشهای نیمکره مجاور خارخچه به هستههای آمبولیفرم و کروی و از آنجا به ساقه مغز میرسد. مخچه به سمت هسته دندانه دار و از آنجا به طور مستقیم یا غیرمستقیم به هسته بطنی جانبی تالاموس میرود.
CEREBELLAR DISEASE
Damage to the cerebellum leads to several characteristic abnormalities, including hypotonia, ataxia, and intention tremor. Motor abnormalities associated with cerebellar damage vary depending on the region involved. Figure 12-20 illustrates some of these abnormalities. Additional information is provided in Clinical Box 12-9.
بیماری مخچه
آسیب به مخچه منجر به چندین ناهنجاری مشخصه از جمله هیپوتونی، آتاکسی و لرزش قصد میشود. ناهنجاریهای حرکتی مرتبط با آسیب مخچه بسته به منطقه درگیر متفاوت است. شکل 12-20 برخی از این ناهنجاریها را نشان میدهد. اطلاعات تکمیلی در جعبه بالینی 12-9 ارائه شده است.
FIGURE 12-20 Typical defects associated with cerebellar disease. A) Lesion of the right cerebellar hemisphere delays initiation of movement. The patient is told to clench both hands simultaneously; right hand clenches later than left (shown by recordings from a pressure bulb transducer squeezed by the patient). B) Dysmetria and decomposition of movement shown by patient moving his arm from a raised position to his nose. Tremor increases on approaching the nose. C) Dysdiadochokinesia occurs in the abnormal position trace of hand and forearm as a cerebellar subject tries alternately to pronate and supinate forearm while flexing and extending elbow as rapidly as possible. (Used with permission from Kandel ER, Schwartz JH, Jessell TM (eds): Principles of Neural Science, 4th ed. New York, NY: McGraw-Hill; 2000.)
شکل 12-20 نقصهای معمول مرتبط با بیماری مخچه. الف) ضایعه نیمکره مخچه راست شروع حرکت را به تاخیر میاندازد. به بیمار گفته میشود که هر دو دست خود را همزمان ببندد. دست راست دیرتر از سمت چپ فشرده میشود (با ضبطهای یک مبدل لامپ فشار که توسط بیمار فشرده شده نشان داده میشود). ب) دیسمتری و تجزیه حرکت نشان داده شده توسط بیمار که دست خود را از حالت برآمده به سمت بینی حرکت میدهد. لرزش با نزدیک شدن به بینی افزایش مییابد. ج) دیسدیادوکوکینزی در وضعیت غیرطبیعی دست و ساعد رخ میدهد، زیرا سوژه مخچهای سعی میکند به طور متناوب ساعد را به صورت پرونیت و خوابیده درآورد در حالی که آرنج را با حداکثر سرعت ممکن خم میکند و دراز میکند. (استفاده با اجازه از Kandel ER، Schwartz JH، Jessell TM (eds): Principles of Neural Science، نسخه چهارم نیویورک، نیویورک: McGraw-Hill؛ 2000.)
THE CEREBELLUM & LEARNING
The cerebellum is concerned with learned adjustments that make coordination easier when a given task is performed over and over. As a motor task is learned, activity in the brain shifts from the prefrontal areas to the parietal and motor cortex and the cerebellum. The basis of the learning in the cerebellum is probably the input via the olivary nuclei. The mossy fiber-granule cell-Purkinje cell pathway is highly divergent, allowing an individual Purkinje cell to receive input from many mossy fibers arising from different regions. In contrast, a Purkinje cell receives input from a single climbing fiber but it makes 2000-3000 synapses on it. Climbing fiber activation produces a large, complex spike in the Purkinje cell, and this spike produces long-term modification of the pattern of mossy fiber input to that particular Purkinje cell. Climbing fiber activity is increased when a new movement is being learned, and selective lesions of the olivary complex abolish the ability to produce long-term adjustments in certain motor responses.
مخچه و یادگیری
مخچه به تنظیمات آموخته شده مربوط میشود که هماهنگی را زمانی که یک کار معین بارها و بارها انجام میشود، آسان تر میکند. همانطور که یک کار حرکتی آموخته میشود، فعالیت در مغز از نواحی پیش پیشانی به قشر جداری و حرکتی و مخچه تغییر میکند. اساس یادگیری در مخچه احتمالاً ورودی از طریق هستههای زیتون است. مسیر سلولی فیبر-گرانول خزه ای- سلول پورکنژ بسیار واگرا است و به یک سلول پورکنژ اجازه میدهد تا ورودی بسیاری از الیاف خزه ای را که از مناطق مختلف به وجود میآیند، دریافت کند. در مقابل، یک سلول پورکنژ ورودی را از یک فیبر بالارونده دریافت میکند اما 2000-3000 سیناپس روی آن ایجاد میکند. فعالسازی فیبر صعودی یک سنبله بزرگ و پیچیده در سلول پورکنژ ایجاد میکند و این سنبله اصلاح طولانیمدت الگوی ورودی فیبر خزهای به آن سلول پورکنژ خاص را ایجاد میکند. فعالیت فیبر بالا رفتن زمانی افزایش مییابد که یک حرکت جدید یاد میشود و ضایعات انتخابی مجتمع زیتون توانایی ایجاد تنظیمات طولانی مدت در پاسخهای حرکتی خاص را از بین میبرد.
CLINICAL BOX 12.9
Cerebellar Disease
Most abnormalities associated with damage to the cerebellum are apparent during movement. The marked ataxia is characterized as incoordination due to errors in the rate, range, force, and direction of movement. Ataxia is manifest not only in the wide-based, unsteady, “drunken” gait of patients but also in defects of the skilled movements involved in the production of speech, so that slurred, scanning speech results. Many types of ataxia are hereditary, including Friedreich ataxia and Machado-Joseph disease. There is no cure for hereditary ataxias. Voluntary movements are also highly abnormal when the cerebellum is damaged. For example, attempting to touch an object with a finger results in overshooting. This dysmetria promptly initiates a gross corrective action, but the correction overshoots to the other side, and the finger oscillates back and forth. This oscillation is called an intention tremor. Another characteristic of cerebellar disease is the inability to stop movement promptly. Normally, for example, flexion of the forearm against resistance is quickly checked when the resistance force is suddenly broken off. A patient with cerebellar disease cannot stop the movement of the limb, and the forearm flies backward in a wide arc. This abnormal response is known as the rebound phenomenon and is one of the reasons these patients show dysdiadochokinesia, the inability to perform rapidly alternating opposite movements such as repeated pronation and supination of the hands. Finally, patients with cerebellar disease have difficulty performing actions that involve simultaneous motion at more than one joint. They dissect such movements and carry them out one joint at a time, a phenomenon known as decomposition of movement. Other signs of cerebellar deficit in humans point to the importance of the cerebellum in the control of movement. Motor abnormalities associated with cerebellar damage vary depending on the region involved. The major dysfunction seen after damage to the vestibulocerebellum is ataxia, disequilibrium, and nystagmus. Damage to the vermis and fastigial nucleus (part of the spinocerebellum) leads to scanning speech and disturbances in the control of axial and trunk muscles during attempted antigravity postures. Degeneration of this portion of the cerebellum can result from thiamine deficiency in alcoholics or malnourished individuals. The major dysfunction seen after damage to the cerebrocerebellum is a delay in initiating movements and decomposition of movement.
جعبه بالینی 12.9
بیماری مخچه
اکثر ناهنجاریهای مرتبط با آسیب مخچه در طول حرکت آشکار میشوند. آتاکسی مشخص به عنوان ناهماهنگی به دلیل خطا در سرعت، برد، نیرو و جهت حرکت مشخص میشود. آتاکسی نه تنها در راه رفتن گسترده، ناپایدار و “مست” بیماران، بلکه در نقص حرکات ماهرانه دخیل در تولید گفتار آشکار میشود، به طوری که گفتار نامفهوم و اسکن میشود. بسیاری از انواع آتاکسی ارثی هستند، از جمله آتاکسی فریدریش و بیماری ماچادو جوزف. هیچ درمانی برای آتاکسی ارثی وجود ندارد. هنگامیکه مخچه آسیب میبیند، حرکات ارادی نیز بسیار غیر طبیعی است. به عنوان مثال، تلاش برای لمس یک شی با انگشت منجر به شتاب بیش از حد میشود. این دیسمتری به سرعت یک اقدام اصلاحی فاحش را آغاز میکند، اما اصلاح به سمت دیگر میرود و انگشت به جلو و عقب نوسان میکند. این نوسان را لرزش قصدی مینامند. یکی دیگر از ویژگیهای بیماری مخچه عدم توانایی در توقف سریع حرکت است. به طور معمول، به عنوان مثال، خم شدن ساعد در برابر مقاومت به سرعت بررسی میشود که نیروی مقاومت به طور ناگهانی قطع شود. بیمار مبتلا به بیماری مخچه نمیتواند حرکت اندام را متوقف کند و ساعد در یک قوس پهن به عقب پرواز میکند. این پاسخ غیرطبیعی به عنوان پدیده برگشتی شناخته میشود و یکی از دلایلی است که این بیماران دیس دیادوکوکینزی را نشان میدهند، ناتوانی در انجام سریع حرکات متناوب متناوب مانند پروناسیون مکرر و خوابیدن دستها. در نهایت، بیماران مبتلا به بیماری مخچه در انجام اعمالی که شامل حرکت همزمان در بیش از یک مفصل است، مشکل دارند. آنها چنین حرکاتی را تشریح میکنند و آنها را یکی یکی انجام میدهند، پدیده ای که به عنوان تجزیه حرکت شناخته میشود. سایر علائم کمبود مخچه در انسان به اهمیت مخچه در کنترل حرکت اشاره دارد. ناهنجاریهای حرکتی مرتبط با آسیب مخچه بسته به منطقه درگیر متفاوت است. اختلال عمده ای که پس از آسیب به دهلیزی مشاهده میشود، آتاکسی، عدم تعادل و نیستاگموس است. آسیب به ورمیس و هسته فاستیژال (بخشی از مخچه نخاعی) منجر به اسکن گفتار و اختلال در کنترل عضلات محوری و تنه در هنگام تلاش برای پوسچرهای ضد جاذبه میشود. انحطاط این قسمت از مخچه میتواند ناشی از کمبود تیامین در افراد الکلی یا افراد دارای سوءتغذیه باشد. اختلال عمده ای که پس از آسیب به مخچه مشاهده میشود، تاخیر در شروع حرکات و تجزیه حرکت است.
THERAPEUTIQ HIGHLIGHTS
Management of ataxia is primarily supportive and often includes physical, occupational, and speech therapy. Attempts to identify effective drug therapies have met with little success. DBS of the ventral intermediate nucleus of the thalamus may reduce cerebellar tremor, but it is less effective in reducing ataxia. A deficiency in coenzyme Q10 (CoQ10) may contribute to the abnormalities seen in some forms of familial ataxia. If low levels of CoQ10 are detected, treatment to replace the missing CoQ10 has been beneficial.
نکات برجسته درمانی
مدیریت آتاکسی در درجه اول حمایتی است و اغلب شامل درمان فیزیکی، شغلی و گفتار درمانی است. تلاشها برای شناسایی درمانهای دارویی موثر با موفقیت اندکی مواجه شده است. DBS هسته میانی شکمیتالاموس ممکن است لرزش مخچه را کاهش دهد، اما در کاهش آتاکسی کمتر موثر است. کمبود کوآنزیم Q10 (CoQ10) ممکن است به ناهنجاریهای دیده شده در برخی از اشکال آتاکسی خانوادگی کمک کند. اگر سطوح پایین CoQ10 تشخیص داده شود، درمان برای جایگزینی CoQ10 از دست رفته مفید بوده است.
CHAPTER SUMMARY
خلاصه فصل
• The basic unit of integrated reflex activity is the reflex arc that consists of a sense organ, an afferent neuron, one or more synapses within a central integrating station, an efferent neuron, and an effector.
• واحد اصلی فعالیت رفلکس یکپارچه، قوس بازتابی است که از یک اندام حسی، یک نورون آوران، یک یا چند سیناپس در یک ایستگاه ادغام مرکزی، یک نورون وابران و یک عامل تشکیل شده است.
• A muscle spindle is a group of specialized intrafusal muscle fibers with contractile polar ends and a noncontractile center that is parallel to the extrafusal muscle fibers and innervated by types Ia and II afferent fibers and efferent y-motor neurons.
• دوک عضلانی گروهی از فیبرهای عضلانی داخل فیوزال تخصصی با انتهای قطبی منقبض و یک مرکز غیر انقباضی است که موازی با فیبرهای عضلانی خارج رحمیاست و توسط فیبرهای آوران نوع Ia و II و نورونهای حرکتی y وابران عصب دهی میشود.
• Tapping the patellar tendon causes muscle stretch that activates type Ia fibers that synapse directly on the motor neurons to the extrafusal muscle fibers in the same muscle to cause it to contract (knee jerk reflex).
• ضربه زدن به تاندون کشکک باعث کشش عضلانی میشود که فیبرهای نوع Ia را فعال میکند که مستقیماً روی نورونهای حرکتی به فیبرهای عضلانی خارج رحمیدر همان عضله سیناپس میشوند و باعث انقباض آن میشوند (رفلکس تکان دادن زانو).
• Activation of y-motor neurons causes the contractile ends of the intrafusal fibers to shorten and thus stretches the nuclear bag portion of the spindles, deforming the endings, and initiating impulses in Ia fibers. This can lead to reflex contraction of the muscle. Thus, the y-motor neurons can initiate contraction indirectly via the stretch reflex.
• فعال شدن نورونهای y-motor باعث کوتاه شدن انتهای انقباضی فیبرهای داخل ریز و در نتیجه کشش بخش کیسه هسته ای دوکها، تغییر شکل انتهایی و شروع تکانهها در فیبرهای Ia میشود. این میتواند منجر به انقباض رفلکس عضله شود. بنابراین، نورونهای y-motor میتوانند انقباض را به طور غیر مستقیم از طریق رفلکس کشش آغاز کنند.
• A Golgi tendon organ is a netlike collection of knobby nerve endings among the fascicles of a tendon that is in series with extrafusal muscle fibers and innervated by type Ib afferents. They are stimulated by both passive stretch and active contraction of the muscle to relax the muscle (inverse stretch reflex) and function as a transducer to regulate muscle force.
• اندام تاندون گلژی مجموعه ای شبکه مانند از انتهای عصب دستگیره ای در میان فاسیکلهای یک تاندون است که به صورت سری با رشتههای عضلانی خارج رحمیاست و توسط آورانهای نوع Ib عصب دهی میشود. آنها با کشش غیرفعال و انقباض فعال عضله تحریک میشوند تا عضله را آرام کنند (رفلکس کشش معکوس) و به عنوان یک مبدل برای تنظیم نیروی عضلانی عمل میکنند.
• Physiologic tremor is a rapid (10 Hz), low-amplitude transient tremor in the limbs of healthy individuals; it can become accentuated with anxiety. Clonus is the occurrence of regular, repetitive, rhythmic contractions of a muscle subjected to sudden, maintained stretch; clonus with five or more beats is considered abnormal. Muscle tone is the resistance to muscle stretch. If the muscle offers very little resistance and is said to be flaccid; if resistance to stretch is high, the muscle is spastic. Damage to the cerebral cortex before or during childbirth or during the first 2-3 years of development can lead to cerebral palsy, a disorder that affects muscle tone, movement, and coordination.
• لرزش فیزیولوژیک یک لرزش گذرا سریع (10 هرتز) با دامنه کم در اندامهای افراد سالم است. میتواند با اضطراب برجسته شود. کلونوس وقوع انقباضات منظم، مکرر و ریتمیک عضله ای است که تحت کشش ناگهانی و مداوم قرار میگیرد. کلونوس با پنج ضربه یا بیشتر غیر طبیعی در نظر گرفته میشود. تون عضلانی مقاومت در برابر کشش عضلانی است. اگر عضله مقاومت بسیار کمیاز خود نشان دهد و گفته شود که شل است. اگر مقاومت در برابر کشش زیاد باشد، عضله اسپاستیک است. آسیب به قشر مغز قبل یا در حین زایمان یا در طی 2 تا 3 سال اول رشد میتواند منجر به فلج مغزی شود، اختلالی که بر تون، حرکت و هماهنگی عضلات تأثیر میگذارد.
• The flexor withdrawal reflex is a polysynaptic reflex that is initiated by nociceptive stimuli; it can serve as a protective mechanism to present further injury.
• رفلکس خروج فلکسور یک رفلکس پلی سیناپسی است که توسط محرکهای درد شروع میشود. میتواند به عنوان یک مکانیسم محافظتی برای ایجاد آسیب بیشتر عمل کند.
• Spinal cord injury is followed by a period of spinal shock during which all spinal reflex responses are depressed. In humans, recovery begins about 2weeks after the injury.
• آسیب نخاعی با یک دوره شوک نخاعی همراه است که در طی آن تمام واکنشهای رفلکس نخاعی افسرده میشوند. در انسان، بهبودی حدود 2 هفته پس از آسیب شروع میشود.
• The supplementary cortex, basal ganglia, and cerebellum participate in the planning of skilled movements. Commands from the primary motor cortex and other cortical regions are relayed via the corticospinal and corticobulbar tracts to spinal and brainstem motor neurons. Cortical areas and motor pathways descending from the cortex are somatotopically organized.
• قشر مکمل، عقدههای قاعده ای و مخچه در برنامه ریزی حرکات ماهرانه شرکت میکنند. دستورات قشر حرکتی اولیه و سایر نواحی قشر مغز از طریق مجاری قشر نخاعی و کورتیکوبولبار به نورونهای حرکتی نخاعی و ساقه مغز منتقل میشوند. نواحی کورتیکال و مسیرهای حرکتی که از قشر فرو میروند به صورت سوماتوتوپی سازماندهی میشوند.
• The ventral corticospinal tract and medial descending brainstem pathways (tectospinal, reticulospinal, and vestibulospinal tracts) regulate proximal muscles and posture. The lateral corticospinal and rubrospinal tracts control distal limb muscles for fine motor control and skilled voluntary movements.
• دستگاه قشر نخاعی شکمیو مسیرهای نزولی ساقه مغز میانی (تکتو نخاعی، شبکه نخاعی و دهلیزی نخاعی) عضلات پروگزیمال و وضعیت بدن را تنظیم میکنند. مجاری جانبی قشر نخاعی و روبروسنخاعی عضلات انتهایی اندام را برای کنترل حرکتی ظریف و حرکات ارادی ماهرانه کنترل میکنند.
• Lower motor neurons are those whose axons terminate on skeletal muscles; damage to these neurons (eg, ALS) is associated with flaccid paralysis, muscular atrophy, fasciculations, hypotonia, and hyporeflexia or areflexia. Upper motor neurons include corticospinal tract neurons that innervate spinal motor neurons; damage to these neurons initially causes muscles to become weak and flaccid but eventually leads to spasticity, hypertonia, hyperactive stretch reflexes, and a positive Babinski sign.
• نورونهای حرکتی تحتانی آنهایی هستند که آکسون آنها به ماهیچههای اسکلتی ختم میشود. آسیب به این نورونها (به عنوان مثال، ALS) با فلج شل، آتروفی عضلانی، فاسیکلاسیون، هیپوتونی، و هیپورفلکسی یا آرفلکسی همراه است. نورونهای حرکتی فوقانی شامل نورونهای دستگاه قشر نخاعی هستند که نورونهای حرکتی نخاعی را عصب دهی میکنند. آسیب به این نورونها در ابتدا باعث ضعیف شدن و شل شدن ماهیچهها میشود اما در نهایت منجر به اسپاسم، هیپرتونی، رفلکسهای کششی بیش فعال و علامت بابینسکی مثبت میشود.
• Damage to the cerebral cortex before or during childbirth can lead to cerebral palsy, a disorder that affects muscle tone, movement, and coordination. Decerebrate rigidity leads to hyperactivity in extensor muscles in all four extremities; the spasticity is due to facilitation of the stretch reflex. It resembles what occurs with uncal herniation due to a supratentorial lesion. Decorticate rigidity is flexion of the upper extremities at the elbow and extensor hyperactivity in the lower extremities. It occurs on the hemiplegic side after hemorrhage or thrombosis in the internal capsule.
• آسیب به قشر مغز قبل یا در حین زایمان میتواند منجر به فلج مغزی شود، اختلالی که بر تون، حرکت و هماهنگی عضلات تأثیر میگذارد. سفتی ضعیف منجر به بیش فعالی در عضلات بازکننده در هر چهار اندام میشود. اسپاستیسیته به دلیل تسهیل رفلکس کشش است. این شبیه همان چیزی است که با فتق انکال به دلیل یک ضایعه فوق تنتوریال رخ میدهد. سفتی تزئینی خم شدن اندام فوقانی در آرنج و بیش فعالی اکستانسور در اندام تحتانی است. در سمت همیپلژیک پس از خونریزی یا ترومبوز در کپسول داخلی رخ میدهد.
• The basal ganglia include the caudate nucleus, putamen, globus pallidus, subthalamic nucleus, and substantia nigra. The connections between the parts of the basal ganglia include a dopaminergic nigrostriatal projection from the substantia nigra to the striatum and a GABAergic projection from the striatum to substantia nigra.
• عقدههای قاعده ای شامل هسته دمی، پوتامن، گلوبوس پالیدوس، هسته ساب تالاموس و ماده سیاه است. اتصالات بین قسمتهای عقدههای قاعدهای شامل یک برآمدگی سیاهدانه دوپامینرژیک از جسم سیاه به جسم مخطط و یک برآمدگی GABAergic از جسم مخطط به جسم سیاه است.
• Parkinson disease is due to degeneration of the nigrostriatal dopaminergic neurons and is characterized by akinesia, bradykinesia, cogwheel rigidity, and tremor at rest. Huntington disease is characterized by choreiform movements due to the loss of the GABAergic inhibitory pathway to the globus pallidus.
• بیماری پارکینسون به دلیل انحطاط نورونهای دوپامینرژیک سیاه پوست است و با آکینزی، برادی کینزی، سفتی چرخ دنده و لرزش در حالت استراحت مشخص میشود. بیماریهانتینگتون با حرکات کوریفرم به دلیل از بین رفتن مسیر مهاری GABAergic به گلوبوس پالیدوس مشخص میشود.
• The cerebellar cortex contains five types of neurons: Purkinje, granule, basket, stellate, and Golgi cells. The two main inputs to the cerebellar cortex are climbing fibers and mossy fibers. Purkinje cells are the only output from the cerebellar cortex, and they generally project to the deep nuclei. Damage to the cerebellum leads to several characteristic abnormalities, including hypotonia, ataxia, and intention tremor.
• قشر مخچه حاوی پنج نوع نورون است: سلولهای پورکنژ، گرانول، سبد، ستاره ای و گلژی. دو ورودی اصلی به قشر مخچه فیبرهای بالا رونده و الیاف خزه ای هستند. سلولهای پورکنژ تنها خروجی از قشر مخچه هستند و عموماً به هستههای عمیق میرسند. آسیب به مخچه منجر به چندین ناهنجاری مشخصه از جمله هیپوتونی، آتاکسی و لرزش قصد میشود.
MULTIPLE-CHOICE QUESTIONS
For all questions, select the single best answer unless otherwise directed.
سوالات چند گزینه ای
برای همه سؤالات، بهترین پاسخ را انتخاب کنید، مگر اینکه دستور دیگری داده شود.
1. A 48-year-old man was undergoing a thorough neurological exam after falling from a construction platform. The test included an evaluation of his deep tendon reflex. What are the elements of the deep tendon reflex?
A. Golgi tendon organ, group Ib afferent fibers, spinal inhibitory interneuron, contralateral a-motor neuron, and extrafusal fibers of the skeletal muscle
B. Golgi tendon organ, group II afferent fibers, spinal inhibitory interneuron, ipsilateral a-motor neuron, and intrafusal fibers of the skeletal muscle
C. Group Ia sensory fibers originating in the contractile portion of the intrafusal fibers of the muscle spindle, spinal dorsal horn neuron, ipsilateral a-motor neuron, and extrafusal fibers of the skeletal muscle
D. Group la sensory fibers originating in the central portion of the intrafusal fibers of the muscle spindle, ipsilateral a-motor neuron, and extrafusal fibers of the skeletal muscle
E. Group II sensory fibers originating in the contractile portion of the intrafusal fibers of the muscle spindle, spinal inhibitory interneuron, ipsilateral a-motor neuron, and extrafusal fibers of the skeletal muscle
1. مردی 48 ساله پس از سقوط از سکوی ساختمانی تحت معاینه کامل اعصاب بود. این آزمایش شامل ارزیابی رفلکس تاندون عمیق او بود. عناصر رفلکس تاندون عمیق کدامند؟
الف. اندام تاندون گلژی، الیاف آوران گروه Ib، نورون بازدارنده نخاعی، نورون حرکتی طرف مقابل و فیبرهای خارج رحمیعضله اسکلتی
ب. اندام تاندون گلژی، فیبرهای آوران گروه دوم، نورون بازدارنده نخاعی، نورون حرکتی الف همان طرف و فیبرهای داخل فیوزال عضله اسکلتی
ج. الیاف حسی گروه Ia که از قسمت انقباضی فیبرهای داخل فیوزال دوک عضلانی، نورون شاخ پشتی نخاعی، نورون a-حرکتی همان طرف و الیاف خارج رحمیعضله اسکلتی منشا میگیرند.
د. گروه الیاف حسی la که از بخش مرکزی الیاف داخل فیوزال دوک عضلانی، نورون a-حرکتی همان طرف و الیاف خارج رحمیعضله اسکلتی منشا میگیرند.
ی. فیبرهای حسی گروه دوم که از قسمت انقباضی فیبرهای داخل ریز دوک عضلانی، نورون بازدارنده نخاعی، نورون a-حرکتی همان طرف، و فیبرهای خارج رحمیعضله اسکلتی منشا میگیرند.
2. When dynamic y-motor neurons are activated at the same time as a-motor neurons to muscle,
A. prompt inhibition of discharge in spindle Ia afferents takes place.
B. clonus is likely to occur.
C. the muscle will not contract.
D. the number of impulses in spindle Ia afferents is smaller than when a discharge alone is increased.
E. the number of impulses in spindle Ia afferents is greater than when a discharge alone is increased.
2. هنگامیکه نورونهای y-motor پویا همزمان با نورونهای a-motor به عضله فعال میشوند،
الف. مهار سریع تخلیه در آورانهای دوکی Ia صورت میگیرد.
ب. clonus به احتمال زیاد رخ میدهد.
ج. عضله منقبض نخواهد شد.
د. تعداد تکانهها در آورانهای دوکی Ia کمتر از زمانی است که یک ترشح به تنهایی افزایش مییابد.
ی. تعداد ایمپالسها در آورانهای دوکی Ia بیشتر از زمانی است که یک ترشح به تنهایی افزایش مییابد.
3. The inverse stretch reflex
A. occurs when Ia spindle afferents are inhibited.
B. is a monosynaptic reflex initiated by activation of the Golgi tendon organ.
C. is a disynaptic reflex with a single interneuron inserted between the afferent and efferent limbs.
D. is a polysynaptic reflex with many interneurons inserted between the afferent and efferent limbs.
E. uses type II afferent fibers from the Golgi tendon organ.
3. رفلکس کشش معکوس
الف. زمانی رخ میدهد که آورانهای دوکی Ia مهار شوند.
ب. یک رفلکس تک سیناپسی است که با فعال شدن اندام تاندون گلژی آغاز میشود.
ج. یک رفلکس دی سیناپسی با یک نورون منفرد است که بین اندامهای آوران و وابران قرار میگیرد.
د. یک رفلکس پلی سیناپسی با تعداد زیادی نورون بین اندام آوران و وابران است.
ی. از الیاف آوران نوع II از اندام تاندون گلژی استفاده میکند.
4. Withdrawal reflexes are
A. initiated by innocuous stimulation of the skin.
B. can lead to the appearance of clonus.
C. prolonged if the stimulus is strong.
D. an example of a stretch reflex.
E. accompanied by the same response on both sides of the body.
4. رفلکسهای خروج هستند
الف. با تحریک بی ضرر پوست شروع میشود.
ب. میتواند منجر به ظهور کلونوس شود.
ج. اگر محرک قوی باشد طولانی شود.
د. مثالی از رفلکس کششی.
ی. همراه با پاسخ یکسان در دو طرف بدن.
5. While exercising, a 42-year-old woman developed sudden onset of tingling in her right leg and an inability to control movement in that limb. A neurologic exam showed a hyperactive knee jerk reflex and a positive Babinski sign. What is a possible basis for these findings and does it reflect damage to an upper or lower motor neuron
A. She had a lower thoracic disk rupture that damaged the right side of her spinal cord (upper motor neuron damage).
B. She had a mid-cervical disk rupture that damaged the left side of her spinal cord (upper motor neuron damage).
C. She had a lower lumbar disk rupture that compressed the spinal nerve at that segmental level (lower motor neuron damage).
D. She had a sacral disk rupture that put pressure on the ventral root at that segmental level (lower motor neuron damage).
E. She was experiencing dysfunction of both a lower motor neuron and an upper motor neuron.
5. زن 42 ساله ای در حین ورزش، دچار شروع ناگهانی گزگز در پای راست و ناتوانی در کنترل حرکت در آن اندام شد. یک معاینه عصبی یک رفلکس تکان دادن بیش فعال زانو و یک علامت بابینسکی مثبت را نشان داد. مبنای احتمالی این یافتهها چیست و آیا آسیب به نورون حرکتی فوقانی یا تحتانی را منعکس میکند؟
الف. او دچار پارگی دیسک پایین قفسه سینه شده بود که به سمت راست نخاعش آسیب رساند (آسیب نورون حرکتی فوقانی).
ب. او دچار پارگی دیسک میانی گردن شده بود که به سمت چپ نخاع آسیب رساند (آسیب نورون حرکتی فوقانی).
ج. او دچار پارگی دیسک کمر تحتانی بود که عصب نخاعی را در آن سطح سگمنتال تحت فشار قرار داد (آسیب نورون حرکتی تحتانی).
د. او دچار پارگی دیسک خاجی بود که بر ریشه شکمیدر آن سطح سگمنتال فشار وارد کرد (آسیب نورون حرکتی تحتانی).
ی. او اختلال عملکرد یک نورون حرکتی تحتانی و یک نورون حرکتی فوقانی را تجربه میکرد.
6. Increased neural activity before a skilled voluntary movement is first seen in the
A. spinal motor neurons.
B. precentral motor cortex.
C. midbrain.
D. cerebellum.
E. cortical association areas.
6. افزایش فعالیت عصبی قبل از اینکه یک حرکت ارادی ماهرانه برای اولین بار در آن دیده شود
الف. نورونهای حرکتی نخاعی.
ب. قشر حرکتی پیش مرکزی.
ج. مغز میانی.
د. مخچه.
ی. مناطق انجمن قشر.
7. A 58-year-old woman was brought to the emergency department of her local hospital because of a sudden change of consciousness. All four limbs were extended, suggestive of decerebrate rigidity. A brain CT showed a rostral pontine hemorrhage. What are the underlying neurophysiological changes that lead to the appearance of decerebrate rigidity?
A. Destruction of the rubrospinal tract eliminates inhibition of the cerebellar fastigial nucleus and secondarily increases excitation to vestibular nuclei to activate extensor muscles in the limbs.
B. Loss of the corticospinal pathway disrupts excitatory input to motor neurons controlling flexor muscles, leaving extensor muscles to undergo sustained contraction.
C. Sensory input activates the medullary reticulospinal pathway, which then directly activates motor neurons to extensor muscles in all four extremities.
D. Sensory input activates neurons in the rubrospinal tract that inhibit flexor a-motor neurons and excite extensor a-motor neurons in all four limbs.
E. Sensory input activates the pontine reticulospinal pathway, which then activates primarily y-motor neurons to extensor muscles in all four extremities.
7. یک زن 58 ساله به دلیل تغییر ناگهانی هوشیاری به بخش اورژانس بیمارستان محل خود آورده شد. هر چهار اندام گسترش یافته بودند، که حاکی از سفتی ضعیف است. سی تی مغزی خونریزی منقاری پونتین را نشان داد. تغییرات نوروفیزیولوژیک زمینه ای که منجر به ظهور سفتی ناخوشایند میشود چیست؟
الف. تخریب دستگاه روبروسنخاعی مهار هسته فاستژیال مخچه را از بین میبرد و به طور ثانویه تحریک به هستههای دهلیزی را افزایش میدهد تا عضلات اکستانسور در اندامها فعال شوند.
ب. از دست دادن مسیر قشر نخاعی، ورودی تحریکی به نورونهای حرکتی کنترل کننده عضلات فلکسور را مختل میکند و عضلات اکستانسور را تحت انقباض پایدار قرار میدهد.
ج. ورودی حسی مسیر شبکهای نخاعی مدولاری را فعال میکند، که سپس نورونهای حرکتی را مستقیماً به عضلات بازکننده در هر چهار اندام فعال میکند.
د. ورودی حسی نورونهایی را در دستگاه روبروسنخاعی فعال میکند که نورونهای حرکتی فلکسور را مهار کرده و نورونهای حرکتی اکستانسور را در هر چهار اندام تحریک میکند.
ی. ورودی حسی مسیر شبکه ای نخاعی پونتین را فعال میکند، که سپس نورونهای حرکتی y را به عضلات اکستانسور در هر چهار اندام فعال میکند.
8. Which of the following correctly describes components of the central pathway responsible for control of posture?
A. The tectospinal pathway originates in the superior colliculus and terminates on neurons in the dorsolateral area of the spinal ventral horn that innervate limb muscles.
B. The medullary reticulospinal pathway terminates on neurons in the ventromedial area of the spinal ventral horn that innervate axial and proximal muscles.
C. The pontine reticulospinal pathway terminates on neurons in the dorsomedial area of the spinal ventral horn that innervate limb muscles.
D. The medial vestibular pathway terminates on neurons in the dorsomedial area of the spinal ventral horn that innervate axial and proximal muscles.
E. The lateral vestibular pathway terminates on neurons in the dorsolateral area of the spinal ventral horn that innervate axial and proximal muscles.
8. کدام یک از موارد زیر اجزای مسیر مرکزی مسئول کنترل وضعیت بدن را به درستی توصیف میکند؟
الف. مسیر تکتو نخاعی از کولیکولوس فوقانی سرچشمه میگیرد و به سلولهای عصبی در ناحیه پشتی جانبی شاخ شکمینخاعی ختم میشود که عضلات اندام را عصب دهی میکند.
ب. مسیر شبکهای نخاعی مدولاری به سلولهای عصبی در ناحیه شکمیمیانی شاخ شکمینخاعی ختم میشود که عضلات محوری و پروگزیمال را عصب دهی میکنند.
ج. مسیر شبکه ای نخاعی پونتین به سلولهای عصبی در ناحیه پشتی میانی شاخ شکمینخاعی ختم میشود که عضلات اندام را عصب دهی میکند.
د. مسیر دهلیزی داخلی به سلولهای عصبی در ناحیه پشتی میانی شاخ شکمینخاعی ختم میشود که عضلات محوری و پروگزیمال را عصب دهی میکنند.
ی. مسیر دهلیزی جانبی به سلولهای عصبی در ناحیه پشتی جانبی شاخ شکمینخاع ختم میشود که عضلات محوری و پروگزیمال را عصب دهی میکند.
9. Which of the following describes a connection between components of the basal ganglia?
A. The subthalamic nucleus releases glutamate to excite the globus pallidus, internal segment.
B. The substantia nigra pars reticulata releases dopamine to inhibit thestriatum.
C. The substantia nigra pars compacta releases dopamine to excite the globus pallidus, external segment.
D. The striatum releases acetylcholine to excite the substantia nigra pars reticulata.
E. The globus pallidus, external segment releases glutamate to excite the striatum.
9. کدام یک از موارد زیر ارتباط بین اجزای عقدههای پایه را توصیف میکند؟
الف. هسته ساب تالاموس گلوتامات را آزاد میکند تا بخش داخلی گلوبوس پالیدوس را تحریک کند.
ب. ماده سیاه پارس رتیکولاتا دوپامین آزاد میکند تا ته استریاتوم را مهار کند.
ج. ماده سیاه پارس فشرده دوپامین آزاد میکند تا گلوبوس پالیدوس، بخش خارجی را تحریک کند.
د. جسم مخطط استیل کولین را برای تحریک ماده سیاه پارس رتیکولاتا آزاد میکند.
ی. گلوبوس پالیدوس، بخش خارجی گلوتامات آزاد میکند تا جسم مخطط را تحریک کند.
10. A 60-year-old man with Parkinson disease, which was diagnosed 15 years ago, has been taking carbidopa and levodopa (Sinemet); until recently, he has been able to continue to work and help with routine jobs around the house. Now his tremor and rigidity interfere with these activities. His clinician has suggested that he undergo deep brain stimulation therapy. The therapeutic effect of L-dopa in patients with Parkinson disease eventually wears off because
A. antibodies to dopamine receptors develop.
B. inhibitory pathways grow into the basal ganglia from the frontal lobe.
C. there is an increase in circulating a-synuclein.
D. the normal action of nerve growth factor (NGF) is disrupted.
E. the dopaminergic neurons in the substantia nigra continue to degenerate.
10. یک مرد 60 ساله مبتلا به بیماری پارکینسون که 15 سال پیش تشخیص داده شد، کاربیدوپا و لوودوپا (سینمت) مصرف میکرد. تا همین اواخر، او میتوانست به کار خود ادامه دهد و در کارهای معمول در خانه کمک کند. اکنون لرزش و سفتی او با این فعالیتها تداخل دارد. پزشک او به او پیشنهاد کرده است که تحت درمان تحریک عمیق مغز قرار گیرد. اثر درمانی L-dopa در بیماران مبتلا به بیماری پارکینسون در نهایت از بین میرود زیرا
الف. آنتی بادی برای گیرندههای دوپامین ایجاد میشود.
ب. مسیرهای مهاری از لوب فرونتال به گانگلیونهای پایه رشد میکنند.
ج. افزایش در گردش a-synuclein وجود دارد.
د. عمل طبیعی فاکتور رشد عصبی (NGF) مختل میشود.
ی. نورونهای دوپامینرژیک در جسم سیاه به انحطاط خود ادامه میدهند.
11. An 8-year-old girl was brought to her pediatrician because her parents noted frequent episodes of gait unsteadiness and speech difficulties. Her mother was concerned because of a family history of Friedreich ataxia. Which of the following is a correct description of connections involving cerebellar neurons?
A. Basket cells release glutamate to activate Purkinje cells.
B. Climbing fiber inputs exert a strong excitatory effect on Purkinje cells, and mossy fiber inputs exert a strong inhibitory effect on Purkinje cells.
C. Granule cells release glutamate to excite basket cells and stellate cells.
D. The axons of Purkinje cells are the sole output of the cerebellar cortex, and they release glutamate to excite the deep cerebellar nuclei.
E. Golgi cells are inhibited by mossy fiber collaterals.
11. یک دختر 8 ساله را نزد پزشک اطفال آوردند زیرا والدینش به دفعات مکرر بی ثباتی در راه رفتن و مشکلات گفتاری اشاره کردند. مادرش به دلیل سابقه خانوادگی آتاکسی فریدریش نگران بود. کدام یک از موارد زیر توصیف صحیحی از اتصالات مربوط به نورونهای مخچه است؟
الف. سلولهای سبد گلوتامات آزاد میکنند تا سلولهای پورکنژ را فعال کنند.
ب. ورودیهای فیبر صعودی یک اثر تحریکی قوی روی سلولهای پورکنژ و ورودیهای فیبر خزه ای اثر بازدارندگی قوی روی سلولهای پورکنژ دارند.
ج. سلولهای گرانول گلوتامات را برای تحریک سلولهای سبد و سلولهای ستاره ای آزاد میکنند.
د. آکسونهای سلولهای پورکنژ تنها خروجی قشر مخچه هستند و گلوتامات را برای تحریک هستههای عمیق مخچه آزاد میکنند.
ی. سلولهای گلژی توسط وثیقههای فیبر خزه ای مهار میشوند.
12. After falling down a flight of stairs, a young woman is found to have partial loss of voluntary movement on the right side of her body and loss of pain and temperature sensation on the left side below the midthoracic region. It is probable that she has a lesion
A. damaging the left half of the spinal cord in the lumbar region.
B. damaging the left half of the spinal cord in the upper thoracic region.
C. damaging sensory and motor pathways on the right side of the pons.
D. damaging the right half of the spinal cord in the upper thoracic region.
E. damaging the dorsal half of the spinal cord in the upper thoracic region.
12. پس از سقوط از پلهها، زن جوانی متوجه شد که حرکت ارادی در سمت راست بدن خود را از دست داده و احساس درد و دما در سمت چپ زیر ناحیه میانی قفسه سینه را از دست داده است. احتمال دارد ضایعه داشته باشد
الف. آسیب رساندن به نیمه چپ نخاع در ناحیه کمر.
ب. آسیب به نیمه چپ نخاع در ناحیه فوقانی قفسه سینه.
ج. آسیب رساندن به مسیرهای حسی و حرکتی در سمت راست پونز.
د. آسیب به نیمه راست نخاع در ناحیه فوقانی قفسه سینه.
ی. آسیب رساندن به نیمه پشتی نخاع در ناحیه فوقانی قفسه سینه.
13. At the age of 30, a male postal worker reported weakness in his right leg. Within a year the weakness had spread to his entire right side. A neurologic examination revealed flaccid paralysis, muscular atrophy, fasciculations, hypotonia, and hyporeflexia of muscles in the right arm and leg. Sensory and cognitive function tests were normal. Which of the following diagnosis is likely?
A. A large tumor in the left primary motor cortex
B. A cerebral infarct in the region of the corona radiate
C. A vestibulocerebellar tumor
D. Damage to the basal ganglia
E. Amyotrophic lateral sclerosis
13. در سن 30 سالگی، یک کارگر پست مرد از ضعف در پای راست خود خبر داد. در عرض یک سال ضعف به تمام سمت راست او سرایت کرده بود. معاینه عصبی فلج شل، آتروفی عضلانی، فاسیکولاسیون، هیپوتونی و هیپورفلکسی عضلات در بازو و پای راست را نشان داد. آزمون عملکرد حسی و شناختی نرمال بود. کدام یک از تشخیصهای زیر محتمل است؟
الف. تومور بزرگ در قشر حرکتی اولیه چپ
ب. انفارکتوس مغزی در ناحیه تشعشع تاج
ج. تومور دهلیزی مخچه
د. آسیب به عقدههای قاعده ای
ی. اسکلروز جانبی آمیوتروفیک
کلیک کنید «منابع»
CHAPTER RESOURCES
Alexi T, Liu X-Z, Qu Y, et al: Neuroprotective strategies for basal ganglia degeneration: Parkinson’s and Huntington’s diseases. Prog Neurbiol 2000;60:409.
De Zeeuw CI, Strata P, Voogd J: The Cerebellum: From Structure to Control. Elsevier, 1997.
Ditunno JF Jr, Formal CF: Chronic spinal cord injury. N Engl J Med 1994; 330:550.
Graybiel AM, Delong MR, Kitai ST: The Basal Ganglia VI. Springer, 2003.
Hunt CC: Mammalian muscle spindle: Peripheral mechanisms. Physiol Rev 1990;70: 643.
Jankowska E: Interneuronal relay in spinal pathways from proprioceptors. Prog Neurobiol 1992;38:335.
Jueptner M, Weiller C: A review of differences between basal ganglia and cerebellar control of movements as revealed by functional imaging studies. Brain 1998;121:1437.
Latash ML: Neurophysiological Basis of Movement, 2nd ed. Human Kinetics, 2008.
Lemon RN: Descending pathways in motor control. Annu Rev Neurosci 2008;31:195.
Lundberg A: Multisensory control of spinal reflex pathways. Prog Brain Res 1979;50:11.
Manto MU, Pandolfo M: The Cerebellum and its Disorders. Cambridge University Press, 2001.
Matyas F, Sreenivasan V, Marbach F, Wacongne C, Barsy B, Mateo C, Aronoff R, Petersen CCH: Motor control of sensory cortex. Science 2010;26:1240.
McDonald JW, Liu X-Z, Qu Y, et al: Transplanted embryonic stem cells survive, differentiate and promote recovery in injured rat spinal cord. Nature Med 1999;5:1410.
Nudo RJ: Postinfarct cortical plasticity and behavioral recovery. Stroke 2007;38:840.
Ramer LM, Ramer MS, Steeves JD: Setting the stage for functional repair of spinal cord injuries: a cast of thousands. Spinal Cord 2005;43:134.
Stein RB, Thompson AK: Muscle reflexes and motion: How, what, and why? Exerc Sport Sci Rev 2006;34:145.
🚀 با ما همراه شوید!
تازهترین مطالب و آموزشهای مغز و اعصاب را از دست ندهید. با فالو کردن کانال تلگرام آیندهنگاران مغز، از ما حمایت کنید!

 ورود/ ثبت نام با جیمیل
ورود/ ثبت نام با جیمیل
