فیزیولوژی پزشکی گانونگ؛ حواس بویایی و چشایی

دعای مطالعه [ نمایش ]
بِسْمِ الله الرَّحْمنِ الرَّحیمِ
اَللّهُمَّ اَخْرِجْنى مِنْ ظُلُماتِ الْوَهْمِ
خدايا مرا بيرون آور از تاريكىهاى وهم،
وَ اَكْرِمْنى بِنُورِ الْفَهْمِ
و به نور فهم گرامى ام بدار،
اَللّهُمَّ افْتَحْ عَلَيْنا اَبْوابَ رَحْمَتِكَ
خدايا درهاى رحمتت را به روى ما بگشا،
وَانْشُرْ عَلَيْنا خَزائِنَ عُلُومِكَ بِرَحْمَتِكَ يا اَرْحَمَ الرّاحِمينَ
و خزانههاى علومت را بر ما باز كن به امید رحمتت اى مهربانترين مهربانان.
کتاب «فیزیولوژی پزشکی گانونگ» بهعنوان یکی از جامعترین و معتبرترین منابع در حوزه علوم پزشکی، همچنان مرجع کلیدی برای درک عملکرد پیچیده بدن انسان است. این اثر با تکیه بر تازهترین پژوهشها و توضیحات دقیق از سازوکارهای فیزیولوژیک، پلی میان علوم پایه پزشکی و کاربردهای بالینی ایجاد میکند و نقشی بیبدیل در آموزش، پژوهش و ارتقای دانش سلامت ایفا مینماید.
ترجمه دقیق و علمی این شاهکار توسط برند علمی آیندهنگاران مغز به مدیریت داریوش طاهری، دسترسی فارسیزبانان به مرزهای نوین دانش فیزیولوژی را ممکن ساخته و رسالتی علمی برای ارتقای آموزش پزشکی، فهم عمیقتر سازوکارهای بدن و توسعه روشهای نوین در حوزه سلامت فراهم آورده است.
» کتاب فیزیولوژی پزشکی گانونگ
» » فصل ۹: بویایی و چشایی
در حال ویرایش
» Ganong’s Review of Medical Physiology
»» CHAPTER 9: CHAPTER 9: Smell & Taste
OBJECTIVES
After studying his chapter, you should be able to:
• Describe the structure and function of the neural elements in the olfactory epithelium and olfactory bulb.
• Identify the significance of the family of olfactory receptor genes.
• Explain how odorant receptors are activated and the mechanism by which signal transduction occurs in these receptors.
• Label the components of the pathway by which impulses generated in the olfactory epithelium reach five regions of the olfactory cortex.
• Describe the location and cellular composition of taste buds.
• Name the five major taste modalities and compare the signal transduction mechanisms in the receptors mediating these different taste modalities.
• Label the components of the pathways by which impulses generated in taste receptors reach the gustatory region of the insular cortex.
Name and discuss abnormalities in odor and taste sensations.
اهداف
پس از مطالعه این فصل، باید بتوانید:
• ساختار و عملکرد عناصر عصبی در اپیتلیوم بویایی و پیاز بویایی را شرح دهید.
• اهمیت خانواده ژنهای گیرنده بویایی را شناسایی کنید.
• نحوه فعال شدن گیرندههای بو و مکانیسم انتقال سیگنال در این گیرندهها را توضیح دهید.
• اجزای مسیری که از طریق آن تکانههای ایجاد شده در اپیتلیوم بویایی به پنج ناحیه قشر بویایی میرسد را علامت گذاری کنید.
• محل و ترکیب سلولی جوانههای چشایی را شرح دهید.
• پنج روش اصلی چشایی را نام ببرید و مکانیسمهای انتقال سیگنال را در گیرندههای واسطه این روشهای مختلف طعم مقایسه کنید.
• اجزای مسیرهایی را که تکانههای تولید شده در گیرندههای چشایی به ناحیه چشایی قشر منزوی میرسند، برچسب بزنید.
ناهنجاریهای موجود در حس بو و مزه را نام ببرید و در مورد آن صحبت کنید.

INTRODUCTION
Smell (olfaction) and taste (gustation) are examples of visceral senses because of their close association with gastrointestinal function. Physiologically, they are related to each other as the flavor of food is a combination of its taste and smell. This is why food may taste “different” if one has a cold that depresses the sense of smell. Smell and taste receptors are chemoreceptors that are stimulated by chemical molecules in solution in mucus in the nose (odorants) and saliva in the mouth (tastants). The sensations of smell and taste likely evolved as protective mechanisms to avoid the intake of potentially harmful substances.
مقدمه
بویایی (بویایی) و چشایی (چشیدن) نمونههایی از حواس احشایی هستند زیرا ارتباط نزدیکی با عملکرد دستگاه گوارش دارند. از نظر فیزیولوژیکی به یکدیگر مرتبط هستند زیرا طعم غذا ترکیبی از طعم و بوی آن است. به همین دلیل است که اگر فرد سرماخورده ای داشته باشد که حس بویایی را تضعیف میکند، ممکن است طعم «متفاوت» داشته باشد. گیرندههای بویایی و چشایی گیرندههای شیمیایی هستند که توسط مولکولهای شیمیایی در محلول موجود در مخاط در بینی (بویندهها) و بزاق در دهان (تستانت) تحریک میشوند. حس بویایی و چشایی احتمالاً به عنوان مکانیسمهای محافظتی برای جلوگیری از مصرف مواد بالقوه مضر تکامل یافته است.
SMELL
OLFACTORY EPITHELIUM
The yellowish pigmented olfactory epithelium is a specialized portion of the nasal mucosa that covers an area of 10 cm2 in the roof of the nasal cavity near the septum in humans (Figure 9-1). The olfactory epithelium is the place in the body where the nervous system is closest to the external world. It contains three types of neurons that are important for olfaction: olfactory sensory neurons, supporting cells, and basal stem cells.
بو
اپیتلیوم بویایی
اپیتلیوم بویایی رنگدانهای مایل به زرد، بخش ویژهای از مخاط بینی است که مساحتی معادل 10 سانتیمتر مربع در سقف حفره بینی نزدیک سپتوم در انسان را پوشش میدهد (شکل 9-1). اپیتلیوم بویایی جایی در بدن است که سیستم عصبی در آن به دنیای خارج نزدیک است. این شامل سه نوع نورون است که برای بویایی مهم هستند: نورونهای حسی بویایی، سلولهای پشتیبان و سلولهای بنیادی پایه.
The bipolar olfactory sensory neurons (also called olfactory receptor cells) are responsible for olfactory transduction. They have a short, thick dendrite that projects into the nasal cavity where it terminates in a knob containing 6-12 cilia that protrude into the thin layer of mucus overlying the epithelium (Figure 9-1). The axons of the olfactory sensory neurons (ie, olfactory nerve) pass through the cribriform plate of the ethmoid bone to enter the olfactory bulbs.
نورونهای حسی بویایی دوقطبی (که سلولهای گیرنده بویایی نیز نامیده میشوند) مسئول انتقال بویایی هستند. آنها دارای یک دندریت کوتاه و ضخیم هستند که به داخل حفره بینی بیرون میزند، جایی که به دستگیره ای حاوی 6-12 مژک ختم میشود که در لایه نازک مخاطی که روی اپیتلیوم قرار دارد بیرون زده است (شکل 9-1). آکسونهای نورونهای حسی بویایی (یعنی عصب بویایی) از صفحه کریبریفرم استخوان اتموئید عبور میکنند تا وارد پیازهای بویایی شوند.
The supporting cells secrete the mucus that provides the appropriate molecular and ionic environment for odor detection in the olfactory epithelium. Odor-producing molecules (odorants) dissolve in the mucus and bind to odorant receptors on the cilia of the olfactory sensory neurons. Odorant-binding proteins in the mucus may facilitate the diffusion of odorants to and from the odorant receptor. Basal stem cells undergo mitosis to generate new olfactory sensory neurons as needed to replace those damaged by exposure to the environment; olfactory sensory neurons generally survive for only 1-2 months.
سلولهای پشتیبان موکوس ترشح میکنند که محیط مولکولی و یونی مناسب را برای تشخیص بو در اپیتلیوم بویایی فراهم میکند. مولکولهای مولد بو (بویندهها) در مخاط حل میشوند و به گیرندههای بویایی روی مژکهای نورونهای حسی بویایی متصل میشوند. پروتئینهای اتصال دهنده بو در مخاط ممکن است انتشار مواد خوشبو کننده به و از گیرنده بو را تسهیل کنند. سلولهای بنیادی پایه تحت میتوز قرار میگیرند تا نورونهای حسی بویایی جدیدی تولید کنند تا در صورت نیاز جایگزین نورونهایی شوند که در اثر قرار گرفتن در معرض محیط آسیب دیدهاند. نورونهای حسی بویایی معمولاً تنها 1-2 ماه زنده میمانند.
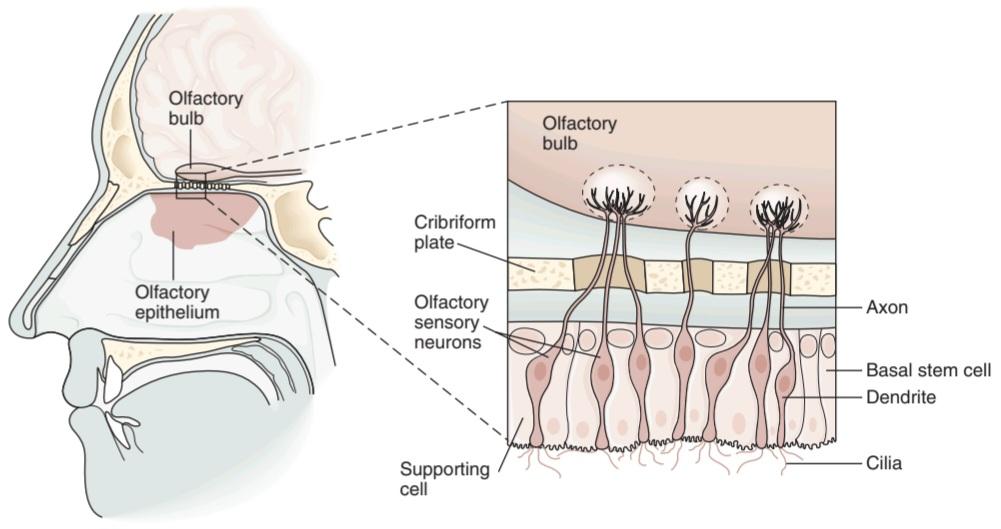
FIGURE 9-1 Structure of the olfactory epithelium. There are three cell types: olfactory sensory neurons, supporting (sustentacular) cells, and basal stem cells at the base of the epithelium. Each olfactory sensory neuron has a dendrite that projects to the epithelial surface. Numerous cilia protrude into the mucus layer lining the nasal lumen. Odorants bind to specific odorant receptors on the cilia and initiate a cascade of events leading to generation of action potentials in the sensory axon. Each olfactory sensory neuron has a single axon that projects to the olfactory bulb, a small ovoid structure that rests on the cribriform plate of the ethmoid bone. (Reproduced with permission from Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. New York, NY: McGraw-Hill; 2000.)
شکل 9-1 ساختار اپیتلیوم بویایی. سه نوع سلول وجود دارد: نورونهای حسی بویایی، سلولهای پشتیبان (sstentacular) و سلولهای بنیادی پایه در پایه اپیتلیوم. هر نورون حسی بویایی دارای یک دندریت است که به سطح اپیتلیال میرسد. مژکهای متعددی به داخل لایه مخاطی پوشاننده مجرای بینی بیرون زده اند. بوها به گیرندههای بویایی خاص روی مژکها متصل میشوند و مجموعه ای از رویدادها را آغاز میکنند که منجر به تولید پتانسیلهای عمل در آکسون حسی میشود. هر نورون حسی بویایی یک آکسون دارد که به پیاز بویایی میتابد، یک ساختار تخمیکوچک که بر روی صفحه کریبریفیک استخوان اتموئید قرار دارد. (تکثیر شده با اجازه از Kandel ER، Schwartz JH، Jessell TM [ویراستاران]: Principles of Neural Science، ویرایش چهارم نیویورک، نیویورک: McGraw-Hill؛ 2000.)
ODORANT RECEPTORS & SIGNAL TRANSDUCTION
The olfactory system can discriminate perhaps more than 1 million distinct odors due in part to the existence of many different functional odorant receptors. There are about 1000 olfactory genes in humans, accounting for 3% of the human genome; approximately 400 of these genes function as odorant receptors. The amino acid sequences of odorant receptors are very diverse, but all are G- protein-coupled receptors (GPCRs). When an odorant molecule binds to its receptor, the G-protein subunits (a, ẞ, y) dissociate (Figure 9-2). The a-subunit activates adenylyl cyclase to catalyze the production of cAMP which acts as a second messenger to open cation channels, increasing the membrane permeability to Na+, K, and Ca2+. The net effect is an inward-directed Ca2+current which produces the graded receptor potential. This then opens Ca2+- activated Cl channels, further depolarizing the cell due to the high intracellular Cl- levels in olfactory sensory neurons. If the stimulus is sufficient for the receptor potential to exceed its threshold, an action potential in the olfactory nerve (1st cranial nerve) is triggered.
گیرندههای بو و انتقال سیگنال
سیستم بویایی میتواند بیش از 1 میلیون بوی متمایز را تشخیص دهد که بخشی از آن به دلیل وجود گیرندههای مختلف بویایی است. حدود 1000 ژن بویایی در انسان وجود دارد که 3 درصد از ژنوم انسان را تشکیل میدهد. تقریباً 400 مورد از این ژنها به عنوان گیرندههای بو عمل میکنند. توالی اسیدهای آمینه گیرندههای بو بسیار متنوع است، اما همه گیرندههای جفت شده با پروتئین G (GPCRs) هستند. هنگامیکه یک مولکول خوشبو کننده به گیرنده خود متصل میشود، زیر واحدهای پروتئین G (a، ẞ، y) جدا میشوند (شکل 9-2). زیرواحد a آدنیلیل سیکلاز را فعال میکند تا تولید cAMP را کاتالیز کند که به عنوان پیام رسان دوم برای باز کردن کانالهای کاتیونی عمل میکند و نفوذپذیری غشاء را به Na+، K و +Ca2 افزایش میدهد. اثر خالص یک جریان +Ca2 به سمت داخل است که پتانسیل گیرنده درجه بندی شده را تولید میکند. سپس کانالهای کلر فعالشده +Ca2– را باز میکند و به دلیل سطح بالای کلر درون سلولی در نورونهای حسی بویایی، سلول را بیشتر دپلاریزه میکند. اگر محرک برای فراتر رفتن پتانسیل گیرنده از آستانه کافی باشد، پتانسیل عمل در عصب بویایی (اعصاب اول جمجمه ای) تحریک میشود.

FIGURE 9-2 Signal transduction in an odorant receptor. A) Olfactory receptors are examples of G-protein-coupled receptors; ie, they are associated with three G-protein subunits (a, ẞ, y). B) When an odorant binds to the receptors, the subunits dissociate. The a-subunit of G-proteins activates adenylyl cyclase to catalyze production of cAMP. cAMP acts as a second messenger to open cation channels. Inward diffusion of Na+ and Ca2+ produces depolarization.
شکل 9-2 انتقال سیگنال در یک گیرنده بو. الف) گیرندههای بویایی نمونههایی از گیرندههای جفت شده با پروتئین G هستند. به عنوان مثال، آنها با سه زیر واحد پروتئین G (a، ẞ، y) مرتبط هستند. ب) هنگامیکه یک ماده خوشبو به گیرندهها متصل میشود، زیر واحدها از هم جدا میشوند. زیرواحد a از پروتئینهای G، آدنیلیل سیکلاز را فعال میکند تا تولید cAMP را کاتالیز کند. cAMP به عنوان پیام رسان دوم برای باز کردن کانالهای کاتیونی عمل میکند. انتشار +Na و +Ca2 به داخل باعث دپلاریزاسیون میشود.
OLFACTORY SENSORY PATHWAY
In the olfactory bulb, the axons of the olfactory sensory neurons synapse on the primary dendrites of the mitral cells and tufted cells (Figure 9-3) to form olfactory glomeruli. Each olfactory sensory neuron expresses only one of the 400 functional olfactory genes, but each odorant can bind to a large pool of odorant receptors. Each olfactory sensory neuron projects to only one or two glomeruli. This provides a distinct two-dimensional map in the olfactory bulb that is unique to the odorant. The mitral cells with their glomeruli project to different parts of the olfactory cortex. The central olfactory system is able to decode the pattern of receptor-cell activity that signals the identity of the odorant.
مسیر حسی بویایی
در پیاز بویایی، آکسونهای نورونهای حسی بویایی روی دندریتهای اولیه سلولهای میترال و سلولهای توفت سیناپس میشوند (شکل 9-3) و گلومرولهای بویایی را تشکیل میدهند. هر نورون حسی بویایی تنها یکی از 400 ژن عملکردی بویایی را بیان میکند، اما هر بو میتواند به مجموعه بزرگی از گیرندههای بویایی متصل شود. هر نورون حسی بویایی تنها به یک یا دو گلومرول منعکس میشود. این یک نقشه دو بعدی متمایز در پیاز بویایی ارائه میدهد که منحصر به فرد بویایی است. سلولهای میترال با گلومرولهای خود به قسمتهای مختلف قشر بویایی میروند. سیستم بویایی مرکزی قادر به رمزگشایی الگوی فعالیت گیرنده سلولی است که هویت ماده بو را نشان میدهد.

FIGURE 9-3 Basic neural circuits in the olfactory bulb. Olfactory receptor cells with one type of odorant receptor project to one olfactory glomerulus (OG) and olfactory receptor cells with another type of receptor project to a different OG. Solid black arrows signify inhibition via GABA release, and white arrows signify excitatory connections via glutamate release. CP, cribriform plate; Gr, granule cell; M, mitral cell; PG, periglomerular cell; T, tufted cell. (Adapted with permission from Mori K, et al: The olfactory bulb: coding and processing of odor molecular information, Science 1999 Oct 22; 286(5440): 711–715.)
شکل 9-3 مدارهای عصبی اساسی در لامپ بویایی. سلولهای گیرنده بویایی با یک نوع گیرنده بویایی به یک گلومرول بویایی (OG) و سلولهای گیرنده بویایی با نوع دیگری از گیرنده به یک OG متفاوت پروژه میدهند. فلشهای سیاه جامد نشان دهنده مهار از طریق انتشار GABA و فلشهای سفید نشان دهنده اتصالات تحریکی از طریق انتشار گلوتامات است. CP، صفحه cribriform; Gr، سلول گرانول. M، سلول میترال؛ PG، سلول پری گلومرولی؛ T، سلول توفتی. (اقتباس شده با اجازه موری کی، و همکاران: لامپ بویایی: کدگذاری و پردازش اطلاعات مولکولی بو، Science 22 اکتبر 1999؛ 286(5440): 711-715.)
The olfactory bulbs also contain periglomerular cells, which are inhibitory neurons connecting one glomerulus to another, and granule cells, which have no axons and make reciprocal synapses with the lateral dendrites of the mitral and tufted cells. At these synapses, the mitral or tufted cells excite the granule cell by releasing glutamate, and the granule cells in turn inhibit the mitral or tufted cell by releasing GABA. The lateral inhibition mediated by periglomerular and granule cells functions to sharpen and focus olfactory signals.
پیازهای بویایی همچنین حاوی سلولهای اطراف گلومرولی هستند که نورونهای بازدارندهای هستند که یک گلومرول را به گلومرول دیگر متصل میکنند و سلولهای گرانولی که فاقد آکسون هستند و سیناپسهای متقابلی را با دندریتهای جانبی سلولهای میترال و توفت ایجاد میکنند. در این سیناپسها، سلولهای میترال یا توفتدار با آزاد کردن گلوتامات، سلول گرانول را تحریک میکنند و سلولهای گرانول نیز به نوبه خود با آزاد کردن GABA سلول میترال یا توفت را مهار میکنند. مهار جانبی که توسط سلولهای پری گلومرولی و گرانول انجام میشود برای تیز کردن و تمرکز سیگنالهای بویایی عمل میکند.
The axons of the mitral and tufted cells pass posteriorly through the lateral olfactory stria to terminate on apical dendrites of pyramidal cells in five regions of the olfactory cortex: anterior olfactory nucleus, olfactory tubercle, piriform cortex, amygdala, and entorhinal cortex (Figure 9-4). From these regions, information travels directly to the frontal cortex or via the thalamus to the orbitofrontal cortex. Conscious discrimination of odors is dependent on the pathway to the orbitofrontal cortex. The pathway to the amygdala mediates the emotional responses to olfactory stimuli, and the pathway to the entorhinal cortex is concerned with olfactory memories.
آکسونهای سلولهای میترال و تافتدار به صورت خلفی از طریق استریای بویایی جانبی عبور میکنند تا در دندریتهای رأسی سلولهای هرمی در پنج ناحیه از قشر بویایی خاتمه یابند: هسته بویایی قدامی، برآمدگی بویایی، قشر پیریفورم، آمیگدال و قشر انتورینال (شکل 9-4). از این نواحی، اطلاعات مستقیماً به قشر پیشانی یا از طریق تالاموس به قشر اوربیتوفرونتال منتقل میشوند. تشخیص آگاهانه بوها به مسیر قشر اوربیتوفرونتال بستگی دارد. مسیر منتهی به آمیگدال واسطه پاسخهای عاطفی به محرکهای بویایی است و مسیر منتهی به قشر انتورینال مربوط به خاطرات بویایی است.
In some mammals, the nasal cavity contains another patch of olfactory epithelium located along the nasal septum in the vomeronasal organ that is concerned with the perception of pheromones. Vomeronasal sensory neurons project to the accessory olfactory bulb (Figure 9-4) and from there to regions of the amygdala and hypothalamus that are concerned with reproduction and ingestive behavior. The vomeronasal organ has about 100 G-protein-coupled odorant receptors that differ in structure from those in the rest of the olfactory epithelium.
در برخی از پستانداران، حفره بینی حاوی لکه دیگری از اپیتلیوم بویایی است که در امتداد سپتوم بینی در اندام وومرونازال قرار دارد که با درک فرومونها مرتبط است. نورونهای حسی وومرونازال به حباب بویایی کمکی (شکل 9-4) و از آنجا به مناطقی از آمیگدال و هیپوتالاموس که مربوط به تولید مثل و رفتار هضمیهستند میروند. اندام وومرونازال حدود 100 گیرنده بویایی جفت شده با پروتئین G دارد که از نظر ساختار با سایر گیرندههای اپیتلیوم بویایی متفاوت است.
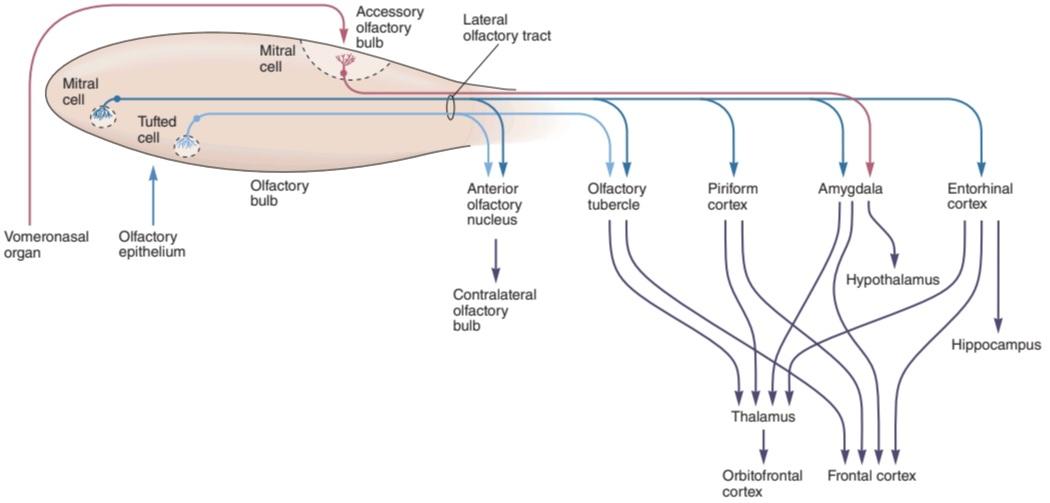
FIGURE 9-4 Diagram of the olfactory pathway. Information is transmitted from the olfactory bulb by axons of mitral and tufted relay neurons in the lateral olfactory tract. Mitral cells project to five regions of the olfactory cortex: anterior olfactory nucleus, olfactory tubercle, piriform cortex, and parts of the amygdala and entorhinal cortex. Tufted cells project to anterior olfactory nucleus and olfactory tubercle; mitral cells in the accessory olfactory bulb project only to the amygdala. Conscious discrimination of odor depends on the neocortex (orbitofrontal and frontal cortices). Emotive aspects of olfaction derive from limbic projections (amygdala and hypothalamus). (Reproduced with permission from Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. New York, NY: McGraw-Hill; 2000.)
شکل 9-4 نمودار مسیر بویایی. اطلاعات از پیاز بویایی توسط آکسونهای نورونهای رله میترال و توفت در مجرای بویایی جانبی منتقل میشود. سلولهای میترال به پنج ناحیه از قشر بویایی پرتاب میشوند: هسته بویایی قدامی، توبرکل بویایی، قشر پیریفورم، و بخشهایی از آمیگدال و قشر آنتورینال. سلولهای توفتی به سمت هسته بویایی قدامیو سل بویایی حرکت میکنند. سلولهای میترال در پیاز بویایی فرعی فقط به آمیگدال میرسند. تشخیص بو به نئوکورتکس (قشر اربیتو فرونتال و فرونتال) بستگی دارد. جنبههای احساسی بویایی از برجستگیهای لیمبیک (آمیگدال و هیپوتالاموس) ناشی میشود. (تکثیر شده با اجازه از Kandel ER، Schwartz JH، Jessell TM [ویراستاران]: Principles of Neural Science، ویرایش چهارم نیویورک، نیویورک: McGraw-Hill؛ 2000.)
ODOR DETECTION THRESHOLD
Odorants are generally small, containing between 3 and 20 carbon atoms; molecules with the same number of carbon atoms but different structural configurations have different odors. Relatively high water and lipid solubility is characteristic of substances with strong odors. Some common abnormalities in odor detection are described in Clinical Box 9-1.
آستانه تشخیص بو
بوها معمولاً کوچک هستند و بین 3 تا 20 اتم کربن دارند. مولکولهایی با تعداد اتمهای کربن یکسان، اما ساختارهای متفاوت، بوهای متفاوتی دارند. حلالیت نسبتاً بالا در آب و چربی مشخصه مواد با بوهای قوی است. برخی از ناهنجاریهای رایج در تشخیص بو در جعبه بالینی 9-1 توضیح داده شده است.
The odor detection threshold is the lowest concentration of a chemical that can be detected. Examples of substances detected at very low concentrations include hydrogen sulfide (0.0005 parts per million, ppm), acetic acid (0.016 ppm), kerosene (0.1 ppm), and gasoline (0.3 ppm). Some toxic substances are essentially odorless; they have odor detection thresholds higher than lethal concentrations. For example, carbon dioxide is detected at 74,000 ppm but is lethal at 50,000 ppm. The odor detection threshold for a given odorant is not the same in all individuals.
آستانه تشخیص بو کمترین غلظت یک ماده شیمیایی است که میتواند تشخیص داده شود. نمونههایی از مواد شناسایی شده در غلظتهای بسیار پایین عبارتند از: سولفید هیدروژن (0.0005 قسمت در میلیون، ppm)، اسید استیک (0.016 ppm)، نفت سفید (0.1 ppm) و بنزین (0.3 ppm). برخی از مواد سمیاساساً بی بو هستند. آنها آستانه تشخیص بو بالاتر از غلظت کشنده دارند. به عنوان مثال، دی اکسید کربن در ppm 74000 تشخیص داده میشود اما در ppm 50000 کشنده است. آستانه تشخیص بو برای یک ماده معطر در همه افراد یکسان نیست.
The sense of smell is said to be more acute in women than in men, and in women it is most acute at the time of ovulation. Although olfactory discrimination is remarkable, determination of differences in the intensity of any given odor is poor. The concentration of an odor-producing substance must be changed by about 30% before a difference can be detected. The comparable visual discrimination threshold is a 1% change in light intensity.
گفته میشود حس بویایی در زنان حادتر از مردان است و در زنان در زمان تخمک گذاری حادتر است. اگرچه تمایز بویایی قابل توجه است، تعیین تفاوت در شدت هر بو ضعیف است. قبل از اینکه بتوان تفاوت را تشخیص داد، غلظت یک ماده تولید کننده بو باید حدود 30 درصد تغییر کند. آستانه تمایز بصری قابل مقایسه 1٪ تغییر در شدت نور است.
CLINICAL BOX 9.1
Abnormalities in Odor Detection
Anosmia (inability to smell) and hyposmia or hypesthesia (diminished olfactory sensitivity) can result from simple nasal congestion, nasal polyps, or prolonged use of nasal decongestants. It may also be a sign of a more serious problem such as damage to the olfactory nerves due to fractures of the cribriform plate or head trauma, tumors (eg, neuroblastomas or meningiomas), and respiratory tract infections. Congenital anosmia is a rare disorder in which an individual is born without the ability to smell. Olfactory dysfunction is often one of the earliest clinical symptoms of Alzheimer disease. According to the National Institutes of Health, 1-2% of the North American population under the age of 65 experiences a significant degree of loss of smell. However, 50% of individuals between the ages of 65 and 80 and >75% of those over the age of 80 have an impaired ability to identify smells. Because of the close relationship between taste and smell, anosmia is associated with a reduction in taste sensitivity (hypogeusia). Anosmia is generally permanent in cases in which the olfactory nerve or other neural elements in the olfactory neural pathway are damaged. In addition to not being able to experience the enjoyment of pleasant aromas and a full spectrum of tastes, individuals with anosmia are at risk because they are not able to detect the odor from dangers such as gas leaks, fire, and spoiled food. Hyperosmia (enhanced olfactory sensitivity) is less common than loss of smell, but pregnant women commonly become oversensitive to smell. Dysosmia (distorted sense of smell) can be caused by several disorders including sinus infections, partial damage to the olfactory nerves, and poor dental hygiene. An aura of a disagreeable odor (eg, burning rubber) can occur when an individual experiences an uncinate seizure that originates in the medial temporal lobe.
جعبه بالینی 9.1
ناهنجاری در تشخیص بو
آنوسمی(ناتوانی در بویایی) و هیپوسمییا هیپستزی (کاهش حساسیت بویایی) میتواند ناشی از احتقان ساده بینی، پولیپ بینی یا استفاده طولانی مدت از داروهای ضد احتقان بینی باشد. همچنین ممکن است نشانه ای از یک مشکل جدی تر مانند آسیب به اعصاب بویایی به دلیل شکستگی صفحه کریبریفرم یا ضربه سر، تومورها (مانند نوروبلاستوما یا مننژیوم) و عفونتهای دستگاه تنفسی باشد. آنوسمیمادرزادی یک اختلال نادر است که در آن فرد بدون توانایی بویایی متولد میشود. اختلال بویایی اغلب یکی از اولین علائم بالینی بیماری آلزایمر است. به گفته مؤسسه ملی بهداشت، 1-2٪ از جمعیت آمریکای شمالی زیر 65 سال درجه قابل توجهی از کاهش بویایی را تجربه میکنند. با این حال، 50٪ از افراد بین 65 تا 80 سال و بیش از 75٪ از افراد بالای 80 سال توانایی تشخیص بوها را مختل میکنند. به دلیل رابطه نزدیک بین طعم و بوی، آنوسمیبا کاهش حساسیت چشایی (hypogeusia) همراه است. آنوسمیبه طور کلی در مواردی که عصب بویایی یا سایر عناصر عصبی در مسیر عصبی بویایی آسیب دیده باشد دائمیاست. افراد مبتلا به آنوسمیعلاوه بر اینکه نمیتوانند لذت رایحههای مطبوع و طیف کاملی از طعمها را تجربه کنند، در معرض خطر هستند، زیرا قادر به تشخیص بوی ناشی از خطراتی مانند نشت گاز، آتش سوزی و مواد غذایی فاسد نیستند. هیپراسمی(افزایش حساسیت بویایی) نسبت به از دست دادن بویایی کمتر شایع است، اما زنان باردار معمولاً بیش از حد به بویایی حساس میشوند. دیسوسمی(تحریف حس بویایی) میتواند ناشی از چندین اختلال از جمله عفونت سینوسی، آسیب جزئی به اعصاب بویایی و بهداشت نامناسب دندان باشد.هاله ای با بوی نامطلوب (مثلاً سوزاندن لاستیک) زمانی رخ میدهد که فرد یک تشنج غیرسینات را تجربه کند که منشأ آن از لوب تمپورال داخلی است.
THERAPEUTIQ HIGHLIGHTS
Quite often anosmia is a temporary condition due to sinus infection or a common cold, but it can be permanent if caused by nasal polyps or trauma. Antibiotics can be prescribed to reduce the inflammation caused by polyps and improve the ability to smell. In some cases, surgery is performed to remove the nasal polyps. Topical corticosteroids have also been shown to be effective in reversing the loss of smell due to nasal and sinus diseases.
نکات برجسته درمانی
اغلب آنوسمییک وضعیت موقتی به دلیل عفونت سینوسی یا سرماخوردگی است، اما اگر ناشی از پولیپ بینی یا ضربه باشد، میتواند دائمیباشد. آنتی بیوتیکها را میتوان برای کاهش التهاب ناشی از پولیپ و بهبود توانایی بویایی تجویز کرد. در برخی موارد، جراحی برای برداشتن پولیپ بینی انجام میشود. همچنین نشان داده شده است که کورتیکواستروئیدهای موضعی در معکوس کردن از دست دادن بویایی ناشی از بیماریهای بینی و سینوسی موثر هستند.
TASTE
TASTE BUDS
The specialized sense organ for taste (gustation) consists of about 5000 taste buds located primarily on the papillae of the dorsal surface of the tongue in humans (Figure 9-5). The fungiform papillae are rounded structures most numerous near the tip of the tongue; the circumvallate papillae are prominent structures arranged in a V on the back of the tongue; the foliate papillae are on the posterior edge of the tongue. Each fungiform papilla has up to five taste buds, mostly located at the top of the papilla, while each circumvallate and foliate papilla contain up to 100 taste buds, mostly located along the sides of the papillae. Taste buds are also located in the soft palate, epiglottis, and pharynx.
طعم و مزه
جوانههای چشایی
اندام حسی تخصصی برای چشایی (چشیدن) از حدود 5000 جوانه چشایی تشکیل شده است که عمدتاً بر روی پاپیلاهای سطح پشتی زبان در انسان قرار دارند (شکل 9-5). پاپیلاهای قارچی شکل ساختارهای گردی هستند که بیشتر در نزدیکی نوک زبان هستند. پاپیلاهای circumvallate ساختارهای برجسته ای هستند که به صورت V در پشت زبان مرتب شده اند. پاپیلاهای برگ در لبه خلفی زبان قرار دارند. هر پاپیلای قارچی تا پنج جوانه چشایی دارد که بیشتر در بالای پاپیلا قرار دارد، در حالی که هر پاپیلای دور و برگی دارای تا 100 جوانه چشایی است که بیشتر در امتداد طرفین پاپیلا قرار دارند. جوانههای چشایی نیز در کام نرم، اپی گلوت و حلق قرار دارند.
Each taste bud contains 50-100 taste receptor cells and numerous basal cells and support cells (Figure 9-5). The taste receptor cells are modified epithelial cells that respond to chemical stimuli or tastants. The apical ends of taste cells have microvilli that project into the taste pore, a small opening on the dorsal surface of the tongue where tastes cells are exposed to the oral contents. Saliva in the oral cavity acts as a solvent for tastants; after dissolving, the chemical diffuses to the taste receptor sites. Saliva may also function to cleanse the mouth to prepare the taste receptors for a new stimulant.
هر جوانه چشایی شامل 50-100 سلول گیرنده چشایی و سلولهای پایه و سلولهای پشتیبان متعدد است (شکل 9-5). سلولهای گیرنده چشایی سلولهای اپیتلیال اصلاح شده ای هستند که به محرکها یا مزههای شیمیایی پاسخ میدهند. انتهای آپیکال سلولهای چشایی دارای میکروویلی است که به داخل منافذ چشایی، یک روزنه کوچک در سطح پشتی زبان که در آن سلولهای چشایی در معرض محتویات دهان قرار میگیرند، بیرون میزند. بزاق در حفره دهان به عنوان یک حلال برای طعم دهندهها عمل میکند. پس از انحلال، این ماده شیمیایی به محلهای گیرنده چشایی پخش میشود. بزاق همچنین ممکن است برای پاکسازی دهان عمل کند تا گیرندههای چشایی برای یک محرک جدید آماده شود.
Each taste bud is innervated by about 50 nerve fibers, and conversely, each nerve fiber receives input from an average of five taste buds. The basal cells arise from the epithelial cells surrounding the taste bud. They differentiate into new taste cells as taste cells survive for only about 10 days. If the sensory nerve is cut, the taste buds it innervates degenerate and eventually disappear.
هر جوانه چشایی توسط حدود 50 رشته عصبی عصب دهی میشود و برعکس، هر فیبر عصبی به طور متوسط از پنج جوانه چشایی ورودی دریافت میکند. سلولهای پایه از سلولهای اپیتلیال اطراف جوانه چشایی به وجود میآیند. آنها به سلولهای چشایی جدید تمایز مییابند زیرا سلولهای چشایی تنها حدود 10 روز زنده میمانند. اگر عصب حسی قطع شود، جوانههای چشایی که آن را عصب دهی میکند، تحلیل رفته و در نهایت ناپدید میشود.

FIGURE 9-5 Taste buds located in papillae of the human tongue. A) Taste buds on the anterior two-thirds of the tongue are innervated by the chorda tympani branch of the facial nerve; those on the posterior one-third of the tongue are innervated by the lingual branch of the glossopharyngeal nerve. B) The three major types of papillae (circumvallate, foliate, and fungiform) are located on specific parts of the tongue. C) Taste buds are composed of basal stem cells and three types of taste cells (dark, light, and intermediate). Taste cells extend from the base of the taste bud to the taste pore, where microvilli contact tastants dissolved in saliva and mucus. (Modified with permission from Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. New York, NY: McGraw-Hill; 2000.)
شکل 9-5 جوانههای چشایی واقع در پاپیلاهای زبان انسان. الف) جوانههای چشایی در دو سوم قدامیزبان توسط شاخه chorda tympani عصب صورت عصب دهی میشوند. آنهایی که در یک سوم خلفی زبان قرار دارند توسط شاخه زبانی عصب گلوفارنژیال عصب دهی میشوند. ب) سه نوع اصلی پاپیلا (دور، شاخ و برگ و قارچ شکل) در قسمتهای خاصی از زبان قرار دارند. ج) جوانههای چشایی از سلولهای بنیادی پایه و سه نوع سلول چشایی (تیره، روشن و متوسط) تشکیل شده اند. سلولهای چشایی از پایه جوانه چشایی تا منافذ چشایی گسترش مییابند، جایی که میکروویلها با مزههای حل شده در بزاق و مخاط تماس میگیرند. (اصلاح شده با اجازه Kandel ER، Schwartz JH، Jessell TM [ویراستاران]: Principles of Neural Science، ویرایش چهارم نیویورک، نیویورک: McGraw-Hill؛ 2000.)
TASTE PATHWAYS
The sensory nerve fibers from the taste buds on the anterior two-thirds of the tongue travel in the chorda tympani branch of the facial nerve, and those from the posterior third of the tongue reach the brainstem via the glossopharyngeal nerve (Figure 9-6). The fibers from areas other than the tongue (eg, pharynx) reach the brain stem via the vagus nerve. On each side, the myelinated but relatively slowly conducting taste fibers in these three nerves unite in the gustatory portion of the nucleus of the tractus solitarius (NTS) in the medulla oblongata (Figure 9-6). From there, axons of second-order neurons ascend in the ipsilateral medial lemniscus and project directly to the ventral posteromedial nucleus of the thalamus. From the thalamus, the axons of the third-order neurons pass to neurons in the anterior insula and the frontal operculum in the ipsilateral cerebral cortex. This region is rostral to the face area of the postcentral gyrus, which is probably the area that mediates conscious perception of taste and taste discrimination.
مسیرهای طعم
رشتههای عصبی حسی از جوانههای چشایی در دو سوم قدامیزبان در شاخه chorda tympani عصب صورت حرکت میکنند و رشتههای عصبی از یک سوم خلفی زبان از طریق عصب گلوفارنکس به ساقه مغز میرسند (شکل 9-6). الیاف نواحی غیر از زبان (مثلاً حلق) از طریق عصب واگ به ساقه مغز میرسد. در هر طرف، الیاف چشایی میلین دار اما نسبتاً آهسته رسانا در این سه عصب در بخش چشایی هسته tractus solitarius (NTS) در بصل النخاع (شکل 9-6) متحد میشوند. از آنجا، آکسونهای نورونهای مرتبه دوم در لمنیسکوس داخلی همان طرف بالا میروند و مستقیماً به هسته خلفی شکمیتالاموس میروند. از تالاموس، آکسونهای نورونهای مرتبه سوم به نورونها در اینسولای قدامیو اپرکولوم پیشانی در قشر مغز همان طرف منتقل میشوند. این ناحیه منقاری به ناحیه صورت شکنج پست مرکزی است، که احتمالاً ناحیه ای است که واسطه درک آگاهانه طعم و مزه است.
Sensory fibers in the trigeminal (5th cranial) nerve also innervate the tongue and contribute to the burning sensation experienced when we eat foods containing capsaicin. Taste buds are surrounded by TRPV1 receptors on trigeminal nociceptive fibers that are activated in response to eating spicy foods.
فیبرهای حسی در عصب سه قلو (پنجم جمجمه) نیز زبان را عصب دهی میکنند و به احساس سوزش در هنگام خوردن غذاهای حاوی کپسایسین کمک میکنند. جوانههای چشایی توسط گیرندههای TRPV1 بر روی فیبرهای درد سه قلو که در پاسخ به خوردن غذاهای تند فعال میشوند، احاطه شده اند.

FIGURE 9-6 Diagram of taste pathways. Signals from the taste buds travel via different nerves to gustatory areas of the nucleus of the tractus solitarius, which relays information to the thalamus; the thalamus projects to the gustatory cortex. (Modified with permission from Kandel ER, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. New York, NY: McGraw-Hill; 2000.)
شکل 9-6 نمودار مسیرهای چشایی. سیگنالهای جوانههای چشایی از طریق اعصاب مختلف به نواحی چشایی هسته tractus solitarius میرسند، که اطلاعات را به تالاموس منتقل میکند. تالاموس به سمت قشر چشایی پیش میرود. (اصلاح شده با اجازه Kandel ER، Schwartz JH، Jessell TM [ویراستاران]: Principles of Neural Science، ویرایش چهارم نیویورک، نیویورک: McGraw-Hill؛ 2000.)
TASTE MODALITIES, RECEPTORS, & TRANSDUCTION
Humans have five basic taste modalities: salt, sweet, sour, bitter, and umami. Common stimuli for these sensory modalities are sodium chloride, sucrose, hydrochloric acid, quinine, and monosodium glutamate, respectively. All tastants are sensed from all parts of the tongue and adjacent structures. Afferent nerves to the NTS contain fibers from all types of taste receptors, without any clear localization of types. An individual taste receptor cell may respond to more than one type of tastant. The central nervous system can distinguish the various tastes from one another because each type of taste receptor cell connects to a particular gustatory axon.
روشهای طعم، گیرندهها، و انتقال
انسانها پنج نوع طعم اساسی دارند: نمک، شیرین، ترش، تلخ و اومامی. محرکهای رایج برای این روشهای حسی به ترتیب کلرید سدیم، ساکارز، اسید هیدروکلریک، کینین و مونوسدیم گلوتامات هستند. تمام مزهها از تمام قسمتهای زبان و ساختارهای مجاور حس میشود. اعصاب آوران به NTS حاوی فیبرهایی از انواع گیرندههای چشایی هستند، بدون هیچ گونه محلی سازی واضحی از انواع. یک سلول گیرنده چشایی فردی ممکن است به بیش از یک نوع طعم پاسخ دهد. سیستم عصبی مرکزی میتواند طعمهای مختلف را از یکدیگر تشخیص دهد زیرا هر نوع سلول گیرنده چشایی به آکسون چشایی خاصی متصل میشود.
Figure 9-7 shows the putative receptors for the five modalities of taste. They include the two major types of receptors: ligand-gated channels (ionotropic receptors) and GPCRs (metabotropic receptors). Salt and sour tastes are triggered by activation of ionotropic receptors; sour, bitter, and umami tastes are triggered by activation of metabotropic receptors. Many GPCRS in the human genome are taste receptors (TIRS, T2Rs families).
شکل 9-7 گیرندههای احتمالی را برای پنج روش چشایی نشان میدهد. آنها شامل دو نوع اصلی گیرنده میشوند: کانالهای دروازهدار لیگاند (گیرندههای یونوتروپیک) و GPCRs (گیرندههای متابوتروپیک). طعم نمک و ترش با فعال شدن گیرندههای یونوتروپیک تحریک میشود. مزههای ترش، تلخ و umami با فعال شدن گیرندههای متابوتروپیک تحریک میشوند. بسیاری از GPCRS در ژنوم انسان گیرندههای چشایی (خانوادههای TIRS، T2Rs) هستند.

FIGURE 9-7 Signal transduction in taste receptors. Salt and sour tastes are mediated via the epithelial sodium channel (ENaC). This receptor has two subunits (a and y), each crossing the membrane twice, resulting in intracellular N and C termini (NT, CT). Salt is sensed following Na* movement; sour is mediated by movement of H+. Sweet, bitter, and umami tastes are sensed via G- protein-coupled receptors that span the membrane seven times and have varying lengths of CT and NT (represented as ribbon structures). Sweet tastes are detected by the T1R2 and T1R3 families; bitter and umami tastes are detected by the T2R family and mGluR4, respectively.
شکل 9-7 انتقال سیگنال در گیرندههای چشایی. طعم نمک و ترش از طریق کانال سدیم اپیتلیال (ENaC) ایجاد میشود. این گیرنده دارای دو زیر واحد (a و y) است که هر کدام دو بار از غشاء عبور میکنند و در نتیجه N و C انتهایی داخل سلولی (NT، CT) ایجاد میشود. نمک به دنبال حرکت +Na حس میشود. ترش با حرکت +H واسطه میشود. مزههای شیرین، تلخ و اومامیاز طریق گیرندههای جفت شده با پروتئین G که هفت بار غشاء را پوشش میدهند و طولهای متفاوتی از CT و NT دارند (که به صورت ساختارهای نواری نشان داده میشوند) حس میشوند. طعم شیرین توسط خانوادههای T1R2 و T1R3 تشخیص داده میشود. طعم تلخ و umami به ترتیب توسط خانواده T2R و mGluR4 شناسایی میشوند.
Salt-sensitive taste is mediated by an epithelial sodium channel (ENaC). The entry of Na+ into the salt receptor depolarizes the membrane, generating a receptor potential. The sour taste is triggered by protons (H+ ions). ENaCs permit the entry of protons and may contribute to the sensation of sour taste. The H+ ions can also bind to and block a K+-sensitive channel. The fall in K+ permeability can depolarize the membrane. Also, a hyperpolarization-activated cyclic nucleotide-gated cation channel (HCN) and other mechanisms may contribute to sour transduction.
طعم حساس به نمک توسط یک کانال سدیم اپیتلیال (ENaC) واسطه میشود. ورود +Na به گیرنده نمک غشاء را دپولاریزه میکند و پتانسیل گیرنده ایجاد میکند. طعم ترش توسط پروتونها (یونهای +H) تحریک میشود. ENaCها اجازه ورود پروتونها را میدهند و ممکن است به احساس طعم ترش کمک کنند. یونهای +H همچنین میتوانند به یک کانال حساس به +K متصل شده و آن را مسدود کنند. کاهش نفوذپذیری +K میتواند غشا را دپلاریزه کند. همچنین، یک کانال کاتیونی حلقوی دردار با نوکلئوتید فعال شده با هیپرپلاریزاسیون (HCN) و مکانیسمهای دیگر ممکن است به انتقال ترش کمک کنند.
Substances that taste sweet are detected by at least two types of GPCRs, T1R2 and T1R3. Sugars taste sweet, but so do compounds such as saccharin that have an entirely different structure. Natural sugars such as sucrose and synthetic sweeteners may act on gustducin via different receptors. Sweet-responsive receptors act via cyclic nucleotides and inositol phosphate metabolism.
موادی که طعم شیرین دارند حداقل توسط دو نوع GPCR، T1R2 و T1R3 شناسایی میشوند. قندها طعم شیرین دارند، اما ترکیباتی مانند ساخارین که ساختار کاملاً متفاوتی دارند نیز طعم شیرینی دارند. قندهای طبیعی مانند ساکارز و شیرین کنندههای مصنوعی ممکن است از طریق گیرندههای مختلف بر روی گاستدوسین اثر بگذارند. گیرندههای پاسخگو به شیرینی از طریق نوکلئوتیدهای حلقوی و متابولیسم اینوزیتول فسفات عمل میکنند.
Bitter taste is produced by a variety of unrelated compounds. Many of these are poisons, and bitterness serves as a warning to avoid them. Some bitter compounds (eg, quinine) are membrane permeable and bind to and block K+- selective channels. Many bitter tastants (eg, strychnine) bind to GPCRs (T2R family) that couple to the heterotrimeric G-protein, gustducin. Gustducin lowers cAMP and increases the formation of inositol phosphates (IP3), which releases Ca Ca2+ to trigger depolarization.
طعم تلخ توسط انواع ترکیبات نامرتبط ایجاد میشود. بسیاری از اینها سم هستند و تلخی به عنوان هشداری برای اجتناب از آنها عمل میکند. برخی از ترکیبات تلخ (مانند کینین) نفوذپذیر به غشاء هستند و به کانالهای انتخابی +K- متصل میشوند و آنها را مسدود میکنند. بسیاری از طعمهای تلخ (مثلاً استریکنین) به GPCRs (خانواده T2R) متصل میشوند که به پروتئین G هتروتریمریک، گوستدوسین متصل میشوند. گاستدوسین cAMP را کاهش میدهد و تشکیل فسفات اینوزیتول (IP3) را افزایش میدهد، که کلسیم +Ca2 را آزاد میکند تا باعث دپلاریزاسیون شود.
Umami tastants activate a receptor comprised of T1R1 and T1R3. Umami taste may also involve the activation of a truncated metabotropic glutamate receptor, mGluR4, in the taste buds.
مزههای Umami گیرنده ای متشکل از T1R1 و T1R3 را فعال میکنند. طعم Umami همچنین ممکن است شامل فعال شدن یک گیرنده متابوتروپیک کوتاه شده گلوتامات، mGluR4، در جوانههای چشایی باشد.
TASTE THRESHOLDS & INTENSITY DISCRIMINATION
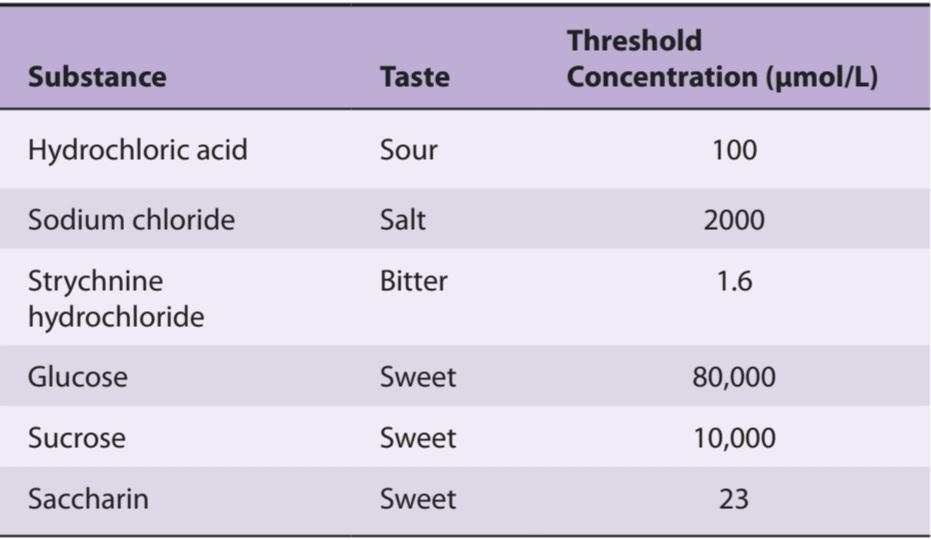
The ability of humans to discriminate differences in the intensity of tastes is relatively crude. A 30% change in the concentration of the tastant is necessary before a difference can be detected. Taste threshold refers to the minimum concentration at which a substance can be perceived. Table 9-1 shows the threshold concentration of various substances needed for a taste bud to respond. Bitter substances tend to have the lowest threshold. Some toxic substances such as strychnine have a bitter taste at very low concentrations, preventing accidental ingestion of this chemical, which causes fatal convulsions. Some common abnormalities in taste detection are described in Clinical Box 9-2.
آستانه طعم و تبعیض شدت
توانایی انسان در تشخیص تفاوت در شدت ذائقه نسبتاً خام است. قبل از اینکه بتوان تفاوت را تشخیص داد، تغییر 30 درصدی در غلظت ماده ضروری است. آستانه طعم اشاره به حداقل غلظتی دارد که در آن یک ماده را میتوان درک کرد. جدول 9-1 غلظت آستانه مواد مختلف مورد نیاز برای واکنش جوانه چشایی را نشان میدهد. مواد تلخ کمترین آستانه را دارند. برخی از مواد سمیمانند استریکنین در غلظتهای بسیار کم طعم تلخی دارند و از بلع تصادفی این ماده شیمیایی که باعث تشنج کشنده میشود جلوگیری میکند. برخی از ناهنجاریهای رایج در تشخیص طعم در جعبه بالینی 9-2 توضیح داده شده است.
TABLE 9-1 Some taste thresholds.
جدول 9-1 برخی از آستانههای طعم.
CLINICAL BOX 9.2
Abnormalities in Taste Detection
Ageusia (absence of the sense of taste) and hypogeusia (diminished taste sensitivity) can be caused by damage to the lingual or glossopharyngeal nerve. Neurological disorders such as vestibular schwannoma, Bell palsy, familial dysautonomia, multiple sclerosis, certain infections (eg, primary ameboid meningoencephalopathy), and poor oral hygiene can also cause problems with taste sensitivity. Ageusia can be an adverse side effect of various drugs, including cisplatin and captopril, or vitamin B3 or zinc deficiencies. Aging and tobacco abuse also contribute to diminished taste. Dysgeusia or parageusia (unpleasant perception of taste) causes a metallic, salty, foul, or rancid taste. In many cases, dysgeusia is a temporary problem. Factors contributing to ageusia or hypogeusia can also lead to abnormal taste sensitivity. Taste disturbances can also occur under conditions in which serotonin (5-HT) and norepinephrine (NE) levels are altered (eg, during anxiety or depression). This implies that these neuromodulators contribute to the determination of taste thresholds. Administration of a 5-HT reuptake inhibitor reduces sensitivity to sucrose (sweet taste) and quinine (bitter taste). In contrast, administration of an NE reuptake inhibitor reduces bitter taste and sour thresholds. About 25% of the population has a heightened sensitivity to taste, in particular to bitterness. These individuals are called supertasters; this may be due to the presence of an increased number of fungiform papillae on their tongue.
جعبه بالینی 9.2
ناهنجاری در تشخیص طعم
آئوژیوزیا (فقدان حس چشایی) و هیپوژئوزیا (کاهش حساسیت چشایی) میتواند در اثر آسیب به عصب زبانی یا گلوفارنکس ایجاد شود. اختلالات عصبی مانند شوانوم دهلیزی، فلج بل، دیساآتونومیخانوادگی، مولتیپل اسکلروزیس، عفونتهای خاص (مثلاً مننگوآنسفالوپاتی آمبوئید اولیه) و بهداشت نامناسب دهان نیز میتوانند باعث ایجاد مشکلاتی در حساسیت چشایی شوند. آئوزیا میتواند از عوارض جانبی داروهای مختلف از جمله سیس پلاتین و کاپتوپریل یا کمبود ویتامین B3 یا روی باشد. افزایش سن و سوء مصرف تنباکو نیز به کاهش طعم کمک میکند. دیسگوزیا یا پاراژئوزیا (درک ناخوشایند طعم) باعث ایجاد طعم فلزی، شور، بد یا ترش میشود. در بسیاری از موارد، دیسژئوزی یک مشکل موقتی است. عوامل موثر در پیری یا هیپوژوزی نیز میتواند منجر به حساسیت چشایی غیرطبیعی شود. اختلالات چشایی همچنین میتواند تحت شرایطی رخ دهد که در آن سطوح سروتونین (5-HT) و نوراپی نفرین (NE) تغییر میکند (به عنوان مثال، در هنگام اضطراب یا افسردگی). این نشان میدهد که این نورومدولاتورها در تعیین آستانه طعم کمک میکنند. تجویز یک مهارکننده بازجذب 5-HT حساسیت به ساکارز (طعم شیرین) و کینین (طعم تلخ) را کاهش میدهد. در مقابل، تجویز یک مهارکننده بازجذب NE طعم تلخ و آستانه ترش را کاهش میدهد. حدود 25 درصد از جمعیت نسبت به طعم، به ویژه تلخی، حساسیت بیشتری دارند. به این افراد فوق ذوق میگویند. این ممکن است به دلیل وجود افزایش تعداد پاپیلاهای قارچی شکل روی زبان آنها باشد.
THERAPEUTIQ HIGHLIGHTS
Improved oral hygiene and adding zinc supplements to one’s diet can correct the inability to taste in some individuals.
نکات برجسته درمانی
بهبود بهداشت دهان و دندان و افزودن مکملهای روی به رژیم غذایی میتواند ناتوانی در چشایی را در برخی افراد اصلاح کند.
CHAPTER SUMMARY
خلاصه فصل
• The olfactory epithelium in the upper portion of the nasal cavity contains three types of cells involved in olfaction: olfactory sensory neurons that are responsible for olfactory transduction, supporting cells that secrete the mucus that provides the appropriate molecular and ionic environment for odor detection, and basal stem cells that generate new olfactory sensory neurons to replace those damaged by exposure to the environment.
• اپیتلیوم بویایی در قسمت فوقانی حفره بینی شامل سه نوع سلول درگیر در بویایی است: نورونهای حسی بویایی که مسئول انتقال بویایی هستند، سلولهای حمایت کننده که مخاطی را ترشح میکنند که محیط مولکولی و یونی مناسب را برای تشخیص بو فراهم میکند، و سلولهای بنیادی بنیادی که توسط سلولهای بنیادی آسیب دیده حسی جدید توسط سلولهای نورون فاکتور ایجاد میکنند.
• Processing of olfactory information occurs in the olfactory bulb where the axons of olfactory sensory neurons synapse on mitral cells and tufted cells to form olfactory glomeruli. The olfactory bulb also contains inhibitory periglomerular cells and granule cells which make reciprocal synapses with mitral and tufted cells.
• پردازش اطلاعات بویایی در پیاز بویایی رخ میدهد که در آن آکسونهای نورونهای حسی بویایی روی سلولهای میترال و سلولهای توفت سیناپس میشوند و گلومرولهای بویایی را تشکیل میدهند. پیاز بویایی همچنین حاوی سلولهای پری گلومرولی و سلولهای گرانولی است که سیناپسهای متقابلی را با سلولهای میترال و توفت ایجاد میکنند.
• The olfactory system can discriminate perhaps more than 1 million distinct odors due in part to the existence of 400 functional odorant genes (receptors). Each olfactory sensory neuron expresses only one of the 400 functional olfactory genes, but each odorant can bind to a large pool of odorant receptors.
• سیستم بویایی میتواند بیش از 1 میلیون بو متمایز را تشخیص دهد که بخشی از آن به دلیل وجود 400 ژن (گیرنده) است. هر نورون حسی بویایی تنها یکی از 400 ژن عملکردی بویایی را بیان میکند، اما هر بو میتواند به مجموعه بزرگی از گیرندههای بویایی متصل شود.
• Odorant receptors are part of a large family of GPCRs. When an odorant binds to a receptor, the G-protein subunits dissociate and the a-subunit activates adenylyl cyclase to increase production of cAMP which opens cation channels to increase membrane permeability to Na*, K, and Ca2+. Cl channels then open to further depolarize olfactory sensory neurons.
• گیرندههای بو بخشی از خانواده بزرگ GPCRها هستند. هنگامیکه یک ماده خوشبو به یک گیرنده متصل میشود، زیر واحدهای پروتئین G جدا میشوند و زیرواحد a آدنیلیل سیکلاز را فعال میکند تا تولید cAMP را افزایش دهد که کانالهای کاتیونی را برای افزایش نفوذپذیری غشاء به Na+، K و +Ca2 باز میکند. سپس کانالهای کلر برای دپلاریزاسیون بیشتر نورونهای حسی بویایی باز میشوند.
• Projections from mitral and tufted cells travel via the lateral olfactory stria directly to five regions of the olfactory cortex: anterior olfactory nucleus, olfactory tubercle, piriform cortex, amygdala, and entorhinal cortex.
• برجستگیهای سلولهای میترال و توفت از طریق استریای بویایی جانبی به طور مستقیم به پنج ناحیه قشر بویایی میروند: هسته بویایی قدامی، سل بویایی، قشر پیریفورم، آمیگدال و قشر آنتورینال.
• Taste buds are the specialized sense organs for taste and are composed of epithelial taste cells and basal stem cells. Taste buds are located primarily in the mucosa of the walls of papillae of the tongue, but also in the epiglottis, palate, and pharynx.
• جوانههای چشایی اندامهای حسی تخصصی برای چشایی هستند و از سلولهای چشایی اپیتلیال و سلولهای بنیادی پایه تشکیل شده اند. جوانههای چشایی عمدتاً در مخاط دیوارههای پاپیلای زبان و همچنین در اپی گلوت، کام و حلق قرار دارند.
• The five major taste modalities are salt, sour, bitter, sweet, and umami. Signal transduction mechanisms include passage through ion channels (ENaC for salt and sour), binding to and blocking ion channels (sour), and binding to GPCRs requiring second messenger systems (T1R2, T1R3 for sweet; T2R for bitter; and mGluR4 for umami).
• پنج نوع طعم اصلی عبارتند از: نمک، ترش، تلخ، شیرین و اومامی. مکانیسمهای انتقال سیگنال شامل عبور از کانالهای یونی (ENaC برای نمک و ترش)، اتصال به کانالهای یونی و مسدود کردن آنها (ترش)، و اتصال به GPCRهایی است که به سیستمهای پیامرسان دوم نیاز دارند (T1R2، T1R3 برای شیرین، T2R برای تلخ، و mGluR4 برای umami).
• The afferents from taste buds in the anterior two-thirds of the tongue travel via the facial nerve, those from the posterior one-third of the tongue travel via the glossopharyngeal nerve, and those located elsewhere travel via the vagus nerve. All of the sensory fibers from taste buds synapse in the NTS. From there, axons ascend via the ipsilateral medial lemniscus to the ventral posteromedial nucleus of the thalamus, and onto the anterior insula and frontal operculum in the ipsilateral cerebral cortex.
• آورانهای جوانههای چشایی در دو سوم قدامیزبان از طریق عصب صورت، آورانهای یک سوم خلفی زبان از طریق عصب گلوسوفارنکس و آنهایی که در جاهای دیگر قرار دارند از طریق عصب واگ حرکت میکنند. تمام فیبرهای حسی جوانههای چشایی در NTS سیناپس میشوند. از آنجا، آکسونها از طریق لمنیسکوس داخلی همان طرف به هسته خلفی شکمیتالاموس و به سمت اینسولای قدامیو اپرکولوم پیشانی در قشر مغز همان طرف صعود میکنند.
• Disorders of olfaction include anosmia (inability to smell), hyposmia (diminished olfactory sensitivity), hyperosmia (enhanced olfactory sensitivity), and dysosmia (distorted sense of smell). Causes include damage to the olfactory nerve, tumors, respiratory tract infections, and poor dental hygiene. Disorders of taste include ageusia (absence of the sense of taste), hypogeusia (diminished taste sensitivity), and dysgeusia (unpleasant perception of taste). Causes include damage to the facial or glossopharyngeal nerve, neurological disorders, drugs, vitamin deficiencies, and poor oral hygiene.
• اختلالات بویایی شامل آنوسمی(ناتوانی در بویایی)، هیپوسمی(کاهش حساسیت بویایی)، هیپراسمی(افزایش حساسیت بویایی) و دیسوسمی(تحریف حس بویایی) است. علل عبارتند از آسیب به عصب بویایی، تومورها، عفونتهای دستگاه تنفسی و بهداشت نامناسب دندان. اختلالات چشایی شامل پیری (فقدان حس چشایی)، هیپوژئوزی (کاهش حساسیت چشایی) و دیسژئوزی (درک ناخوشایند چشایی) است. علل شامل آسیب به صورت یا عصب گلوفارنکس، اختلالات عصبی، داروها، کمبود ویتامین و بهداشت نامناسب دهان.
MULTIPLE-CHOICE QUESTIONS
For all questions, select the single best answer unless otherwise directed.
سوالات چند گزینه ای
برای همه سؤالات، بهترین پاسخ را انتخاب کنید، مگر اینکه دستور دیگری داده شود.
1. A young boy was diagnosed with congenital anosmia, a rare disorder in which an individual is born without the ability to smell. Which parts of the nervous system might be defective in an individual with congenital anosmia to account for the inability to detect odors?
A. Glossopharyngeal nerve, olfactory bulb, ventral posterior medial nucleus of the thalamus, and anterior insula-frontal operculum
B. Olfactory sensory neurons, olfactory glomeruli, nucleus of the tractus solitarius, and ventral posterior lateral nucleus of the thalamus
C. Olfactory nerve, olfactory bulb, medial olfactory tract, and anterior insula- frontal operculum
D. Olfactory sensory neuron, 1st cranial nerve, olfactory glomeruli, and frontal cortex
E. Trigeminal nerve, olfactory glomeruli, lateral olfactory tubercle, and entorhinal cortex
1. یک پسر جوان به آنوسمیمادرزادی تشخیص داده شد، یک اختلال نادر که در آن فرد بدون توانایی بویایی متولد میشود. کدام بخش از سیستم عصبی ممکن است در یک فرد مبتلا به آنوسمیمادرزادی معیوب باشد تا علت ناتوانی در تشخیص بو باشد؟
الف. عصب گلوفارنکس، پیاز بویایی، هسته داخلی خلفی شکمیتالاموس، و اپرکولوم قدامیاینسولا- فرونتال
ب. نورونهای حسی بویایی، گلومرولهای بویایی، هسته تراکتوس سولیتریوس و هسته خلفی شکمیجانبی تالاموس
ج. عصب بویایی، پیاز بویایی، دستگاه بویایی داخلی و اپرکولوم قدامی- فرونتال
د. نورون حسی بویایی، عصب اول جمجمه، گلومرول بویایی و قشر فرونتال
ی. عصب سه قلو، گلومرول بویایی، سل بویایی جانبی و قشر آنتورینال
2. While working in a laboratory studying the olfactory system, a medical student was intrigued by the fact that a simple sense organ like the human olfactory epithelium can discriminate perhaps more than 1 million distinct odors. What factors may contribute to this phenomenon?
A. There are 500 types of odorant receptors and over 1000 types of odorant- binding proteins that sequester odorants to enhance sensory discrimination.
B. Each olfactory sensory neuron expresses a single odorant receptor gene and projects to a particular subset of mitral cells that connect to distinct parts of the olfactory cortex.
C. Odorants bind to a mixture of GPCR and ion channel receptors on olfactory sensory neurons and the axons of these sensory neurons form anatomically discrete synaptic units called olfactory glomeruli.
D. Lateral inhibition within olfactory glomeruli sharpen and focus olfactory signals and granule cells within the olfactory glomerulus make specific projections to the postcentral gyrus in the somatosensory cortex.
E. There are about 5000 types of odorant receptors and each odorant binds to only one of these.
2. هنگامیکه در آزمایشگاهی مشغول مطالعه سیستم بویایی بود، یک دانشجوی پزشکی مجذوب این واقعیت شد که یک اندام حسی ساده مانند اپیتلیوم بویایی انسان میتواند بیش از 1 میلیون بو متمایز را تشخیص دهد. چه عواملی ممکن است در ایجاد این پدیده موثر باشد؟
الف. 500 نوع گیرنده بو و بیش از 1000 نوع پروتئین متصل شونده به بو وجود دارد که بوها را جدا میکند تا تمایز حسی را افزایش دهد.
ب. هر نورون حسی بویایی یک ژن گیرنده بویایی منفرد را بیان میکند و به زیر مجموعه خاصی از سلولهای میترال که به قسمتهای مشخصی از قشر بویایی متصل میشوند، حرکت میکند.
ج. مواد بويايي به مخلوطي از گيرندههاي GPCR و كانال يوني روي نورونهاي حسي بويايي متصل ميشوند و آكسونهاي اين نورونهاي حسي واحدهاي سيناپسي مجزاي آناتوميك به نام گلومرولهاي بويايي را تشكيل ميدهند.
د. مهار جانبی در گلومرول بویایی، سیگنالهای بویایی را تیز و متمرکز میکند و سلولهای گرانول درون گلومرول بویایی، برجستگیهای خاصی را به شکنج پس مرکزی در قشر حسی تنی ایجاد میکنند.
ی. حدود 5000 نوع گیرنده بو وجود دارد و هر ماده بویایی تنها به یکی از آنها متصل میشود.
3. As part of a research experience, a medical student was reviewing reports on the effects of exposure to various neurotoxins on odor detection in humans. Which cells in the olfactory system are responsible for the ability to retain the sense of smell despite the fact that toxins can damage elements of the nasal mucosa?
A. Basal cells in the olfactory bulb undergo mitosis to generate new olfactory sensory neurons.
B. Surviving olfactory sensory neurons undergo neuroplasticity and make connections with the mitral and tufted cells that were originally connected to the destroyed sensory neurons.
C. Supporting cells in the olfactory epithelium release neurotrophic factors that stimulate the genesis of new olfactory sensory neurons.
D. Basal cells in the olfactory epithelium regenerate the neurons comprising the 1st cranial nerve.
E. Olfactory sensory neurons in the olfactory bulb are repaired because the surrounding environment contains neurotrophic factors.
3. به عنوان بخشی از یک تجربه تحقیقاتی، یک دانشجوی پزشکی در حال بررسی گزارشهایی در مورد اثرات قرار گرفتن در معرض نوروتوکسینهای مختلف بر تشخیص بو در انسان بود. با وجود اینکه سموم میتوانند به عناصر مخاط بینی آسیب برسانند، کدام سلولها در سیستم بویایی مسئول توانایی حفظ حس بویایی هستند؟
الف. سلولهای پایه در پیاز بویایی برای تولید نورونهای حسی بویایی جدید تحت میتوز قرار میگیرند.
ب. سلولهای عصبی حسی بویایی زنده مانده دچار نوروپلاستیسیته میشوند و با سلولهای میترال و توفتی که در ابتدا به نورونهای حسی تخریب شده متصل بودند، ارتباط برقرار میکنند.
ج. سلولهای پشتیبان در اپیتلیوم بویایی عوامل نوروتروفیک را آزاد میکنند که باعث تحریک پیدایش نورونهای حسی بویایی جدید میشود.
د. سلولهای پایه در اپیتلیوم بویایی نورونهای تشکیل دهنده عصب جمجمه 1 را بازسازی میکنند.
ی. نورونهای حسی بویایی در پیاز بویایی ترمیم میشوند زیرا محیط اطراف حاوی عوامل نوروتروفیک است.
4. A 9-year-old boy had frequent episodes of uncontrollable nose bleeds. At the advice of his clinician, he underwent surgery to correct a problem in his nasal septum. A few days after the surgery, he told his mother he could not smell the cinnamon rolls she was baking in the oven. Which of the following sequence of events occurs when an odorant binds to an odorant receptor?
A. Binding of the odorant to a ligand-gated ion channel promotes the influx of Na+ ions in the olfactory sensory neuron which leads to the induction of an action potential in the olfactory nerve.
B. Binding of the odorant promotes dissociation of G-protein subunits and the y-subunit activates adenylyl cyclase to increase cGMP which then selectively opens Na+ channels on the nerve membrane to induce an action potential in the olfactory nerve.
C. When the odorant binds to a combination of ligand-gated ion channels and GPCRs, there is an influx of Ca2+ that triggers an influx of Na* to induce an action potential in the olfactory sensory neuron.
D. Binding of the odorant to a GPCR on the cilia of the olfactory sensory neurons causes dissociation of the G-protein subunits, the a-subunit activates adenylyl cyclase to increase cAMP which opens cation channels, leading to increased permeability to Na, K, and Ca2+ on the nerve membrane and depolarization of the olfactory nerve.
4. یک پسر 9 ساله به دفعات خونریزیهای غیرقابل کنترل بینی داشت. او به توصیه پزشک معالجش برای رفع مشکل تیغه بینی تحت عمل جراحی قرار گرفت. چند روز پس از عمل جراحی، او به مادرش گفت که بوی رول دارچینی را که او در فر میپخت، حس نمیکند. کدام یک از توالی رویدادهای زیر زمانی رخ میدهد که یک ماده خوشبو کننده به گیرنده بویایی متصل شود؟
الف. اتصال ماده خوشبو به یک کانال یونی دارای لیگاند باعث هجوم یونهای Na+ در نورون حس بویایی میشود که منجر به القای پتانسیل عمل در عصب بویایی میشود.
ب. اتصال ماده خوشبو باعث تجزیه زیر واحدهای پروتئین G میشود و زیرواحد y آدنیلیل سیکلاز را فعال میکند تا cGMP را افزایش دهد که سپس به طور انتخابی کانالهای Na+ را روی غشای عصبی باز میکند تا پتانسیل عمل در عصب بویایی را القا کند.
ج. هنگامیکه ماده خوشبو به ترکیبی از کانالهای یونی دارای لیگاند و GPCR متصل میشود، هجوم +Ca2 وجود دارد که هجوم +Na را برای القای پتانسیل عمل در نورون حسی بویایی ایجاد میکند.
د. اتصال ماده خوشبو به یک GPCR روی مژکهای نورونهای حسی بویایی باعث جدا شدن زیرواحدهای پروتئین G میشود، زیرواحد a آدنیلیل سیکلاز را فعال میکند تا cAMP را افزایش دهد که کانالهای کاتیونی را باز میکند و منجر به افزایش نفوذپذیری Na، K و +Ca2 روی غشای عصب قطبی شدن غشاء عصبی و denolol میشود.
5. After watching the movie Christmas Story, a 10-year-old boy wanted to see if his tongue would really stick to a frozen pole. Much to his surprise, it did. When he pulled his tongue from the pole, he injured the anterior one-third of his tongue. What sensory nerve arises from this portion of the tongue, where are the cell bodies of these sensory neurons, and where does the nerve terminate?
A. Facial nerve, geniculate ganglion, and gustatory area of the nucleus of the tractus solitarius
B. Vagus nerve, nodose ganglion, and gustatory area of the nucleus ambiguus
C. Chorda tympani branch of the facial nerve, taste buds, and gustatory area of nucleus of the tractus solitarius
D. Glossopharyngeal nerve, petrosal ganglion, and gustatory area of the nucleus ambiguus
E. Glossopharyngeal nerve, taste buds, and caudal portion of the nucleus of the tractus solitarius
5. پس از تماشای فیلم داستان کریسمس، پسری 10 ساله میخواست ببیند آیا واقعاً زبانش به یک میله یخ زده میچسبد؟ با کمال تعجب او این اتفاق افتاد. وقتی زبانش را از تیرک بیرون کشید، یک سوم قدامیزبانش آسیب دید. کدام عصب حسی از این قسمت از زبان سرچشمه میگیرد، بدنه سلولی این نورونهای حسی کجا هستند و عصب به کجا ختم میشود؟
الف. عصب صورت، گانگلیون ژنیکوله و ناحیه چشایی هسته tractus solitarius
ب. عصب واگ، گانگلیون گره و ناحیه چشایی هسته مبهم
ج. شاخه تیمپانی Chorda از عصب صورت، جوانههای چشایی و ناحیه چشایی هسته tractus solitarius
د. عصب گلوفارنکس، گانگلیون پتروزال و ناحیه چشایی هسته مبهم
ی. عصب گلوفارنکس، جوانههای چشایی و بخش دمیهسته tractus solitarius
6. A 37-year-old woman was diagnosed with multiple sclerosis. One of the potential consequences of this disorder is diminished taste and smell sensitivity. What is the relationship between the sensations of taste and smell?
A. Odorant receptors and taste receptors are both innervated by trigeminal nerves.
B. The afferent fibers from odorant receptors and taste receptors terminate on the same second-order neurons in the brainstem.
C. Odors from food enter our nasal passages at the same time that the taste receptors in our mouth arestimulated by the food, and the two chemosensory systems interact to establish the flavor of what we ingest.
D. Olfaction is closely related to gustation because odorant and taste receptors send signals to adjacent regions of the postcentral gyrus of the cortex which are connected via axon collaterals.
6. زن 37 ساله ای به بیماری ام اس مبتلا شد. یکی از پیامدهای بالقوه این اختلال کاهش حساسیت چشایی و بویایی است. چه رابطه ای بین حس چشایی و بویایی وجود دارد؟
الف. گیرندههای بو و گیرندههای چشایی هر دو توسط اعصاب سه قلو عصب دهی میشوند.
ب. فیبرهای آوران از گیرندههای بو و گیرندههای طعم به همان نورونهای مرتبه دوم در ساقه مغز ختم میشوند.
ج. بوهای غذا در همان زمان وارد مجرای بینی ما میشود که گیرندههای چشایی در دهان ما توسط غذا تحریک میشوند و دو سیستم شیمیحسی برای ایجاد طعم و مزه چیزی که میخوریم با هم تعامل دارند.
د. بویایی ارتباط نزدیکی با چشیدن دارد زیرا گیرندههای بو و طعم سیگنالهایی را به نواحی مجاور شکنج پست مرکزی قشر مغز میفرستند که از طریق وثیقههای آکسون به هم متصل هستند.
7. A medical student was doing research on gustatory function with a focus on the anatomy and physiology of taste buds. What are the locations and cellular composition of taste buds?
A. Taste buds are located on the sensory endings of the 7th, 9th, and 10th cranial nerves and contain taste receptors, salivary glands, and basal stem cells.
B. Taste buds are located on the dendrites of the 7th, 9th, and 10th cranial nerves and contain papillae, salivary glands, and taste cells.
C. Taste buds are located in the papillae of the tongue and epiglottis and contain tastants, salivary glands, and basal cells.
D. Taste buds are located in the papillae of the tongue and contain tastants, neuroepithelial cells, and the axons of the 7th, 9th, and 10th cranial nerves.
E. Taste buds are located in the papillae of the tongue and contain taste receptor cells, support cells, and basal cells.
7. یک دانشجوی پزشکی در حال تحقیق در مورد عملکرد چشایی با تمرکز بر آناتومیو فیزیولوژی جوانههای چشایی بود. مکان و ترکیب سلولی جوانههای چشایی چیست؟
الف. جوانههای چشایی در انتهای حسی اعصاب جمجمه ای 7، 9 و 10 قرار دارند و حاوی گیرندههای چشایی، غدد بزاقی و سلولهای بنیادی پایه هستند.
ب. جوانههای چشایی روی دندریتهای اعصاب جمجمه ای 7، 9 و 10 قرار دارند و حاوی پاپیلا، غدد بزاقی و سلولهای چشایی هستند.
ج. جوانههای چشایی در پاپیلای زبان و اپی گلوت قرار دارند و حاوی مواد مزه، غدد بزاقی و سلولهای پایه هستند.
د. جوانههای چشایی در پاپیلای زبان قرار دارند و حاوی تستانتها، سلولهای عصبی اپیتلیال و آکسونهای اعصاب جمجمه ای 7، 9 و 10 هستند.
ی. جوانههای چشایی در پاپیلای زبان قرار دارند و حاوی سلولهای گیرنده چشایی، سلولهای پشتیبان و سلولهای پایه هستند.
8. A 31-year-old woman is a smoker who has had poor oral hygiene for most of her life. In the past few years she has noticed a reduced sensitivity to the taste of foods which she used to enjoy eating. What types of taste receptors may be damaged if she has difficulty sensing sweet and bitter substances?
A. Epithelial sodium channel (sweet) and hyperpolarization-activated cyclic nucleotide-gated cation channel (bitter)
B. Hyperpolarization-activated cyclic nucleotide-gated cation channel (sweet) and epithelial sodium channel (bitter)
C. T2R family of GPCRS (sweet) and metabotropic glutamate receptor (mGluR4, bitter)
D. T1R2 and T1R3 family of GPCRs (sweet) and T2R family of GPCRs (bitter)
E. Metabotropic glutamate receptor (mGluR4, sweet) and epithelial sodium channel (bitter)
8. زن 31 ساله سیگاری است که در بیشتر عمر خود بهداشت دهان و دندان ضعیفی داشته است. در چند سال گذشته او متوجه کاهش حساسیت به طعم غذاهایی شده است که از خوردن آنها لذت میبرد. اگر او در حس کردن مواد شیرین و تلخ مشکل داشته باشد، چه نوع گیرندههای چشایی ممکن است آسیب ببینند؟
الف. کانال سدیم اپیتلیال (شیرین) و کانال کاتیونی حلقوی دردار نوکلئوتیدی فعال شده با هیپرپلاریزاسیون (تلخ)
ب. کانال کاتیونی حلقوی دردار نوکلئوتیدی فعال شده با هیپرپلاریزاسیون (شیرین) و کانال سدیم اپیتلیال (تلخ)
ج. خانواده T2R از GPCRS (شیرین) و گیرنده متابوتروپیک گلوتامات (mGluR4، تلخ)
د. خانواده T1R2 و T1R3 از GPCRها (شیرین) و خانواده T2R از GPCRها (تلخ)
ی. گیرنده متابوتروپیک گلوتامات (mGluR4، شیرین) و کانال سدیم اپیتلیال (تلخ)
کلیک کنید «منابع»
CHAPTER RESOURCES
Adler E, Melichar JK, Nutt DJ, et al: A novel family of mammalian taste receptors. Cell 2000;100:693.
Anholt RRH: Odor recognition and olfactory transduction: The new frontier. Chem Senses 1991;16:421.
Bachmanov AA, Beauchamp GK: Taste receptor genes. Annu Rev Nutrition 2007;27:389.
Gilbertson TA, Damak S, Margolskee RF: The molecular physiology of taste transduction. Curr Opin Neurobiol 2000;10:519.
Gold GH: Controversial issues in vertebrate olfactory transduction. Annu Rev Physiol 1999;61:857.
Heath TP, Melichar JK, Nutt DJ, Donaldson LF. Human taste thresholds are modulated by serotonin and noradrenaline. J Neurosci 2006;26:12664.
Herness HM, Gilbertson TA: Cellular mechanisms of taste transduction. Annu Rev Physiol 1999;61:873.
Kato A, Touhara K. Mammalian olfactory receptors: pharmacology, G protein coupling and desensitization. Cell Mol Life Sci 2009;66:3743.
Lindemann B: Receptors and transduction in taste. Nature 2001;413:219.
Mombaerts P: Genes and ligands for odorant, vomeronasal and taste receptors. Nature Rev Neurosci 2004;5:263.
Reisert J, Restrepo D: Molecular tuning of odorant receptors and its implication for odor signal processing. Chem Senses 2009;34:535.
Ronnett GV, Moon C: G proteins and olfactory signal transduction. Annu Rev Physiol 2002;64:189.
Shepherd GM, Singer MS, Greer CA: Olfactory receptors: A large gene family with broad affinities and multiple functions (Review). Neuroscientist 1996;2:262.
Stern P, Marks J (editors): Making sense of scents. Science 1999;286:703
🚀 با ما همراه شوید!
تازهترین مطالب و آموزشهای مغز و اعصاب را از دست ندهید. با فالو کردن کانال تلگرام آیندهنگاران مغز، از ما حمایت کنید!

 ورود/ ثبت نام با جیمیل
ورود/ ثبت نام با جیمیل
