فیزیولوژی پزشکی گانونگ؛ بخش مرکزی و بخش قشری فوق کلیه

دعای مطالعه [ نمایش ]
بِسْمِ الله الرَّحْمنِ الرَّحیمِ
اَللّهُمَّ اَخْرِجْنى مِنْ ظُلُماتِ الْوَهْمِ
خدايا مرا بيرون آور از تاريكىهاى وهم،
وَ اَكْرِمْنى بِنُورِ الْفَهْمِ
و به نور فهم گرامى ام بدار،
اَللّهُمَّ افْتَحْ عَلَيْنا اَبْوابَ رَحْمَتِكَ
خدايا درهاى رحمتت را به روى ما بگشا،
وَانْشُرْ عَلَيْنا خَزائِنَ عُلُومِكَ بِرَحْمَتِكَ يا اَرْحَمَ الرّاحِمينَ
و خزانههاى علومت را بر ما باز كن به امید رحمتت اى مهربانترين مهربانان.
کتاب «فیزیولوژی پزشکی گانونگ» بهعنوان یکی از جامعترین و معتبرترین منابع در حوزه علوم پزشکی، همچنان مرجع کلیدی برای درک عملکرد پیچیده بدن انسان است. این اثر با تکیه بر تازهترین پژوهشها و توضیحات دقیق از سازوکارهای فیزیولوژیک، پلی میان علوم پایه پزشکی و کاربردهای بالینی ایجاد میکند و نقشی بیبدیل در آموزش، پژوهش و ارتقای دانش سلامت ایفا مینماید.
ترجمه دقیق و علمی این شاهکار توسط برند علمی آیندهنگاران مغز به مدیریت داریوش طاهری، دسترسی فارسیزبانان به مرزهای نوین دانش فیزیولوژی را ممکن ساخته و رسالتی علمی برای ارتقای آموزش پزشکی، فهم عمیقتر سازوکارهای بدن و توسعه روشهای نوین در حوزه سلامت فراهم آورده است.
» کتاب فیزیولوژی پزشکی گانونگ
» » فصل ۱۹: بخش مرکزی و قشری غده فوق کلیوی
در حال ویرایش
» Ganong’s Review of Medical Physiology
»» CHAPTER 19: The Adrenal Medulla & Adrenal Cortex
OBJECTIVES
After studying his chapter, you should be able to:
• Name the three catecholamines secreted by the adrenal medulla and summarize their biosynthesis, metabolism, and function.
• List the stimuli that increase adrenal medullary secretion.
• Differentiate between C18, C19, and C21 steroids and give examples of each.
• Outline the steps involved in steroid biosynthesis in the adrenal cortex.
• Name the plasma proteins that bind adrenocortical steroids and discuss their physiologic role.
• Name the major site of adrenocortical hormone metabolism and the principal metabolites produced from glucocorticoids, adrenal androgens, and aldosterone.
• Describe the mechanisms by which glucocorticoids and aldosterone produce changes in cellular function.
• Define the physiological and pharmacological effects of glucocorticoids.
• Contrast the physiological and pathological effects of adrenal androgens.
• Describe the mechanisms that regulate secretion of glucocorticoids and adrenal sex hormones.
• Explain the actions of aldosterone and describe the mechanisms that regulate aldosterone secretion.
• Describe the main features of the diseases caused by an excess or deficiency of each of the hormones of the adrenal gland.
اهداف
پس از مطالعه فصل، باید بتوانید:
• سه کاتکول آمین ترشح شده از مدولای آدرنال را نام ببرید و بیوسنتز، متابولیسم و عملکرد آنها را خلاصه کنید.
• محرکهایی را که ترشح مدولاری آدرنال را افزایش میدهند فهرست کنید.
• بین استروئیدهای C18، C19 و C21 تفاوت قائل شوید و از هر کدام مثال بزنید.
• مراحل دخیل در بیوسنتز استروئید در قشر آدرنال را تشریح کنید.
• پروتئینهای پلاسما که استروئیدهای قشر آدرنال را متصل میکنند نام ببرید و نقش فیزیولوژیکی آنها را توضیح دهید.
• محل اصلی متابولیسم هورمون قشر آدرنال و متابولیتهای اصلی تولید شده از گلوکوکورتیکوئیدها، آندروژنهای آدرنال و آلدوسترون را نام ببرید.
• مکانیسمهایی را که توسط آن گلوکوکورتیکوئیدها و آلدوسترون تغییراتی در عملکرد سلولی ایجاد میکنند را شرح دهید.
• اثرات فیزیولوژیکی و فارماکولوژیک گلوکوکورتیکوئیدها را تعریف کنید.
• در مقابل اثرات فیزیولوژیکی و پاتولوژیک آندروژنهای آدرنال قرار دهید.
• مکانیسمهای تنظیم کننده ترشح گلوکوکورتیکوئیدها و هورمونهای جنسی آدرنال را شرح دهید.
• عملکردهای آلدوسترون را توضیح دهید و مکانیسمهای تنظیم کننده ترشح آلدوسترون را شرح دهید.
• ویژگیهای اصلی بیماریهای ناشی از کمبود یا زیاد شدن هر یک از هورمونهای غده فوق کلیوی را شرح دهید.

INTRODUCTION
The adrenal glands are endocrine organs that produce several hormones including catecholamines and steroid hormones. There are two adrenal glands, one sitting on top of each kidney (Figure 19-1). Each adrenal gland has an outer cortex that secretes the steroid hormones, mineralocorticoids, glucocorticoids and androgens, and an inner medulla that secretes the catecholamines, epinephrine, norepinephrine, and dopamine.
مقدمه
غدد فوق کلیوی اندامهای غدد درون ریز هستند که چندین هورمون از جمله کاتکولامینها و هورمونهای استروئیدی تولید میکنند. دو غده فوق کلیوی وجود دارد که یکی در بالای هر کلیه قرار دارد (شکل 19-1). هر غده فوق کلیوی دارای یک قشر بیرونی است که هورمونهای استروئیدی، مینرالوکورتیکوئیدها، گلوکوکورتیکوئیدها و آندروژنها را ترشح میکند و یک مدولای داخلی که کاتکولامینها، اپی نفرین، نوراپی نفرین و دوپامین را ترشح میکند.

FIGURE 19-1 Human adrenal glands. Adrenocortical tissue is yellow; adrenal medullary tissue is blue. Note the location of the adrenals at the superior pole of each kidney. Also shown are extra-adrenal sites (gray) at which cortical and medullary tissue is sometimes found. (Reproduced with permission from Williams RH: Textbook of Endocrinology, 4th ed. St. Louis, MO: Saunders; 1968.)
شکل 19-1 غدد فوق کلیوی انسان. بافت قشر آدرنال زرد است. بافت مدولاری آدرنال آبی است. به محل غده فوق کلیوی در قطب فوقانی هر کلیه توجه کنید. همچنین مکانهای خارج آدرنال (خاکستری) نشان داده شدهاند که گاهی اوقات بافت قشر و مدولاری در آنها یافت میشود. (تکثیر شده با اجازه ویلیامز RH: کتاب درسی غدد درون ریز، ویرایش چهارم سنت لوئیس، MO: ساندرز؛ 1968.)
The adrenal cortex secretes glucocorticoids (eg, cortisol) which are steroids with widespread effects on the metabolism of carbohydrate and protein; and a mineralocorticoid (aldosterone) essential to the maintenance of Na+ balance and extracellular fluid (ECF) volume.Mineralocorticoids and the glucocorticoids are necessary for survival. It is also a secondary site of androgen synthesis, secreting sex hormones such as testosterone, which can exert effects on reproductive function. Adrenocortical secretion is controlled primarily by adrenocorticotropic hormone (ACTH), from the anterior pituitary (Chapter 20), but mineralocorticoid secretion is also subject to independent control by circulating factors, of which the most important is angiotensin II, a peptide formed in the bloodstream by the action of renin.
قشر آدرنال گلوکوکورتیکوئیدها (مثلاً کورتیزول) ترشح میکند که استروئیدهایی با اثرات گسترده بر متابولیسم کربوهیدرات و پروتئین هستند. و یک مینرالوکورتیکوئید (آلدوسترون) برای حفظ تعادل +Na و حجم مایع خارج سلولی (ECF) ضروری است. مینرالوکورتیکوئیدها و گلوکوکورتیکوئیدها برای بقا ضروری هستند. همچنین محل ثانویه سنتز آندروژن است و هورمونهای جنسی مانند تستوسترون ترشح میکند که میتواند بر عملکرد تولید مثل تأثیر بگذارد. ترشح قشر آدرنال عمدتاً توسط هورمون آدرنوکورتیکوتروپیک (ACTH) از هیپوفیز قدامیکنترل میشود (فصل 20)، اما ترشح مینرالوکورتیکوئید نیز تحت کنترل مستقل توسط عوامل گردشی است که مهمترین آن آنژیوتانسین II است، یک پپتید که در جریان خون با اثر رنین تشکیل میشود.
The adrenal medulla is in effect a sympathetic ganglion in which the postganglionic neurons have lost their axons and become secretory cells. The cells secrete when stimulated by the preganglionic nerve fibers that reach the gland via the splanchnic nerves. Adrenal medullary hormones (epinephrine, norepinephrine, and dopamine) work mostly to prepare the body for emergencies, the so-called “fight-or-flight” responses.
مدولای فوق کلیوی در واقع یک گانگلیون سمپاتیک است که در آن نورونهای پس گانگلیونی آکسونهای خود را از دست داده و به سلولهای ترشحی تبدیل شده اند. سلولها زمانی ترشح میشوند که توسط رشتههای عصبی پیش گانگلیونی تحریک شوند که از طریق اعصاب splanchnic به غده میرسند. هورمونهای مدولاری آدرنال (اپی نفرین، نوراپی نفرین و دوپامین) بیشتر برای آماده سازی بدن برای شرایط اضطراری، به اصطلاح پاسخهای “جنگ یا گریز” عمل میکنند.
ADRENAL MORPHOLOGY
The adrenal medulla, which constitutes 28% of the mass of the adrenal gland, is made up of interlacing cords of densely innervated granule-containing cells that abut on venous sinuses. Two cell types can be distinguished morphologically: an epinephrine-secreting type that has larger, less dense granules; and a norepinephrine-secreting type in which smaller, very dense granules fail to fill the vesicles in which they are contained. In humans, 90% of the cells are the epinephrine-secreting type and 10% are the norepinephrine-secreting type. The type of cell that secretes dopamine is unknown. Paraganglia, small groups of cells resembling those in the adrenal medulla, are found near the thoracic and abdominal sympathetic ganglia (Figure 19-1).
مورفولوژی آدرنال
مدولای آدرنال، که 28 درصد از جرم غده فوق کلیوی را تشکیل میدهد، از طنابهای به هم پیوسته ای از سلولهای حاوی گرانول عصب دهی متراکم تشکیل شده است که بر روی سینوسهای وریدی قرار دارند. دو نوع سلول را میتوان از نظر مورفولوژیکی متمایز کرد: یک نوع ترشح کننده اپی نفرین که دارای دانههای بزرگتر و کم تراکم است. و یک نوع ترشح کننده نوراپی نفرین که در آن گرانولهای کوچکتر و بسیار متراکم نمیتوانند وزیکولهایی را که در آن قرار دارند پر کنند. در انسان 90 درصد سلولها از نوع ترشح کننده اپی نفرین و 10 درصد از نوع ترشح کننده نوراپی نفرین هستند. نوع سلولی که دوپامین ترشح میکند ناشناخته است. پاراگانگلیون، گروههای کوچکی از سلولهای شبیه به سلولهای مدولای آدرنال، در نزدیکی گانگلیونهای سمپاتیک سینهای و شکمییافت میشوند (شکل 19-1).
In adult mammals, the adrenal cortex is divided into three zones (Figure 19- 2). The outer zona glomerulosa is made up of whorls of cells that are continuous with the columns of cells that form the zona fasciculata. These columns are separated by venous sinuses. The inner portion of the zona fasciculata merges into the zona reticularis, where the cell columns become interlaced in a network. The zona glomerulosa makes up 15% of the mass of the adrenal gland; the zona fasciculata, 50%; and the zona reticularis, 7%. The adrenocortical cells contain abundant lipid, especially in the outer portion of the zona fasciculata. All three cortical zones secrete corticosterone, but the active enzymatic mechanism for aldosterone biosynthesis is limited to the zona glomerulosa, whereas the enzymatic mechanisms for forming cortisol and sex hormones are found in the two inner zones. Furthermore, subspecialization occurs within the inner two zones, with the zona fasciculata secreting mostly glucocorticoids and the zona reticularis secreting mainly sex hormones.
در پستانداران بالغ، قشر آدرنال به سه ناحیه تقسیم میشود (شکل 19-2). زونا گلومرولوزا خارجی از حلقههای سلولی که با ستونهای سلولی که زونا فاسیکولاتا را تشکیل میدهند پیوسته تشکیل شده است. این ستونها توسط سینوسهای وریدی از هم جدا میشوند. بخش داخلی زونا فاسیکولاتا به زونا رتیکولاریس ادغام میشود، جایی که ستونهای سلولی در یک شبکه به هم میپیوندند. زونا گلومرولوزا 15 درصد از توده غده فوق کلیوی را تشکیل میدهد. Zona fasciculata، 50%؛ و زونا رتیکولاریس، 7٪. سلولهای قشر آدرنال حاوی چربی فراوانی هستند، به ویژه در قسمت بیرونی زونا فاسیکولاتا. هر سه ناحیه قشر کورتیکوسترون ترشح میکنند، اما مکانیسم آنزیمیفعال برای بیوسنتز آلدوسترون به زونا گلومرولوزا محدود میشود، در حالی که مکانیسمهای آنزیمیبرای تشکیل کورتیزول و هورمونهای جنسی در دو ناحیه داخلی یافت میشود. علاوه بر این، زیرتخصصی شدن در دو ناحیه داخلی رخ میدهد، به طوری که زونا فاسیکولاتا عمدتاً گلوکوکورتیکوئیدها ترشح میکند و زونا رتیکولاریس عمدتاً هورمونهای جنسی ترشح میکند.

FIGURE 19-2 Section through an adrenal gland showing both the medulla and the zones of the cortex, as well as the hormones they secrete. (Reproduced with permission from Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology: The Mechanisms of Body Function, 11th ed. New York, NY: McGraw-Hill; 2008.)
شکل 19-2 از طریق غده فوق کلیوی که هم بصل النخاع و نواحی قشر و هم هورمونهایی که ترشح میکنند را نشان میدهد. (تکثیر شده با اجازه Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology: The Mechanisms of Body Function, ed. 11 New York, NY: McGraw-Hill; 2008.)
Arterial blood reaches the adrenal from many small branches of the phrenic and renal arteries and the aorta. From a plexus in the capsule, blood flows through the cortex to the sinusoids of the medulla. The medulla is also supplied by a few arterioles that pass directly to it from the capsule. In most species, including humans, blood from the medulla flows into a central adrenal vein. The blood flow through the adrenal is large, as it is in most endocrine glands.
خون شریانی از بسیاری از شاخههای کوچک سرخرگهای فرنیک و کلیوی و آئورت به آدرنال میرسد. از شبکه ای در کپسول، خون از طریق قشر به سینوسیهای مدولا جریان مییابد. بصل النخاع نیز توسط چند شریان که مستقیماً از کپسول به آن عبور میکند، تامین میشود. در بیشتر گونهها، از جمله انسان، خون از مدولا به داخل سیاهرگ مرکزی آدرنال جریان مییابد. جریان خون از طریق آدرنال زیاد است، همانطور که در اکثر غدد درون ریز وجود دارد.
During fetal life, the human adrenal is large and under pituitary control, but the three zones of the permanent cortex represent only 20% of the gland. The remaining 80% is the large fetal adrenal cortex, which undergoes rapid degeneration at the time of birth. A major function of this fetal adrenal is synthesis and secretion of sulfate conjugates of androgens that are converted in the placenta to estrogens (see Chapter 22). No structure is comparable to the human fetal adrenal in laboratory animals.
در طول زندگی جنینی، غده فوق کلیوی انسان بزرگ و تحت کنترل هیپوفیز است، اما سه ناحیه قشر دائمیتنها 20٪ از غده را نشان میدهد. 80 درصد باقیمانده قشر بزرگ آدرنال جنین است که در زمان تولد دچار انحطاط سریع میشود. عملکرد اصلی این آدرنال جنینی، سنتز و ترشح مزدوجات سولفات آندروژن است که در جفت به استروژن تبدیل میشود (به فصل 22 مراجعه کنید). هیچ ساختاری با آدرنال جنین انسان در حیوانات آزمایشگاهی قابل مقایسه نیست.
An important function of the zona glomerulosa, in addition to aldosterone synthesis, is the formation of new cortical cells. The adrenal medulla does not regenerate, but when the inner two zones of the cortex are removed, a new zona fasciculata and zona reticularis regenerate from glomerular cells attached to the capsule. Small capsular remnants regrow large pieces of adrenocortical tissue. Immediately after hypophysectomy, the zona fasciculata and zona reticularis begin to atrophy, whereas the zona glomerulosa is unchanged because of the action of angiotensin II on this zone. The ability to secrete aldosterone and conserve Na+ is normal for some time after hypophysectomy, but in long- standing hypopituitarism, aldosterone deficiency may develop, apparently because of the absence of a pituitary factor that maintains the responsiveness of the zona glomerulosa. Injections of ACTH and stimuli that cause endogenous ACTH secretion produce hypertrophy of the zona fasciculata and zona reticularis but actually decrease, rather than increase, the size of the zona glomerulosa.
یکی از عملکردهای مهم زونا گلومرولوزا، علاوه بر سنتز آلدوسترون، تشکیل سلولهای قشر جدید است. مدولای آدرنال بازسازی نمیشود، اما زمانی که دو ناحیه داخلی قشر جدا میشوند، یک زونا فاسیکولاتا و زونا رتیکولاریس جدید از سلولهای گلومرولی متصل به کپسول بازسازی میشوند. بقایای کپسولی کوچک، تکههای بزرگی از بافت قشر آدرنال را دوباره رشد میدهند. بلافاصله پس از هیپوفیزکتومی، زونا فاسیکولاتا و زونا رتیکولاریس شروع به آتروفی میکنند، در حالی که زونا گلومرولوزا به دلیل اثر آنژیوتانسین II در این ناحیه بدون تغییر است. توانایی ترشح آلدوسترون و حفظ Na+ برای مدتی پس از هیپوفیزیکتومیطبیعی است، اما در هیپوفیزیکتومیطولانی مدت، کمبود آلدوسترون ممکن است ظاهر شود، ظاهراً به دلیل عدم وجود یک فاکتور هیپوفیز که پاسخگویی زونا گلومرولوزا را حفظ میکند. تزریق ACTH و محرکهایی که باعث ترشح درونزای ACTH میشوند، هیپرتروفی زونا فاسیکولاتا و زونا رتیکولاریس را ایجاد میکنند، اما در واقع به جای افزایش، اندازه زونا گلومرولوزا را کاهش میدهند.
The cells of the adrenal cortex contain large amounts of smooth endoplasmic reticulum, which is involved in the steroid-forming process. Other steps in steroid biosynthesis occur in the mitochondria. The structure of steroid-secreting cells is very similar throughout the body. The typical features of such cells are shown in Figure 19-3.
سلولهای قشر آدرنال حاوی مقادیر زیادی شبکه آندوپلاسمیصاف است که در فرآیند تشکیل استروئید نقش دارد. مراحل دیگر در بیوسنتز استروئید در میتوکندری رخ میدهد. ساختار سلولهای ترشح کننده استروئید در سراسر بدن بسیار شبیه است. ویژگیهای معمولی چنین سلولهایی در شکل 19-3 نشان داده شده است.
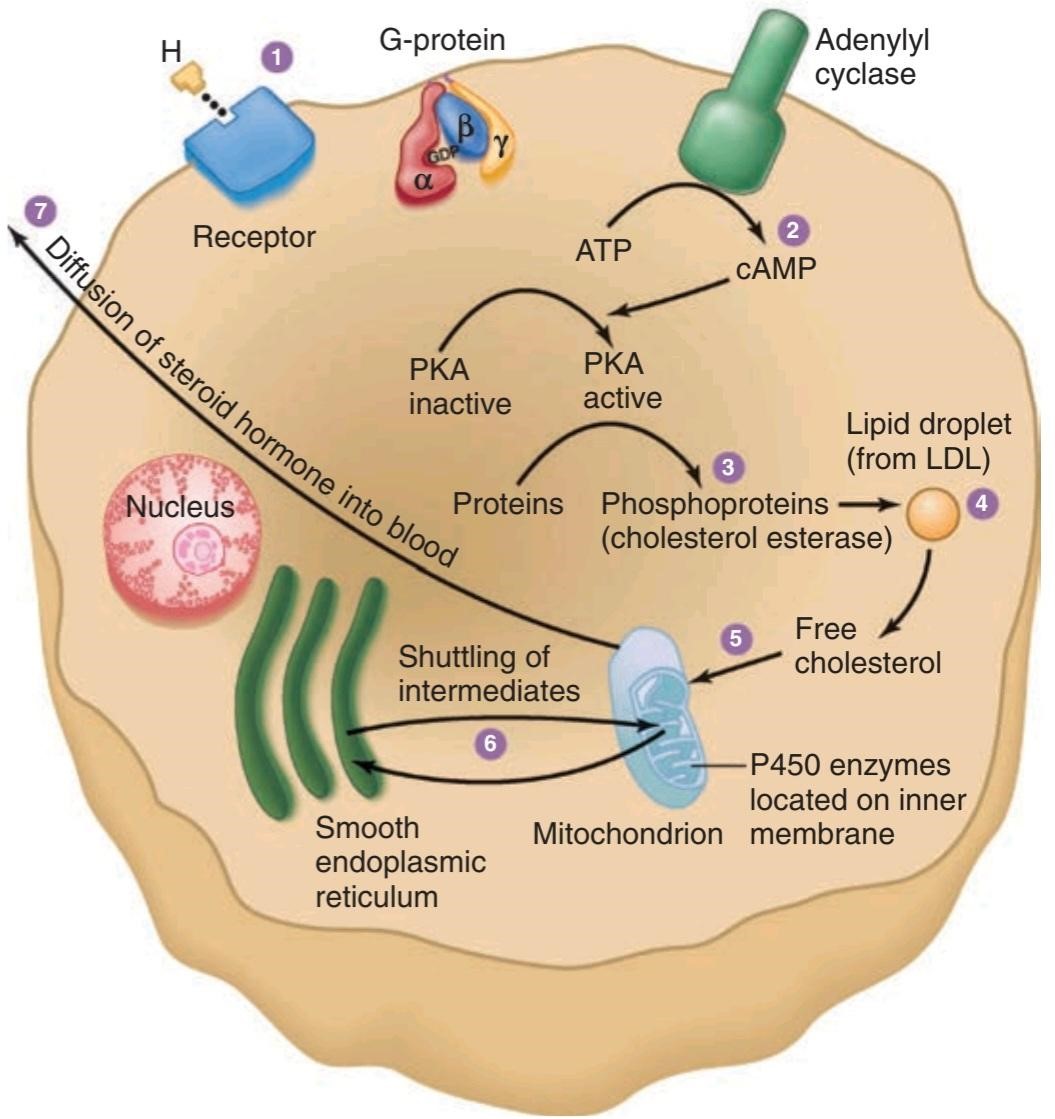
FIGURE 19-3 Schematic overview of the structures of steroid-secreting cells and the intracellular pathway of steroid synthesis. LDL, low-density lipoprotein; PKA, protein kinase A. (Reproduced with permission from
Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology: The Mechanisms of Body Function, 11th ed. New York, NY: McGraw-Hill; 2008.)
شکل 19-3 نمای کلی ساختار سلولهای ترشح کننده استروئید و مسیر درون سلولی سنتز استروئید. LDL، لیپوپروتئین با چگالی کم؛ PKA، پروتئین کیناز A. (تکثیر شده با مجوز از
Widmaier EP, Raff H, Strang KT: Vander’s Human Physiology: The Mechanisms of Body Function, ed 11th. نیویورک، نیویورک: McGraw-Hill; 2008.)
ADRENAL MEDULLA: STRUCTURE & FUNCTION OF MEDULLARY HORMONES
CATECHOLAMINES
Norepinephrine, epinephrine, and small amounts of dopamine are synthesized by the adrenal medulla. Cats and some other species secrete mainly norepinephrine, but in dogs and humans, most of the catecholamine output in the adrenal vein is epinephrine. Norepinephrine also enters the circulation from noradrenergic nerve endings.
مدولای آدرنال: ساختار و عملکرد هورمونهای مدولاری
کاتکولامینها
نوراپی نفرین، اپی نفرین و مقادیر کمیدوپامین توسط بصل الکلیف آدرنال سنتز میشوند. گربهها و برخی گونههای دیگر عمدتا نوراپی نفرین ترشح میکنند، اما در سگها و انسانها، بیشتر کاتکول آمین خروجی در سیاهرگ آدرنال اپی نفرین است. نوراپی نفرین نیز از انتهای عصب نورآدرنرژیک وارد گردش خون میشود.
The structures of norepinephrine, epinephrine, and dopamine and the pathways for their biosynthesis and metabolism are discussed in Chapter 7. Norepinephrine is formed by hydroxylation and decarboxylation of tyrosine, and epinephrine by methylation of norepinephrine. Phenylethanolamine- N- methyltransferase (PNMT), the enzyme that catalyzes the formation of epinephrine from norepinephrine, is found in appreciable quantities only in the brain and the adrenal medulla. Adrenal medullary PNMT is induced by glucocorticoids. Although relatively large amounts are required, the glucocorticoid concentration is high in the blood draining from the cortex to the medulla. After hypophysectomy, the glucocorticoid concentration of this blood falls and epinephrine synthesis is decreased. In addition, glucocorticoids are apparently necessary for the normal development of the adrenal medulla; in 21ẞ- hydroxylase deficiency, glucocorticoid secretion is reduced during fetal life and the adrenal medulla is dysplastic. In untreated 21ẞ-hydroxylase deficiency, circulating catecholamines are low after birth.
ساختار نوراپی نفرین، اپی نفرین و دوپامین و مسیرهای بیوسنتز و متابولیسم آنها در فصل 7 مورد بحث قرار گرفته است. نوراپی نفرین از هیدروکسیلاسیون و دکربوکسیلاسیون تیروزین و اپی نفرین با متیلاسیون نوراپی نفرین تشکیل میشود. فنیل اتانول آمین- N- متیل ترانسفراز (PNMT)، آنزیمیکه تشکیل اپی نفرین از نوراپی نفرین را کاتالیز میکند، به مقدار قابل توجهی فقط در مغز و مدولای آدرنال یافت میشود. PNMT مدولاری آدرنال توسط گلوکوکورتیکوئیدها القا میشود. اگرچه مقادیر نسبتاً زیادی مورد نیاز است، غلظت گلوکوکورتیکوئید در خونی که از قشر به بصل النخاع تخلیه میشود، زیاد است. پس از هیپوفیزکتومی، غلظت گلوکوکورتیکوئید این خون کاهش مییابد و سنتز اپی نفرین کاهش مییابد. علاوه بر این، گلوکوکورتیکوئیدها ظاهراً برای رشد طبیعی مدولای آدرنال ضروری هستند. در کمبود هیدروکسیلاز 21، ترشح گلوکوکورتیکوئید در طول زندگی جنینی کاهش یافته و مدولای آدرنال دیسپلاستیک است. در کمبود 21-هیدروکسیلاز درمان نشده، کاتکول آمینهای در گردش پس از تولد کم هستند.
In plasma, about 95% of the dopamine and 70% of the norepinephrine and epinephrine are conjugated to sulfate. Sulfate conjugates are inactive and their function is unsettled. In recumbent humans, the normal plasma level of free norepinephrine is about 300 pg/mL (1.8 nmol/L). On standing, the level increases 50-100% (Figure 19-4). The plasma norepinephrine level is generally unchanged after adrenalectomy, but the free epinephrine level, which is normally about 30 pg/mL (0.16 nmol/L), falls to essentially zero. The epinephrine found in tissues other than the adrenal medulla and the brain is for the most part absorbed from the bloodstream rather than synthesized in situ. Interestingly, low levels of epinephrine reappear in the blood some time after bilateral adrenalectomy, and these levels are regulated like those secreted by the adrenal medulla. They may come from cells such as the intrinsic cardiac adrenergic (ICA) cells (see Chapter 13), but their exact source is unknown.
در پلاسما حدود 95 درصد دوپامین و 70 درصد نوراپی نفرین و اپی نفرین به سولفات کونژوگه میشوند. کونژوگههای سولفات غیر فعال بوده و عملکرد آنها ته نشین نشده است. در انسانهای دراز کشیده، سطح طبیعی نوراپی نفرین آزاد در پلاسما حدود 300 pg/mL (1.8 نانومول در لیتر) است. در حالت ایستاده، سطح 50-100٪ افزایش مییابد (شکل 19-4). سطح نوراپی نفرین پلاسما به طور کلی پس از آدرنالکتومیبدون تغییر است، اما سطح اپی نفرین آزاد که معمولاً حدود 30 pg/mL (0.16 نانومول در لیتر) است، اساساً به صفر میرسد. اپی نفرین موجود در بافتهای دیگر به جز مدولای آدرنال و مغز بیشتر از جریان خون جذب میشود تا درجا سنتز شود. جالب اینجاست که سطوح پایین اپی نفرین مدتی پس از آدرنالکتومیدوطرفه مجدداً در خون ظاهر میشود و این سطوح مانند سطوح ترشح شده توسط بصل الکلیه آدرنال تنظیم میشود. آنها ممکن است از سلولهایی مانند سلولهای آدرنرژیک قلبی ذاتی (ICA) باشند (به فصل 13 مراجعه کنید)، اما منبع دقیق آنها ناشناخته است.

FIGURE 19-4 Norepinephrine and epinephrine levels in human venous blood in various physiologic and pathologic states. Note that the horizontal scales are different. The numbers to the left in parentheses are the numbers of subjects tested. In each case, the vertical dashed line identifies the threshold plasma concentration at which detectable physiologic changes are observed. (Modified and reproduced with permission from Cryer PE: Physiology and pathophysiology of the human sympathoadrenal neuroendocrine system. N Engl J Med 1980; Aug 21; 303(8):436-444.)
شکل 19-4 سطوح نوراپی نفرین و اپی نفرین در خون وریدی انسان در حالات مختلف فیزیولوژیک و پاتولوژیک. توجه داشته باشید که مقیاسهای افقی متفاوت است. اعداد سمت چپ داخل پرانتز، تعداد موضوعات مورد آزمایش است. در هر مورد، خط چین عمودی آستانه غلظت پلاسمایی را که در آن تغییرات فیزیولوژیکی قابل تشخیص مشاهده میشود، مشخص میکند. (اصلاح و تکثیر شده با مجوز Cryer PE: Physiology and pathophysiology of the human neuroendocrine sympathoadrenal system. N Engl J Med 1980; Aug 21; 303(8):436-444.)
Plasma dopamine levels are normally very low, about 0.13 nmol/L. Most plasma dopamine is thought to be derived from sympathetic noradrenergic ganglia.
سطح دوپامین پلاسما به طور معمول بسیار پایین است، حدود 0.13 نانومول در لیتر. تصور میشود که بیشتر دوپامین پلاسما از گانگلیونهای نورآدرنرژیک سمپاتیک مشتق میشود.
The catecholamines have a half-life of about 2 min in the circulation. For the most part, they are methoxylated and then oxidized to 3-methoxy-4- hydroxymandelic acid (vanillylmandelic acid [VMA]; see Chapter 7). About 50% of the secreted catecholamines appear in the urine as free or conjugated metanephrine and normetanephrine, and 35% as VMA. Only small amounts of free norepinephrine and epinephrine are excreted. In normal humans, about 30 μg of norepinephrine, 6 μg of epinephrine, and 700 μg of VMA are excreted per day.
نیمه عمر کاتکول آمینها در گردش خون حدود 2 دقیقه است. در بیشتر موارد، آنها متوکسیله میشوند و سپس به اسید 3-متوکسی-4- هیدروکسی ماندلیک اکسید میشوند (وانیلیل ماندلیک اسید [VMA]؛ به فصل 7 مراجعه کنید). حدود 50 درصد از کاتکول آمینهای ترشح شده در ادرار به صورت متانفرین آزاد یا کونژوگه و نورمتانفرین و 35 درصد به صورت VMA ظاهر میشوند. فقط مقادیر کمیاز نوراپی نفرین آزاد و اپی نفرین دفع میشود. در انسان عادی روزانه حدود 30 میکروگرم نوراپی نفرین، 6 میکروگرم اپی نفرین و 700 میکروگرم VMA دفع میشود.
OTHER SUBSTANCES SECRETED BY THE ADRENAL MEDULLA
In the medulla, norepinephrine and epinephrine are stored in granules with ATP. The granules also contain chromogranin A (see Chapter 7). Secretion is initiated by acetylcholine released from the preganglionic neurons that innervate the secretory cells. Acetylcholine activates cation channels allowing Ca2+ to enter the cells from the ECF and trigger the exocytosis of the granules. In this fashion, catecholamines, ATP, and proteins from the granules are all released into the blood together.
سایر موادی که توسط مدولای آدرنال ترشح میشود
در بصل النخاع، نوراپی نفرین و اپی نفرین در دانههایی با ATP ذخیره میشوند. گرانولها همچنین حاوی کروموگرانین A هستند (به فصل 7 مراجعه کنید). ترشح توسط استیل کولین آزاد شده از نورونهای پیش گانگلیونی که سلولهای ترشحی را عصب دهی میکنند، آغاز میشود. استیل کولین کانالهای کاتیونی را فعال میکند که اجازه میدهد +Ca2 از ECF وارد سلولها شود و اگزوسیتوز گرانولها را تحریک کند. در این روش، کاتکول آمینها، ATP و پروتئینهای گرانولها همه با هم در خون آزاد میشوند.
Epinephrine-containing cells of the medulla also contain and secrete opioid peptides (see Chapter 7). The precursor molecule is preproenkephalin. Most of the circulating metenkephalin comes from the adrenal medulla. The circulating opioid peptides do not cross the blood-brain barrier.
سلولهای حاوی اپی نفرین در بصل النخاع نیز حاوی پپتیدهای مخدر هستند و ترشح میکنند (به فصل 7 مراجعه کنید). مولکول پیش ساز پیش پروانکفالین است. بیشتر متنکفالین در گردش از بصل الکلیه آدرنال میآید. پپتیدهای اپیوئیدی در گردش از سد خونی مغزی عبور نمیکنند.
Adrenomedullin, a vasodepressor polypeptide found in the adrenal medulla, is discussed in Chapter 32.
آدرنو مدولین، یک پلی پپتید فشار دهنده عروقی که در بصل الکلیف آدرنال یافت میشود، در فصل 32 مورد بحث قرار گرفته است.
EFFECTS OF EPINEPHRINE & NOREPINEPHRINE
In addition to mimicking the effects of noradrenergic nervous discharge, norepinephrine and epinephrine exert metabolic effects that include glycogenolysis in liver and skeletal muscle, mobilization of free fatty acids (FFA), increased plasma lactate, and stimulation of the metabolic rate. The effects of norepinephrine and epinephrine are brought about by actions on two classes of receptors: a- and ẞ-adrenergic receptors. a-Receptors are subdivided into two groups, a1 and α2 receptors, and ẞ-receptors into three groups, B1, B2, and B3 receptors (see Chapter 7). There are three subtypes of a1-receptors and three subtypes of a2-receptors (see Table 7-2).
اثرات اپی نفرین و نوراپی نفرین
نوراپی نفرین و اپی نفرین علاوه بر تقلید از اثرات ترشحات عصبی نورآدرنرژیک، اثرات متابولیکی نیز دارند که شامل گلیکوژنولیز در کبد و ماهیچههای اسکلتی، بسیج اسیدهای چرب آزاد (FFA)، افزایش لاکتات پلاسما و تحریک سرعت متابولیسم میشود. اثرات نوراپی نفرین و اپی نفرین با اعمال بر روی دو دسته از گیرندهها ایجاد میشود: گیرندههای a- و ẞ-آدرنرژیک. گیرندههای a به دو گروه، گیرندههای a1 و α2، و گیرندههای ẞ به سه گروه، گیرندههای B1، B2 و B3 تقسیم میشوند (به فصل 7 مراجعه کنید). سه زیرگروه گیرنده a1 و سه زیرگروه گیرنده a2 وجود دارد (جدول 7-2 را ببینید).
Norepinephrine and epinephrine both increase the force and rate of contraction of the isolated heart. These responses are mediated by B1-receptors. The catecholamines also increase myocardial excitability, causing extrasystoles and, occasionally, more serious cardiac arrhythmias. Norepinephrine produces vasoconstriction in most if not all organs via a1-receptors, but epinephrine dilates the blood vessels in skeletal muscle and the liver via ẞ2-receptors. This usually overbalances the vasoconstriction produced by epinephrine elsewhere, and the total peripheral resistance drops. When norepinephrine is infused slowly in normal animals or humans, the systolic and diastolic blood pressures rise. The hypertension stimulates the carotid and aortic baroreceptors, producing reflex bradycardia that overrides the direct cardioacceleratory effect of norepinephrine. Consequently, cardiac output per minute falls. Epinephrine causes a widening of the pulse pressure but because baroreceptor stimulation is insufficient to obscure the direct effect of the hormone on the heart, cardiac rate and output increase. These changes are summarized in Figure 19-5.
نوراپی نفرین و اپی نفرین هر دو نیرو و سرعت انقباض قلب جدا شده را افزایش میدهند. این پاسخها توسط گیرندههای B1 انجام میشود. کاتکول آمینها همچنین تحریک پذیری میوکارد را افزایش میدهند و باعث اکستراسیستول و گاهی اوقات آریتمیهای قلبی جدی تر میشوند. نوراپی نفرین باعث انقباض عروق در اکثر اندامها و نه در تمام اندامها از طریق گیرندههای a1 میشود، اما اپی نفرین رگهای خونی در ماهیچههای اسکلتی و کبد را از طریق گیرندههای ẞ2 گشاد میکند. این معمولا انقباض عروق تولید شده توسط اپی نفرین در جاهای دیگر را بیش از حد متعادل میکند و مقاومت کلی محیطی کاهش مییابد. هنگامیکه نوراپی نفرین به آرامیدر حیوانات یا انسانهای عادی تزریق میشود، فشار خون سیستولیک و دیاستولیک افزایش مییابد. فشار خون بالا، بارورسپتورهای کاروتید و آئورت را تحریک میکند و برادی کاردی انعکاسی ایجاد میکند که اثر مستقیم شتاب دهنده قلبی نوراپی نفرین را نادیده میگیرد. در نتیجه، برون ده قلبی در هر دقیقه کاهش مییابد. اپی نفرین باعث افزایش فشار نبض میشود، اما از آنجایی که تحریک گیرنده بارور برای پنهان کردن اثر مستقیم هورمون بر قلب کافی نیست، ضربان قلب و برون ده افزایش مییابد. این تغییرات در شکل 19-5 خلاصه شده است.

FIGURE 19-5 Circulatory changes produced in humans by the slow intravenous infusion of epinephrine and norepinephrine.
شکل 19-5 تغییرات گردش خون در انسان توسط انفوزیون داخل وریدی آهسته اپی نفرین و نوراپی نفرین ایجاد میشود.
Catecholamines increase alertness (see Chapter 14). Epinephrine and norepinephrine are equally potent in this regard, although in humans epinephrine usually evokes more anxiety and fear.
کاتکول آمینها هوشیاری را افزایش میدهند (به فصل 14 مراجعه کنید). اپی نفرین و نوراپی نفرین در این زمینه به یک اندازه قوی هستند، اگرچه در انسان اپی نفرین معمولاً اضطراب و ترس بیشتری را برمیانگیزد.
The catecholamines have several different actions that affect blood glucose. Epinephrine and norepinephrine both cause glycogenolysis. They produce this effect via ẞ-adrenergic receptors that increase cyclic adenosine monophosphate (CAMP), with activation of phosphorylase, and via a-adrenergic receptors that increase intracellular Ca2+ (see Chapter 7). In addition, the catecholamines increase the secretion of insulin and glucagon via ẞ-adrenergic mechanisms and inhibit the secretion of these hormones via a-adrenergic mechanisms.
کاتکول آمینها چندین اثر متفاوت دارند که بر گلوکز خون تأثیر میگذارد. اپی نفرین و نوراپی نفرین هر دو باعث گلیکوژنولیز میشوند. آنها این اثر را از طریق گیرندههای آدرنرژیک که آدنوزین مونوفسفات حلقوی (CAMP) را افزایش میدهند، با فعال شدن فسفوریلاز، و از طریق گیرندههای a-آدرنرژیک که Ca2+ داخل سلولی را افزایش میدهند، ایجاد میکنند (به فصل 7 مراجعه کنید). علاوه بر این، کاتکول آمینها ترشح انسولین و گلوکاگون را از طریق مکانیسمهای آدرنرژیک افزایش میدهند و از طریق مکانیسمهای آدرنرژیک از ترشح این هورمونها جلوگیری میکنند.
Norepinephrine and epinephrine also produce a prompt rise in the metabolic rate that is independent of the liver and a smaller, delayed rise that is abolished by hepatectomy and coincides with the rise in blood lactate concentration. The initial rise in metabolic rate may be due to cutaneous vasoconstriction, which decreases heat loss and leads to a rise in body temperature, or to increased muscular activity, or both. The second rise is probably due to oxidation of lactate in the liver. Mice unable to make norepinephrine or epinephrine because their dopamine ẞ-hydroxylase gene is knocked out are intolerant of cold, but surprisingly, their basal metabolic rate is elevated. The cause of this elevation is unknown.
نوراپی نفرین و اپی نفرین همچنین باعث افزایش سریع نرخ متابولیسم میشوند که مستقل از کبد است و افزایش کوچکتر و تاخیری ایجاد میکند که با هپاتکتومیاز بین میرود و همزمان با افزایش غلظت لاکتات خون است. افزایش اولیه در سرعت متابولیسم ممکن است به دلیل انقباض عروق پوستی باشد که از دست دادن گرما را کاهش میدهد و منجر به افزایش دمای بدن یا افزایش فعالیت عضلانی یا هر دو میشود. افزایش دوم احتمالاً به دلیل اکسیداسیون لاکتات در کبد است. موشهایی که قادر به ساخت نوراپی نفرین یا اپی نفرین نیستند، زیرا ژن دوپامین ẞ-هیدروکسیلاز آنها حذف شده است، سرما را تحمل نمیکنند، اما در کمال تعجب، میزان متابولیسم پایه آنها افزایش یافته است. علت این ارتفاع ناشناخته است.
When injected, epinephrine and norepinephrine cause an initial rise in plasma K+ because of release of K+ from the liver and then a prolonged fall in plasma K+ because of an increased entry of K+ into skeletal muscle that is mediated by B2-adrenergic receptors. Some evidence suggests that activation of a-receptors opposes this effect.
هنگامیکه تزریق میشود، اپی نفرین و نوراپی نفرین باعث افزایش اولیه +K پلاسما به دلیل آزاد شدن +K از کبد و سپس کاهش طولانی مدت +K پلاسما به دلیل افزایش ورود +K به ماهیچه اسکلتی میشوند که توسط گیرندههای -B2 آدرنرژیک واسطه میشود. برخی شواهد نشان میدهد که فعال شدن گیرندههای a با این اثر مخالف است.
The increases in plasma norepinephrine and epinephrine that are needed to produce the various effects listed above have been determined by infusion of catecholamines in resting humans. In general, the threshold for the cardiovascular and the metabolic effects of norepinephrine is about 1500 pg/mL, that is, about five times the resting value (Figure 19-4). Epinephrine, on the other hand, produces tachycardia when the plasma level is about 50 pg/mL, that is, about twice the resting value. The threshold for increased systolic blood pressure and lipolysis is about 75 pg/mL; the threshold for hyperglycemia, increased plasma lactate, and decreased diastolic blood pressure is about 150 pg/mL; and the threshold for the a-mediated decrease in insulin secretion is about 400 pg/mL. Plasma epinephrine often exceeds these thresholds. On the other hand, plasma norepinephrine rarely exceeds the threshold for its cardiovascular and metabolic effects, and most of its effects are due to its local release from postganglionic sympathetic neurons. Most adrenal medullary tumors (pheochromocytomas) secrete norepinephrine or epinephrine or both and produce sustained hypertension. However, 15% of epinephrine-secreting tumors secrete this catecholamine episodically, producing intermittent bouts of palpitations, headache, glycosuria, and extreme systolic hypertension. These same symptoms are produced by intravenous injection of a large dose of epinephrine.
افزایش نوراپی نفرین و اپی نفرین پلاسما که برای ایجاد اثرات مختلف ذکر شده در بالا مورد نیاز است با تزریق کاتکول آمینها در انسانهای در حال استراحت مشخص شده است. به طور کلی، آستانه برای اثرات قلبی عروقی و متابولیک نوراپی نفرین حدود 1500 pg/mL است، یعنی حدود پنج برابر مقدار استراحت (شکل 19-4). از طرف دیگر، اپی نفرین زمانی که سطح پلاسما حدود 50 pg/mL باشد، یعنی حدود دو برابر مقدار استراحت، تاکی کاردی ایجاد میکند. آستانه افزایش فشار خون سیستولیک و لیپولیز حدود 75 pg/mL است. آستانه هیپرگلیسمی، افزایش لاکتات پلاسما و کاهش فشار خون دیاستولیک حدود 150 pg/mL است. و آستانه کاهش با واسطه a در ترشح انسولین حدود 400 pg/mL است. اپی نفرین پلاسما اغلب از این آستانه فراتر میرود. از سوی دیگر، نوراپی نفرین پلاسما به ندرت از آستانه اثرات قلبی عروقی و متابولیکی خود فراتر میرود و بیشتر اثرات آن به دلیل آزادسازی موضعی آن از نورونهای سمپاتیک پس گانگلیونی است. اکثر تومورهای مدولاری آدرنال (فئوکروموسیتومها) نوراپی نفرین یا اپی نفرین یا هر دو ترشح میکنند و باعث افزایش فشار خون پایدار میشوند. با این حال، 15 درصد از تومورهای ترشح کننده اپی نفرین، این کاتکول آمین را به صورت اپیزودیک ترشح میکنند و باعث ایجاد حملات متناوب تپش قلب، سردرد، گلیکوزوری و فشار خون شدید سیستولیک میشود. همین علائم با تزریق داخل وریدی دوز زیادی از اپی نفرین ایجاد میشود.
EFFECTS OF DOPAMINE
The physiologic function of the dopamine in the circulation is unknown. However, injected dopamine produces renal vasodilation, probably by acting on a specific dopaminergic receptor. It also produces vasodilation in the mesentery. Elsewhere, it produces vasoconstriction, probably by releasing norepinephrine, and it has a positive inotropic effect on the heart by an action on ẞ1-adrenergic receptors. The net effect of moderate doses of dopamine is an increase in systolic pressure and no change in diastolic pressure. Because of these actions, dopamine is useful in the treatment of traumatic and cardiogenic shock (see Chapter 32).
اثرات دوپامین
عملکرد فیزیولوژیکی دوپامین در گردش خون ناشناخته است. با این حال، دوپامین تزریقی باعث اتساع عروق کلیه میشود، احتمالاً با اثر بر روی یک گیرنده دوپامینرژیک خاص. همچنین باعث اتساع عروق در مزانتر میشود. در جاهای دیگر، احتمالاً با آزاد کردن نوراپی نفرین، انقباض عروق ایجاد میکند و با اثر بر گیرندههای ۱-آدرنرژیک، اثر اینوتروپیک مثبت بر قلب دارد. اثر خالص دوزهای متوسط دوپامین افزایش فشار سیستولیک و عدم تغییر در فشار دیاستولیک است. به دلیل این اعمال، دوپامین در درمان شوک تروماتیک و قلبی مفید است (به فصل 32 مراجعه کنید).
Dopamine is made in the renal cortex. It causes natriuresis and may exert this effect by inhibiting renal Na, K, ATPase.
دوپامین در قشر کلیه ساخته میشود. باعث ناتریورز میشود و ممکن است این اثر را با مهار Na, K, ATPase کلیه اعمال کند.
REGULATION OF ADRENAL MEDULLARY SECRETION
NEURAL CONTROL
Certain drugs act directly on the adrenal medulla, but physiologic stimuli affect medullary secretion through the nervous system. Catecholamine secretion is low in basal states, but the secretion of epinephrine and, to a lesser extent, that of norepinephrine is reduced even further during sleep.
تنظیم ترشح مدولاری آدرنال
کنترل عصبی
برخی داروها مستقیماً بر روی بصل الکلیف آدرنال تأثیر میگذارند، اما محرکهای فیزیولوژیک بر ترشح مدولاری از طریق سیستم عصبی تأثیر میگذارد. ترشح کاتکولامین در حالتهای پایه کم است، اما ترشح اپی نفرین و تا حدی نوراپی نفرین در طول خواب حتی بیشتر کاهش مییابد.
Increased adrenal medullary secretion is part of the diffuse sympathetic discharge provoked in emergency situations, which Cannon called the “emergency function of the sympathoadrenal system.” The ways in which this discharge prepares the individual for flight or fight are described in Chapter 13, and the increases in plasma catecholamines under various conditions are shown in Figure 19-4.
افزایش ترشح مدولاری آدرنال بخشی از ترشحات سمپاتیک منتشر است که در شرایط اضطراری تحریک میشود، که کانن آن را “عملکرد اورژانسی سیستم سمپاتوآدرنال” نامید. راههایی که این ترشحات فرد را برای پرواز یا مبارزه آماده میکند در فصل 13 توضیح داده شده است و افزایش کاتکول آمینهای پلاسما در شرایط مختلف در شکل 19-4 نشان داده شده است.
The metabolic effects of circulating catecholamines are probably important, especially in certain situations. The calorigenic action of catecholamines in animals exposed to cold is an example, and so is the glycogenolytic effect (see Chapter 24) in combating hypoglycemia.
اثرات متابولیکی کاتکول آمینهای در گردش احتمالاً مهم است، به ویژه در شرایط خاص. اثر کالری زایی کاتکول آمینها در حیواناتی که در معرض سرما قرار گرفته اند یک مثال است، و همچنین اثر گلیکوژنولیتیک (به فصل 24 مراجعه کنید) در مبارزه با هیپوگلیسمی.
SELECTIVE SECRETION
When adrenal medullary secretion is increased, the ratio of norepinephrine to epinephrine in the adrenal effluent is generally unchanged. However,norepinephrine secretion tends to be selectively increased by emotional stresses with which the individual is familiar, whereas epinephrine secretion rises selectively in situations in which the individual does not know what to expect.
ترشح انتخابی
هنگامیکه ترشح مدولاری آدرنال افزایش مییابد، نسبت نوراپی نفرین به اپی نفرین در پساب آدرنال به طور کلی بدون تغییر است. با این حال، ترشح نوراپی نفرین به طور انتخابی توسط استرسهای احساسی که فرد با آن آشنا است افزایش مییابد، در حالی که ترشح اپی نفرین به طور انتخابی در موقعیتهایی که فرد نمیداند چه انتظاری دارد افزایش مییابد.
ADRENAL CORTEX: STRUCTURE & BIOSYNTHESIS OF ADRENOCORTICAL HORMONES
CLASSIFICATION & STRUCTURE
The hormones of the adrenal cortex are derivatives of cholesterol. Like cholesterol, bile acids, vitamin D, and ovarian and testicular steroids, they contain the cyclopentanoperhydrophenanthrene nucleus (Figure 19-6). Gonadal and adrenocortical steroids are of three types: C21 steroids, which have a two-carbon side chain at position 17; C19 steroids, which have a keto or hydroxyl group at position 17; and C18 steroids, which, in addition to a 17-keto or hydroxyl group, have no angular methyl group attached to position 10. The adrenal cortex secretes primarily C21 and C19 steroids. Most of the C19 steroids have a keto group at position 17 and are therefore called 17-ketosteroids. The C21 steroids that have a hydroxyl group at the 17 position in addition to the side chain are often called 17-hydroxycorticoids or 17-hydroxycorticosteroids.
قشر آدرنال: ساختار و بیوسنتز هورمونهای غده آدرنال
طبقه بندی و ساختار
هورمونهای قشر آدرنال مشتقات کلسترول هستند. مانند کلسترول، اسیدهای صفراوی، ویتامین D و استروئیدهای تخمدانی و بیضه، آنها حاوی هسته سیکلوپنتانوپیدروفنانترن هستند (شکل 19-6). استروئیدهای غدد جنسی و قشر آدرنال سه نوع هستند: استروئیدهای C21 که دارای یک زنجیره جانبی دو کربنه در موقعیت 17 هستند. استروئیدهای C19 که دارای گروه کتو یا هیدروکسیل در موقعیت 17 هستند. و استروئیدهای C18، که علاوه بر یک گروه 17-کتو یا هیدروکسیل، هیچ گروه متیل زاویه ای متصل به موقعیت 10 ندارند. قشر آدرنال عمدتاً استروئیدهای C21 و C19 را ترشح میکند. اکثر استروئیدهای C19 دارای یک گروه کتو در موقعیت 17 هستند و بنابراین 17-کتوستروئید نامیده میشوند. استروئیدهای C21 که علاوه بر زنجیره جانبی دارای گروه هیدروکسیل در موقعیت 17 هستند، اغلب 17 هیدروکسی کورتیکواستروئید یا 17 هیدروکسی کورتیکواستروئید نامیده میشوند.

FIGURE 19-6 Basic structure of adrenocortical and gonadal steroids. The letters in the formula for cholesterol identify the four basic rings, and the numbers identify the positions in the molecule. As shown here, the angular methyl groups (positions 18 and 19) are usually indicated simply by straight lines.
شکل 19-6 ساختار اساسی استروئیدهای قشر آدرنال و غدد جنسی. حروف موجود در فرمول کلسترول چهار حلقه اصلی را مشخص میکنند و اعداد موقعیتهای موجود در مولکول را مشخص میکنند. همانطور که در اینجا نشان داده شده است، گروههای متیل زاویه ای (موقعیت 18 و 19) معمولاً به سادگی با خطوط مستقیم نشان داده میشوند.
The C19 steroids have androgenic activity. The C21 steroids are classified, using Selye’s terminology, as mineralocorticoids or glucocorticoids. All secreted C21 steroids have both mineralocorticoid and glucocorticoid activity; mineralocorticoids are those in which effects on Na* and K+ excretion predominate and glucocorticoids are those in which effects on glucose and protein metabolism predominate.
استروئیدهای C19 دارای فعالیت آندروژنیک هستند. استروئیدهای C21 با استفاده از اصطلاح Selye به عنوان مینرالوکورتیکوئیدها یا گلوکوکورتیکوئیدها طبقه بندی میشوند. همه استروئیدهای C21 ترشح شده دارای فعالیت مینرالوکورتیکوئید و گلوکوکورتیکوئید هستند. مینرالوکورتیکوئیدها آنهایی هستند که در آنها تأثیر بر دفع Na* و K+ غالب است و گلوکوکورتیکوئیدها آنهایی هستند که تأثیرات آنها بر متابولیسم گلوکز و پروتئین غالب است.
The details of steroid nomenclature and isomerism can be found elsewhere. However, it is pertinent to mention that the Greek letter A indicates a double bond and that the groups that lie above the plane of each of the steroid rings are indicated by the Greek letter ẞ and a solid line (OH), whereas those that lie below the plane are indicated by a and a dashed line (- – -OH). Thus, the C21 steroids secreted by the adrenal have a 4-3-keto configuration in the A ring. In most naturally occurring adrenal steroids, 17-hydroxy groups are in the a configuration, whereas 3-, 11-, and 21-hydroxy groups are in the ß configuration. The 18-aldehyde configuration of naturally occurring aldosterone is the D form. L-aldosterone is physiologically inactive.
جزئیات نامگذاری استروئیدها و ایزومریسم را میتوان در جاهای دیگر یافت. با این حال، لازم به ذکر است که حرف یونانی A یک پیوند دوگانه را نشان میدهد و گروههایی که در بالای صفحه هر یک از حلقههای استروئیدی قرار دارند با حرف یونانی ẞ و یک خط ثابت (OH) نشان داده میشوند، در حالی که آنهایی که در زیر صفحه قرار دارند با a و یک خط چین (OH—) نشان داده میشوند. بنابراین، استروئیدهای C21 ترشح شده از غده فوق کلیوی دارای یک پیکربندی 4-3-keto در حلقه A هستند. در بیشتر استروئیدهای آدرنال طبیعی، گروههای 17 هیدروکسی در یک پیکربندی هستند، در حالی که گروههای 3-، 11- و 21-هیدروکسی در پیکربندی ß هستند. پیکربندی 18 آلدهیدی آلدوسترون طبیعی به شکل D است. ال-آلدوسترون از نظر فیزیولوژیکی غیر فعال است.
SECRETED STEROIDS
Innumerable steroids have been isolated from adrenal tissue, but the only steroids normally secreted in physiologically significant amounts are the mineralocorticoid aldosterone, the glucocorticoids cortisol and corticosterone, and the androgens dehydroepiandrosterone (DHEA) and androstenedione. The structures of these steroids are shown in Figure 19-7 and Figure 19-8. Deoxycorticosterone is a mineralocorticoid that is normally secreted in about the same amount as aldosterone (Table 19-1) but has only 3% of the mineralocorticoid activity of aldosterone. Its effect on mineral metabolism is usually negligible, but in diseases in which its secretion is increased, its effect can be appreciable. Most of the estrogens that are not formed in the ovaries are produced in the circulation from adrenal androstenedione. Almost all the dehydroepiandrosterone is secreted conjugated with sulfate, although most if not all of the other steroids are secreted in the free, unconjugated form (Clinical Box 19-1).
استروئیدهای ترشح شده
استروئیدهای بیشماری از بافت آدرنال جدا شده اند، اما تنها استروئیدهایی که به طور معمول در مقادیر قابل توجه فیزیولوژیکی ترشح میشوند مینرالوکورتیکوئید آلدوسترون، گلوکوکورتیکوئیدهای کورتیزول و کورتیکوسترون، و آندروژنهای دهیدرواپی آندروسترون (DHEA) و آندروستندیون هستند. ساختار این استروئیدها در شکل 19-7 و شکل 19-8 نشان داده شده است. دئوکسی کورتیکوسترون یک مینرالوکورتیکوئید است که به طور معمول تقریباً به اندازه آلدوسترون ترشح میشود (جدول 1-19) اما فقط 3٪ از فعالیت مینرالوکورتیکوئیدی آلدوسترون را دارد. تأثیر آن بر متابولیسم مواد معدنی معمولاً ناچیز است، اما در بیماریهایی که ترشح آن افزایش مییابد، تأثیر آن قابل ملاحظه است. بیشتر استروژنهایی که در تخمدانها تشکیل نمیشوند در گردش خون از آندروستندیون آدرنال تولید میشوند. تقریباً تمام دهیدرواپی آندروسترون به صورت کونژوگه با سولفات ترشح میشود، اگرچه اکثر استروئیدها، اگر نه همه استروئیدهای دیگر به صورت آزاد و غیر کونژوگه ترشح میشوند (باکس بالینی 19-1).
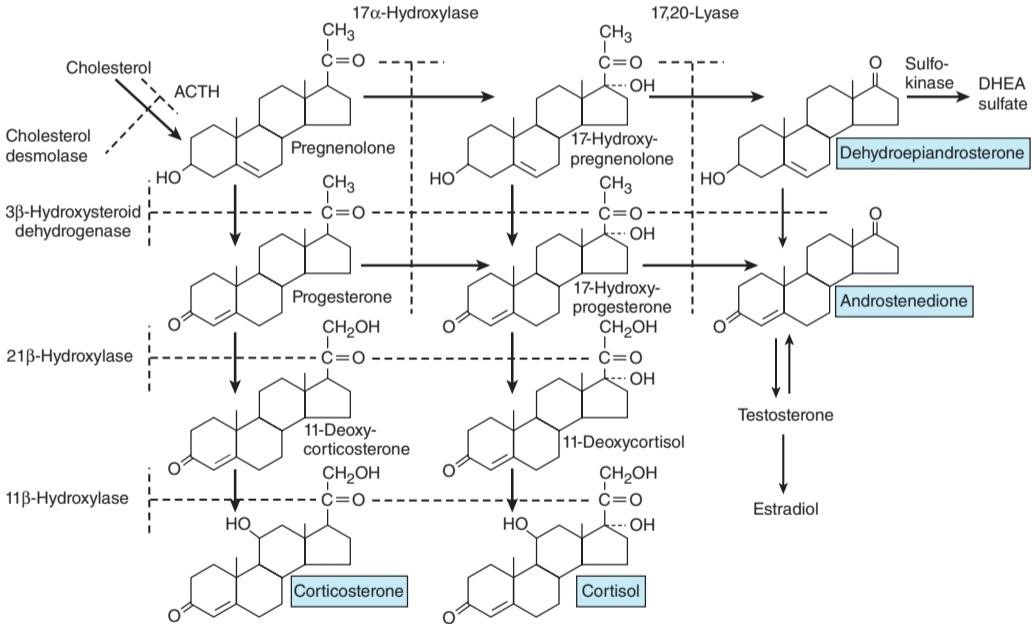
FIGURE 19-7 Outline of hormone biosynthesis in the zona fasciculata and zona reticularis of the adrenal cortex. The major secretory products are underlined. The enzymes for the reactions are shown on the left and at the top of the chart. When a particular enzyme is deficient, hormone production is blocked at the points indicated by the broken lines. ACTH, adrenocorticotropic hormone; DHEA, dehydroepiandrosterone.
شکل 19-7 طرح کلی بیوسنتز هورمون در زونا فاسیکولاتا و زونا رتیکولاریس قشر آدرنال. محصولات ترشحی اصلی زیر خط کشیده شده اند. آنزیمهای واکنشها در سمت چپ و بالای نمودار نشان داده شده اند. هنگامیکه یک آنزیم خاص کمبود داشته باشد، تولید هورمون در نقاطی که با خطوط شکسته نشان داده شده است مسدود میشود. ACTH، هورمون آدرنوکورتیکوتروپیک؛ DHEA، دهیدرواپی آندروسترون.
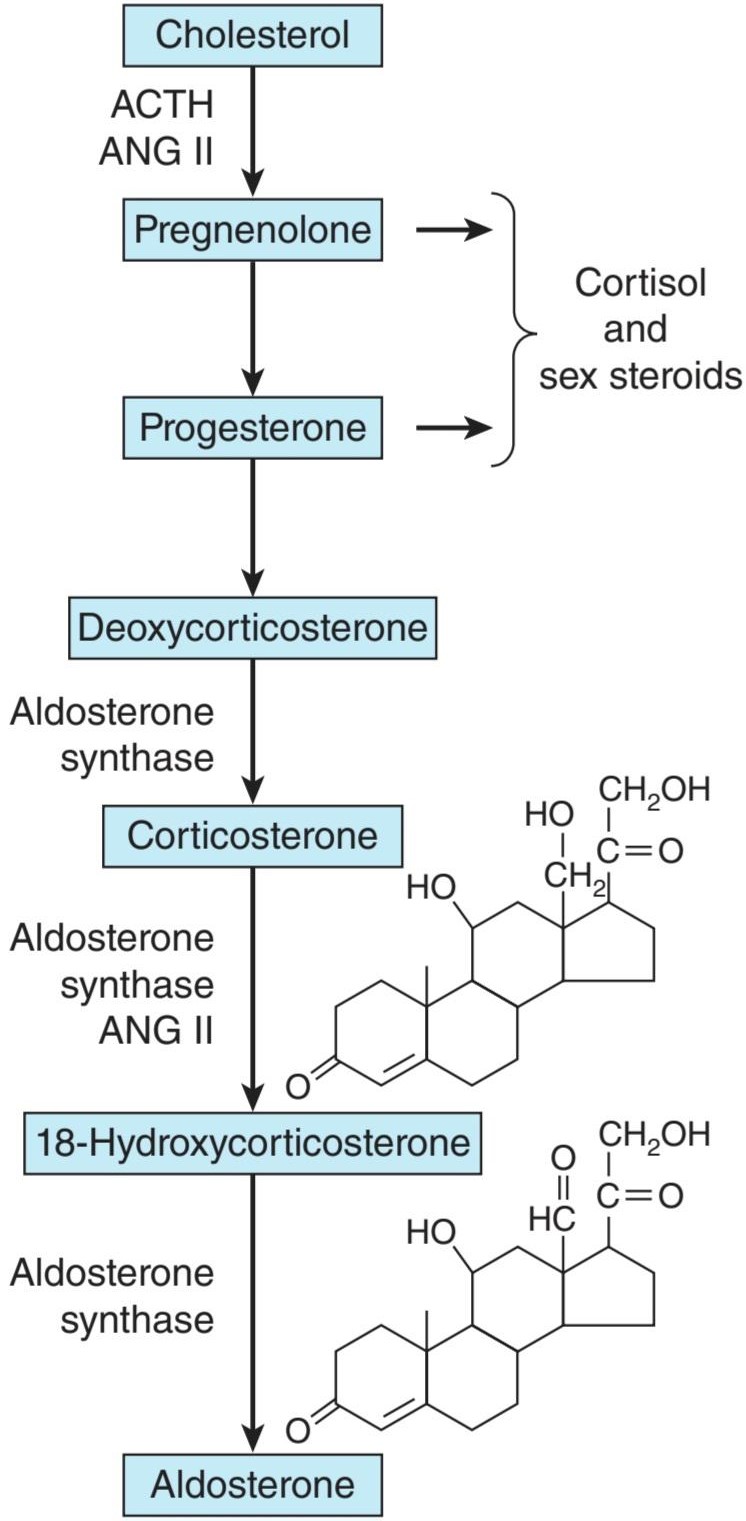
FIGURE 19-8 Hormone synthesis in the zona glomerulosa. The zona glomerulosa lacks 17a-hydroxylase activity, and only the zona glomerulosa can convert corticosterone to aldosterone because it is the only zone that normally contains aldosterone synthase. ACTH, adrenocorticotropic hormone; ANG II, angiotensin II.
شکل 19-8 سنتز هورمون در زونا گلومرولوزا. زونا گلومرولوزا فاقد فعالیت 17a-هیدروکسیلاز است و فقط زونا گلومرولوزا میتواند کورتیکوسترون را به آلدوسترون تبدیل کند زیرا تنها منطقه ای است که به طور معمول حاوی آلدوسترون سنتاز است. ACTH، هورمون آدرنوکورتیکوتروپیک؛ ANG II، آنژیوتانسین II.
TABLE 19-1 Principal adrenocortical hormones in adult humans.
جدول 19-1 هورمونهای اصلی قشر آدرنال در انسان بالغ.
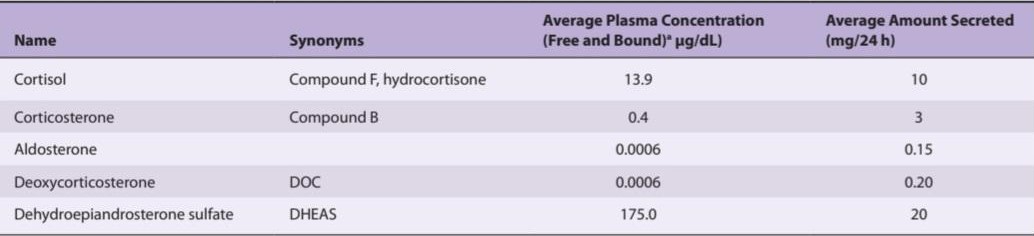
aAll plasma concentration values except DHEAS are fasting morning values after overnight recumbency.
aتمام مقادیر غلظت پلاسما به جز DHEAS مقادیر صبح ناشتا پس از خوابیدن یک شبه است.
CLINICAL BOX 19- 1
Synthetic Steroids
As with many other naturally occurring substances, the activity of adrenocortical steroids can be increased by altering their structure. A number of synthetic steroids are available that have many times the activity of cortisol. The relative glucocorticoid and mineralocorticoid potencies of the natural steroids are compared with those of the synthetic steroids 9a- fluorocortisol, prednisolone, and dexamethasone in Table 19-2. The potency of dexamethasone is due to its high affinity for glucocorticoid receptors and its long half-life. Prednisolone also has a long half-life.
جعبه بالینی 19-1
استروئیدهای مصنوعی
مانند بسیاری از مواد طبیعی دیگر، فعالیت استروئیدهای قشر آدرنال را میتوان با تغییر ساختار آنها افزایش داد. تعدادی از استروئیدهای مصنوعی موجود است که فعالیت آنها چندین برابر کورتیزول است. قدرت نسبی گلوکوکورتیکوئید و مینرالوکورتیکوئید استروئیدهای طبیعی با استروئیدهای مصنوعی 9a- فلوروکورتیزول، پردنیزولون و دگزامتازون در جدول 19-2 مقایسه شده است. قدرت دگزامتازون به دلیل تمایل زیاد آن به گیرندههای گلوکوکورتیکوئید و نیمه عمر طولانی آن است. پردنیزولون نیز نیمه عمر طولانی دارد.
TABLE 19-2 Relative potencies of corticosteroids compared with cortisol.
جدول 19-2 قدرت نسبی کورتیکواستروئیدها در مقایسه با کورتیزول.

aValues are approximations based on liver glycogen deposition or antiinflammatory assays for glucocorticoid activity, and effect on urinary Na+/K+ or maintenance of adrenalectomized animals for mineralocorticoid activity. The last three steroids listed are synthetic compounds that do not occur naturally.
aمقادیر تقریبی بر اساس رسوب گلیکوژن کبدی یا سنجشهای ضد التهابی برای فعالیت گلوکوکورتیکوئید و تأثیر بر +Na+/K ادرار یا نگهداری حیوانات آدرنالکتومیشده برای فعالیت مینرالوکورتیکوئیدی هستند. سه استروئید آخر لیست شده ترکیبات مصنوعی هستند که به طور طبیعی وجود ندارند.
SPECIES DIFFERENCES
In all species from amphibia to humans, the major C21 steroid hormones secreted by adrenocortical tissue appear to be aldosterone, cortisol, and corticosterone, although the ratio of cortisol to corticosterone varies. Birds, mice, and rats secrete corticosterone almost exclusively; dogs secrete approximately equal amounts of the two glucocorticoids; and cats, sheep, monkeys, and humans secrete predominantly cortisol. In humans, the ratio of secreted cortisol to corticosterone is approximately 7:1.
تفاوت گونهها
در همه گونهها از دوزیستان تا انسان، هورمونهای استروئیدی اصلی C21 که توسط بافت قشر آدرنال ترشح میشوند آلدوسترون، کورتیزول و کورتیکوسترون هستند، اگرچه نسبت کورتیزول به کورتیکوسترون متفاوت است. پرندگان، موشها و موشها تقریباً به طور انحصاری کورتیکوسترون ترشح میکنند. سگها تقریباً مقادیر مساوی از دو گلوکوکورتیکوئید ترشح میکنند. و گربهها، گوسفندان، میمونها و انسانها عمدتاً کورتیزول ترشح میکنند. در انسان، نسبت کورتیزول ترشح شده به کورتیکوسترون تقریباً 7:1 است.
STEROID BIOSYNTHESIS
The major paths by which the naturally occurring adrenocortical hormones are synthesized in the body are summarized in Figure 19-7 and 19-8. The precursor of all steroids is cholesterol. Some of the cholesterol is synthesized from acetate, but most of it is taken up from LDL in the circulation. LDL receptors are especially abundant in adrenocortical cells. The cholesterol is esterified and stored in lipid droplets. Cholesterol ester hydrolase catalyzes the formation of free cholesterol in the lipid droplets (Figure 19-9). The cholesterol is transported to mitochondria by a sterol carrier protein. In the mitochondria, it is converted to pregnenolone in a reaction catalyzed by an enzyme known as cholesterol desmolase or side-chain cleavage enzyme. This enzyme, like most of the enzymes involved in steroid biosynthesis, is a member of the cytochrome P450 superfamily and is also known as P450scc or CYP11A1. For convenience, the various names of the enzymes involved in adrenocortical steroid biosynthesis are summarized in Table 19-3.
بیوسنتز استروئیدی
مسیرهای اصلی که توسط آنها هورمونهای قشر آدرنال به طور طبیعی در بدن سنتز میشوند در شکل 19-7 و 19-8 خلاصه شده است. پیش ساز همه استروئیدها کلسترول است. مقداری از کلسترول از استات سنتز میشود، اما بیشتر آن از LDL در گردش خون جذب میشود. گیرندههای LDL به خصوص در سلولهای قشر آدرنال به وفور یافت میشوند. کلسترول استری شده و در قطرات چربی ذخیره میشود. کلسترول استر هیدرولاز تشکیل کلسترول آزاد در قطرات چربی را کاتالیز میکند (شکل 19-9). کلسترول توسط یک پروتئین حامل استرول به میتوکندری منتقل میشود. در میتوکندری، در واکنشی که توسط آنزیمیبه نام کلسترول دزمولاز یا آنزیم برش زنجیره جانبی کاتالیز میشود، به پرگننولون تبدیل میشود. این آنزیم، مانند بسیاری از آنزیمهای دخیل در بیوسنتز استروئید، عضوی از خانواده سیتوکروم P450 است و با نامهای P450scc یا CYP11A1 نیز شناخته میشود. برای سهولت، نامهای مختلف آنزیمهای دخیل در بیوسنتز استروئیدی قشر آدرنال در جدول 19-3 خلاصه شده است.

FIGURE 19-9 Mechanism of action of ACTH on cortisol-secreting cells in the inner two zones of the adrenal cortex. When ACTH binds to its receptor (R), adenylyl cyclase (AC) is activated via Gs. The resulting increase in cAMP activates protein kinase A, and the kinase phosphorylates cholesteryl ester hydrolase (CEH), increasing its activity. Consequently, more free cholesterol is formed and converted to pregnenolone. Note that in the subsequent steps in steroid biosynthesis, products are shuttled between the mitochondria and the smooth endoplasmic reticulum (SER). Corticosterone is also synthesized and secreted. ACTH, adrenocorticotropic hormone; ATP, adenosine triphosphate; CAMP, cyclic adenosine monophosphate.
شکل 19-9 مکانیسم اثر ACTH بر سلولهای ترشح کننده کورتیزول در دو ناحیه داخلی قشر آدرنال. هنگامیکه ACTH به گیرنده خود (R) متصل میشود، آدنیلیل سیکلاز (AC) از طریق Gs فعال میشود. افزایش حاصل در cAMP پروتئین کیناز A را فعال میکند و کیناز فسفریله کلستریل استر هیدرولاز (CEH) و افزایش فعالیت آن میشود. در نتیجه، کلسترول آزاد بیشتری تشکیل شده و به پرگننولون تبدیل میشود. توجه داشته باشید که در مراحل بعدی بیوسنتز استروئیدها، محصولات بین میتوکندری و شبکه آندوپلاسمیصاف (SER) جابجا میشوند. کورتیکوسترون نیز سنتز و ترشح میشود. ACTH، هورمون آدرنوکورتیکوتروپیک؛ ATP، آدنوزین تری فسفات؛ CAMP، آدنوزین مونوفسفات حلقوی.
TABLE 19-3 Nomenclature for adrenal steroidogenic enzymes and their location in adrenal cells.
جدول 19-3 نامگذاری آنزیمهای استروئیدوژن آدرنال و مکان آنها در سلولهای آدرنال.

SER, smooth endoplasmic reticulum.
SER، شبکه آندوپلاسمیصاف.
Pregnenolone moves to the smooth endoplasmic reticulum, where some of it is dehydrogenated to form progesterone in a reaction catalyzed by 3ẞ- hydroxysteroid dehydrogenase. This enzyme has a molecular weight of 46,000 and is not a cytochrome P450. It also catalyzes the conversion of 17α- hydroxypregnenolone to 17a-hydroxyprogesterone, and dehydroepiandrosterone to androstenedione (Figure 22–7) in the smooth endoplasmic reticulum. The 17a-hydroxypregnenolone and the 17a-hydroxyprogesterone are formed from pregnenolone and progesterone, respectively (Figure 19-7) by the action of 17a hydroxylase. This is another mitochondrial P450, and it is also known as P450c17 or CYP17. Located in another part of the same enzyme is 17,20-lyase activity that breaks the 17,20 bond, converting 17a-pregnenolone and 17α- progesterone to the C19 steroids dehydroepiandrosterone and androstenedione.
پرگننولون به سمت شبکه آندوپلاسمیصاف حرکت میکند، جایی که مقداری از آن برای تشکیل پروژسترون در واکنشی که توسط 3- هیدروکسی استروئید دهیدروژناز کاتالیز میشود، هیدروژنه میشود. این آنزیم دارای وزن مولکولی 46000 است و سیتوکروم P450 نیست. همچنین تبدیل 17α-هیدروکسی پرگننولون به 17a-هیدروکسی پروژسترون و دهیدرواپی آندروسترون به آندروستندیون (شکل 22-7) را در شبکه آندوپلاسمیصاف کاتالیز میکند. 17a-hydroxypregnenolone و 17a-hydroxyprogesterone به ترتیب از پرگننولون و پروژسترون (شکل 19-7) با اثر 17a هیدروکسیلاز تشکیل میشوند. این یکی دیگر از P450 میتوکندریایی است و به عنوان P450c17 یا CYP17 نیز شناخته میشود. در قسمت دیگری از همان آنزیم، فعالیت 17،20-لیاز قرار دارد که پیوند 17،20 را میشکند و 17a-pregnenolone و 17α-progesterone را به استروئیدهای C19 دهیدرواپی آندروسترون و آندروستندیون تبدیل میکند.
Hydroxylation of progesterone to 11-deoxycorticosterone and of 17α- hydroxyprogesterone to 11-deoxycortisol occurs in the smooth endoplasmic reticulum. These reactions are catalyzed by 21ẞ-hydroxylase, a cytochrome P450 that is also known as P450c21 or CYP21A2.
هیدروکسیلاسیون پروژسترون به 11-دئوکسی کورتیکوسترون و 17α- هیدروکسی پروژسترون به 11-دئوکسی کورتیزول در شبکه آندوپلاسمیصاف رخ میدهد. این واکنشها توسط 21آ-هیدروکسیلاز، یک سیتوکروم P450 که به نامهای P450c21 یا CYP21A2 نیز شناخته میشود، کاتالیز میشوند.
11-Deoxycorticosterone and the 11-deoxycortisol move back to the mitochondria, where they are 11-hydroxylated to form corticosterone and cortisol. These reactions occur in the zona fasciculata and zona reticularis and are catalyzed by 11ẞ-hydroxylase, a cytochrome P450 also known as P450c11 or CYP11B1.
11-دئوکسی کورتیکوسترون و 11-دئوکسی کورتیزول به میتوکندری برمیگردند، جایی که 11 هیدروکسیله میشوند تا کورتیکوسترون و کورتیزول را تشکیل دهند. این واکنشها در زونا فاسیکولاتا و زونا رتیکولاریس رخ میدهند و توسط 11آ-هیدروکسیلاز، سیتوکروم P450 که با نامهای P450c11 یا CYP11B1 نیز شناخته میشود، کاتالیز میشوند.
In the zona glomerulosa there is no 11ẞ-hydroxylase, but a closely related enzyme called aldosterone synthase is present. This cytochrome P450 is 95% identical to 11ẞ-hydroxylase and is also known as P450c11AS or CYP11B2. The genes that code CYP11B1 and CYP11B2 are both located on chromosome 8. However, aldosterone synthase is normally found only in the zona glomerulosa. The zona glomerulosa also lacks 17α-hydroxylase. This is why the zona glomerulosa makes aldosterone but fails to make cortisol or sex hormones.
در زونا گلومرولوزا 11-هیدروکسیلاز وجود ندارد، اما آنزیمینزدیک به هم به نام آلدوسترون سنتاز وجود دارد. این سیتوکروم P450 95٪ با 11آ-هیدروکسیلاز یکسان است و با نامهای P450c11AS یا CYP11B2 نیز شناخته میشود. ژنهایی که CYP11B1 و CYP11B2 را کد میکنند، هر دو در کروموزوم 8 قرار دارند. با این حال، آلدوسترون سنتاز معمولاً فقط در زونا گلومرولوزا یافت میشود. زونا گلومرولوزا نیز فاقد 17α هیدروکسیلاز است. به همین دلیل است که زونا گلومرولوزا آلدوسترون میسازد اما کورتیزول یا هورمونهای جنسی را نمیسازد.
Furthermore, subspecialization occurs within the inner two zones. The zona fasciculata has more 3ẞ-hydroxysteroid dehydrogenase activity than the zona reticularis, and the zona reticularis has more of the cofactors required for the 17,20-lyase activity of 17α-hydroxylase. Therefore, the zona fasciculata makes more cortisol and corticosterone, and the zona reticularis makes more androgens. Most of the dehydroepiandrosterone that is formed is converted to dehydroepiandrosterone sulfate (DHEAS) by adrenal sulfokinase, and this enzyme is localized in the zona reticularis as well.
علاوه بر این، فوق تخصص در دو منطقه داخلی رخ میدهد. زونا فاسیکولاتا دارای فعالیت 3آ-هیدروکسی استروئید دهیدروژناز بیشتری نسبت به زونا رتیکولاریس است و زونا رتیکولاریس دارای کوفاکتورهای بیشتری برای فعالیت 17،20-لیاز 17α-هیدروکسیلاز است. بنابراین، زونا فاسیکولاتا کورتیزول و کورتیکوسترون بیشتری تولید میکند و زونا رتیکولاریس آندروژن بیشتری تولید میکند. بیشتر دهیدرواپی آندروسترون تشکیل شده توسط سولفوکیناز آدرنال به دهیدرواپی آندروسترون سولفات (DHEAS) تبدیل میشود و این آنزیم در زونا رتیکولاریس نیز موضعی میشود.
ACTION OF ACTH
ACTH binds to high-affinity receptors on the plasma membrane of adrenocortical cells. This activates adenylyl cyclase via G. The resulting reactions (Figure 19-9) lead to a prompt increase in the formation of pregnenolone and its derivatives, with secretion of the latter. Over longer periods, ACTH also increases the synthesis of the P450s involved in the synthesis of glucocorticoids.
عمل ACTH
ACTH به گیرندههای با میل ترکیبی بالا روی غشای پلاسمایی سلولهای قشر آدرنال متصل میشود. این کار آدنیلیل سیکلاز را از طریق G فعال میکند. واکنشهای حاصله (شکل 19-9) منجر به افزایش سریع تشکیل پرگننولون و مشتقات آن با ترشح دومیمیشود. در طول دورههای طولانی تر، ACTH همچنین سنتز P450s را افزایش میدهد که در سنتز گلوکوکورتیکوئیدها نقش دارند.
ACTIONS OF ANGIOTENSIN II
Angiotensin II binds to AT1 receptors (see Chapter 38) in the zona glomerulosa that act via a G-protein to activate phospholipase C. The resulting increase in protein kinase C fosters the conversion of cholesterol to pregnenolone (Figure 19-8) and facilitates the action of aldosterone synthase, resulting in increased secretion of aldosterone.
اقدامات آنژیوتانسین II
آنژیوتانسین II به گیرندههای AT1 متصل میشود (به فصل 38 مراجعه کنید) در زونا گلومرولوزا که از طریق یک پروتئین G برای فعال کردن فسفولیپاز C عمل میکنند. افزایش حاصل در پروتئین کیناز C تبدیل کلسترول به پرگننولون را تقویت میکند (شکل 19-8 باعث افزایش ترشح آلدوسترون میشود). آلدوسترون
ENZYME DEFICIENCIES
The consequences of inhibiting any of the enzyme systems involved in steroid biosynthesis can be predicted from Figures 19-7 and 19-8. Congenital defects in the enzymes lead to deficient cortisol secretion and the syndrome of congenital adrenal hyperplasia. The hyperplasia is due to increased ACTH secretion. Cholesterol desmolase deficiency is fatal in utero because it prevents the placenta from making the progesterone necessary for pregnancy to continue. A cause of severe congenital adrenal hyperplasia in newborns is a loss of function mutation of the gene for the steroidogenic acute regulatory (STAR) protein. This protein is essential in the adrenals and gonads but not in the placenta for the normal movement of cholesterol into the mitochondria to reach cholesterol desmolase, which is located on the matrix space side of the internal mitochondrial membrane (see Chapter 16, Figure 16-1). In its absence, only small amounts of steroids are formed. The degree of ACTH stimulation is marked, resulting eventually in accumulation of large numbers of lipoid droplets in the adrenal. For this reason, the condition is called congenital lipoid adrenal hyperplasia. Because androgens are not formed, female genitalia develop regardless of genetic sex (see Chapter 22). In 3ẞ hydroxysteroid dehydrogenase deficiency, another rare condition, DHEA secretion is increased. This steroid is a weak androgen that can cause some masculinization in females with the disease, but it is not adequate to produce full masculinization of the genitalia in genetic males. Consequently, hypospadias, a condition where the opening of the urethra is on the underside of the penis rather than its tip, is common. In fully developed 17a-hydroxylase deficiency, a third rare condition due to a mutated gene for CYP17, no sex hormones are produced, so female external genitalia are present. However, the pathway leading to corticosterone and aldosterone is intact, and elevated levels of 11-deoxycorticosterone and other mineralocorticoids produce hypertension and hypokalemia. Cortisol is deficient, but this is partially compensated by the glucocorticoid activity of corticosterone.
کمبود آنزیمها
پیامدهای مهار هر یک از سیستمهای آنزیمیدرگیر در بیوسنتز استروئید را میتوان از شکلهای 19-7 و 19-8 پیش بینی کرد. نقص مادرزادی در آنزیمها منجر به کمبود ترشح کورتیزول و سندرم هیپرپلازی مادرزادی آدرنال میشود. هیپرپلازی به دلیل افزایش ترشح ACTH است. کمبود کلسترول دسمولاز در رحم کشنده است زیرا از جفت جلوگیری میکند تا پروژسترون لازم برای ادامه بارداری را بسازد. یکی از دلایل هیپرپلازی مادرزادی آدرنال شدید در نوزادان، از دست دادن جهش عملکردی ژن پروتئین تنظیم کننده حاد استروئیدی (STAR) است. این پروتئین در غدد فوق کلیوی و غدد جنسی ضروری است اما نه در جفت برای حرکت طبیعی کلسترول به داخل میتوکندری برای رسیدن به کلسترول دزمولاز، که در سمت فضای ماتریکس غشای داخلی میتوکندری قرار دارد (نگاه کنید به فصل 16، شکل 16-1). در غیاب آن، فقط مقادیر کمیاستروئید تشکیل میشود. درجه تحریک ACTH مشخص میشود که در نهایت منجر به تجمع تعداد زیادی قطرات چربی در غده فوق کلیوی میشود. به همین دلیل، این بیماری را هیپرپلازی مادرزادی لیپویید آدرنال مینامند. از آنجا که آندروژنها تشکیل نمیشوند، اندام تناسلی زنان بدون توجه به جنسیت ژنتیکی رشد میکنند (به فصل 22 مراجعه کنید). در کمبود 3 هیدروکسی استروئید دهیدروژناز، یکی دیگر از شرایط نادر، ترشح DHEA افزایش مییابد. این استروئید یک آندروژن ضعیف است که میتواند باعث مردانه شدن در زنان مبتلا به این بیماری شود، اما برای ایجاد مردانه شدن کامل دستگاه تناسلی در مردان ژنتیکی کافی نیست. در نتیجه، هیپوسپادیاس، وضعیتی که در آن دهانه مجرای ادرار به جای نوک آلت تناسلی، در قسمت زیرین آلت تناسلی قرار دارد، شایع است. در کمبود کامل 17a-hydroxylase، سومین بیماری نادر ناشی از ژن جهش یافته برای CYP17، هیچ هورمون جنسی تولید نمیشود، بنابراین اندام تناسلی خارجی زنان وجود دارد. با این حال، مسیر منتهی به کورتیکوسترون و آلدوسترون دست نخورده است و سطوح بالای 11-دئوکسی کورتیکوسترون و سایر مینرالوکورتیکوئیدها باعث افزایش فشار خون و هیپوکالمیمیشود. کورتیزول کمبود دارد، اما تا حدی با فعالیت گلوکوکورتیکوئیدی کورتیکوسترون جبران میشود.
Unlike the defects discussed in the preceding paragraph, 21ẞ-hydroxylase deficiency is common, accounting for 90% or more of the enzyme deficiency cases. The 21ẞ-hydroxylase gene, which is in the HLA complex of genes on the short arm of chromosome 6 (see Chapter 3) is one of the most polymorphic in the human genome. Mutations occur at many different sites in the gene, and the abnormalities that are produced therefore range from mild to severe. Production of cortisol and aldosterone are generally reduced, so ACTH secretion and consequently production of precursor steroids are increased. These steroids are converted to androgens, producing virilization. The characteristic pattern that develops in females in the absence of treatment is the adrenogenital syndrome. Masculization may not be marked until later in life and mild cases can be detected only by laboratory tests. In 75% of the cases, aldosterone deficiency causes appreciable loss of Na+ (salt-losing form of adrenal hyperplasia). The resulting hypovolemia can be severe.
برخلاف نقایص مورد بحث در پاراگراف قبل، کمبود 21-هیدروکسیلاز رایج است که 90٪ یا بیشتر از موارد کمبود آنزیم را تشکیل میدهد. ژن 21ẞ-hydroxylase که در مجموعه ژنهای HLA روی بازوی کوتاه کروموزوم 6 قرار دارد (به فصل 3 مراجعه کنید) یکی از چندشکلیترین ژنها در ژنوم انسان است. جهشها در مکانهای مختلف ژن اتفاق میافتند و بنابراین ناهنجاریهایی که ایجاد میشوند از خفیف تا شدید متغیر هستند. تولید کورتیزول و آلدوسترون به طور کلی کاهش مییابد، بنابراین ترشح ACTH و در نتیجه تولید استروئیدهای پیش ساز افزایش مییابد. این استروئیدها به آندروژن تبدیل میشوند و باعث ویریل شدن میشوند. الگوی مشخصی که در زنان در غیاب درمان ایجاد میشود، سندرم آدرنوژنیتال است. مردانگی ممکن است تا اواخر زندگی مشخص نشود و موارد خفیف فقط با آزمایشهای آزمایشگاهی قابل تشخیص است. در 75 درصد موارد، کمبود آلدوسترون باعث از دست دادن قابل ملاحظه Na+ (شکل از دست دادن نمک هیپرپلازی آدرنال) میشود. هیپوولمیحاصل میتواند شدید باشد.
In 11ẞ-hydroxylase deficiency, virilization plus excess secretion of 11- deoxycortisol and 11-deoxycorticosterone take place. Because the former is an active mineralocorticoid, patients with this condition also have salt and water retention and, in two-thirds of the cases, hypertension (hypertensive form of congenital adrenal hyperplasia).
در کمبود 11 هیدروکسیلاز، ویریل شدن به همراه ترشح بیش از حد 11-دئوکسی کورتیزول و 11-دئوکسی کورتیکوسترون اتفاق میافتد. از آنجا که اولی یک مینرالوکورتیکوئید فعال است، بیماران مبتلا به این بیماری همچنین دارای احتباس نمک و آب و در دو سوم موارد فشار خون بالا (شکل پرفشاری خون از هیپرپلازی مادرزادی آدرنال) هستند.
Glucocorticoid treatment is indicated in all of the virilizing forms of congenital adrenal hyperplasia because it repairs the glucocorticoid deficit and inhibits ACTH secretion, reducing the abnormal secretion of androgens and other steroids.
درمان با گلوکوکورتیکوئید در تمام اشکال ویریل کنندههایپرپلازی مادرزادی آدرنال اندیکاسیون دارد زیرا کمبود گلوکوکورتیکوئید را ترمیم میکند و ترشح ACTH را مهار میکند و ترشح غیر طبیعی آندروژنها و سایر استروئیدها را کاهش میدهد.
Expression of the cytochrome P450 enzymes responsible for steroid hormone biosynthesis depends on steroid factor-1 (SF-1), an orphan nuclear receptor. If Ft2-F1, the gene for SF-1, is knocked out, the gonads as well as adrenals fail to develop and additional abnormalities are present at the pituitary and hypothalamic level.
بیان آنزیمهای سیتوکروم P450 مسئول بیوسنتز هورمون استروئیدی به فاکتور 1 استروئیدی (SF-1)، یک گیرنده هسته ای یتیم بستگی دارد. اگر Ft2-F1، ژن SF-1 حذف شود، غدد جنسی و همچنین غدد فوق کلیوی رشد نمیکنند و ناهنجاریهای اضافی در سطح هیپوفیز و هیپوتالاموس وجود دارد.
TRANSPORT, METABOLISM, & EXCRETION OF ADRENOCORTICAL HORMONES
GLUCOCORTICOID BINDING
Cortisol is bound in the circulation to an a globulin called transcortin or corticosteroid-binding globulin (CBG). A minor degree of binding to albumin also takes place. Corticosterone is similarly bound but to a lesser degree. The half-life of cortisol in the circulation is therefore longer (about 60-90 min) than that of corticosterone (50 min). Bound steroids are physiologically inactive (see Chapter 16). In addition, relatively little free cortisol and corticosterone are found in the urine because of protein binding.
انتقال، متابولیسم و دفع هورمونهای غده آدرنوکورتیکال
اتصال گلوکوکورتیکوئید
کورتیزول در گردش خون به یک گلوبولین به نام ترانسکورتین یا گلوبولین اتصال به کورتیکواستروئید (CBG) متصل میشود. میزان کمیاز اتصال به آلبومین نیز اتفاق میافتد. کورتیکوسترون نیز به همین ترتیب محدود میشود اما به میزان کمتری. بنابراین نیمه عمر کورتیزول در گردش خون بیشتر از کورتیکوسترون (50 دقیقه) است (حدود 60 تا 90 دقیقه). استروئیدهای متصل از نظر فیزیولوژیکی غیر فعال هستند (به فصل 16 مراجعه کنید). علاوه بر این، کورتیزول و کورتیکوسترون آزاد نسبتا کمیبه دلیل اتصال به پروتئین در ادرار یافت میشود.
The equilibrium between cortisol and its binding protein and the implications of binding in terms of tissue supplies and ACTH secretion are summarized in Figure 19-10. The bound cortisol functions as a circulating reservoir of hormone that keeps a supply of free cortisol available to the tissues. The relationship is similar to that of T4 and its binding protein (see Chapter 20). At normal levels of total plasma cortisol (13.5 μg/dL or 375 nmol/L), very little free cortisol is present in the plasma, but the binding sites on CBG become saturated when the total plasma cortisol exceeds 20 μg/dL. At higher plasma levels, binding to albumin increases, but the main increase is in the unbound fraction.
تعادل بین کورتیزول و پروتئین اتصال دهنده آن و پیامدهای اتصال از نظر منابع بافتی و ترشح ACTH در شکل 19-10 خلاصه شده است. کورتیزول متصل به عنوان یک مخزن در گردش هورمون عمل میکند که منبع کورتیزول آزاد را در دسترس بافتها نگه میدارد. رابطه مشابه با T4 و پروتئین اتصال آن است (به فصل 20 مراجعه کنید). در سطوح نرمال کورتیزول کل پلاسما (13.5 میکروگرم در دسی لیتر یا 375 نانومول در لیتر)، کورتیزول آزاد بسیار کمیدر پلاسما وجود دارد، اما مکانهای اتصال در CBG زمانی که کل کورتیزول پلاسما از 20 میکروگرم در دسی لیتر فراتر رود، اشباع میشوند. در سطوح پلاسمایی بالاتر، اتصال به آلبومین افزایش مییابد، اما افزایش اصلی در بخش غیر متصل است.

FIGURE 19-10 The interrelationships of free and bound cortisol. The dashed arrow indicates that cortisol inhibits ACTH secretion. The value for free cortisol is an approximation; in most studies, it is calculated by subtracting the protein-bound cortisol from the total plasma cortisol. ACTH, adrenocorticotropic hormone.
شکل 19-10 روابط متقابل کورتیزول آزاد و محدود. فلش چین نشان میدهد که کورتیزول ترشح ACTH را مهار میکند. مقدار کورتیزول آزاد یک تقریبی است. در بیشتر مطالعات، با کم کردن کورتیزول متصل به پروتئین از کل کورتیزول پلاسما محاسبه میشود. ACTH، هورمون آدرنوکورتیکوتروپیک.
CBG is synthesized in the liver and its production is increased by estrogen. CBG levels are elevated during pregnancy and depressed in cirrhosis, nephrosis, and multiple myeloma. When the CBG level rises, more cortisol is bound, and initially the free cortisol level drops. This stimulates ACTH secretion, and more cortisol is secreted until a new equilibrium is reached at which the bound cortisol is elevated but the free cortisol is normal. Changes in the opposite direction occur when the CBG level falls. This explains why pregnant women have high total plasma cortisol levels without symptoms of glucocorticoid excess and, conversely, why some patients with nephrosis have low total plasma cortisol without symptoms of glucocorticoid deficiency.
CBG در کبد سنتز میشود و تولید آن توسط استروژن افزایش مییابد. سطح CBG در دوران بارداری افزایش مییابد و در سیروز، نفروز و میلوم متعدد افسرده میشود. هنگامیکه سطح CBG افزایش مییابد، کورتیزول بیشتری محدود میشود و در ابتدا سطح کورتیزول آزاد کاهش مییابد. این باعث تحریک ترشح ACTH میشود و کورتیزول بیشتری ترشح میشود تا زمانی که تعادل جدیدی حاصل شود که در آن کورتیزول محدود شده بالا میرود اما کورتیزول آزاد طبیعی است. هنگامیکه سطح CBG کاهش مییابد، تغییرات در جهت مخالف رخ میدهد. این توضیح میدهد که چرا زنان باردار سطح کورتیزول تام پلاسما بالا بدون علائم اضافی گلوکوکورتیکوئید دارند و برعکس، چرا برخی از بیماران مبتلا به نفروز کورتیزول تام پلاسما پایین بدون علائم کمبود گلوکوکورتیکوئید دارند.
METABOLISM & EXCRETION OF GLUCOCORTICOIDS
Cortisol is metabolized in the liver, which is the principal site of glucocorticoid catabolism. Most of the cortisol is reduced to dihydrocortisol and then to tetrahydrocortisol, which is conjugated to glucuronic acid (Figure 19-11). The glucuronyl transferase system responsible for this conversion also catalyzes the formation of the glucuronides of bilirubin (see Chapter 28) and a number of hormones and drugs. Competitive inhibition takes place between these substrates for the enzyme system.
متابولیسم و دفع گلوکوکورتیکوئیدها
کورتیزول در کبد، که محل اصلی کاتابولیسم گلوکوکورتیکوئید است، متابولیزه میشود. بیشتر کورتیزول به دی هیدروکورتیزول و سپس به تتراهیدروکورتیزول کاهش مییابد که به اسید گلوکورونیک کونژوگه میشود (شکل 19-11). سیستم گلوکورونیل ترانسفراز مسئول این تبدیل همچنین تشکیل گلوکورونیدهای بیلی روبین (به فصل 28 را ببینید) و تعدادی از هورمونها و داروها را کاتالیز میکند. مهار رقابتی بین این بسترها برای سیستم آنزیمیصورت میگیرد.

FIGURE 19-11 Outline of hepatic metabolism of cortisol.
شکل 19-11 طرح کلی متابولیسم کبدی کورتیزول.
The liver and other tissues contain the enzyme 11ẞ hydroxysteroid dehydrogenase. There are at least two forms of this enzyme. Type 1 catalyzes the conversion of cortisol to cortisone and the reverse reaction, though it functions primarily as a reductase, forming cortisol from corticosterone. Type 2 catalyzes almost exclusively the one-way conversion of cortisol to cortisone. Cortisone is an active glucocorticoid because it is converted to cortisol, and it is well known because of its extensive use in medicine. It is not secreted in appreciable quantities by the adrenal glands. Little, if any, of the cortisone formed in the liver enters the circulation, because it is promptly reduced and conjugated to form tetrahydrocortisone glucuronide. The tetrahydroglucuronide derivatives (“conjugates”) of cortisol and corticosterone are freely soluble. They enter the circulation, where they do not become bound to protein. They are rapidly excreted in the urine.
کبد و سایر بافتها حاوی آنزیم 11 یا هیدروکسی استروئید دهیدروژناز هستند. حداقل دو شکل از این آنزیم وجود دارد. نوع 1 تبدیل کورتیزول به کورتیزون و واکنش معکوس را کاتالیز میکند، اگرچه در درجه اول به عنوان ردوکتاز عمل میکند و کورتیزول را از کورتیکوسترون تشکیل میدهد. نوع 2 تقریباً به طور انحصاری تبدیل یک طرفه کورتیزول به کورتیزون را کاتالیز میکند. کورتیزون یک گلوکوکورتیکوئید فعال است زیرا به کورتیزول تبدیل میشود و به دلیل استفاده زیاد در پزشکی شناخته شده است. به مقدار قابل توجهی از غدد فوق کلیوی ترشح نمیشود. مقدار کمیاز کورتیزون تشکیل شده در کبد وارد گردش خون میشود، زیرا به سرعت کاهش یافته و برای تشکیل تتراهیدروکورتیزون گلوکورونید کونژوگه میشود. مشتقات تتراهیدروگلوکورونید (“کونژوگه”) کورتیزول و کورتیکوسترون آزادانه محلول هستند. آنها وارد گردش خون میشوند، جایی که به پروتئین متصل نمیشوند. آنها به سرعت از طریق ادرار دفع میشوند.
About 10% of the secreted cortisol is converted in the liver to the 17- ketosteroid derivatives of cortisol and cortisone. The ketosteroids are conjugated for the most part to sulfate and then excreted in the urine. Other metabolites, including 20-hydroxy derivatives, are formed. There is an enterohepatic circulation of glucocorticoids and about 15% of the secreted cortisol is excreted in the stool. The metabolism of corticosterone is similar to that of cortisol, except that it does not form a 17-ketosteroid derivative (see Clinical Box 19-2).
حدود 10 درصد از کورتیزول ترشح شده در کبد به مشتقات 17 کتواستروئیدی کورتیزول و کورتیزون تبدیل میشود. کتواستروئیدها بیشتر به سولفات کونژوگه میشوند و سپس از طریق ادرار دفع میشوند. متابولیتهای دیگر، از جمله مشتقات 20-هیدروکسی، تشکیل میشوند. گردش روده کبدی گلوکوکورتیکوئیدها وجود دارد و حدود 15 درصد از کورتیزول ترشح شده در مدفوع دفع میشود. متابولیسم کورتیکوسترون مشابه کورتیزول است، با این تفاوت که یک مشتق 17 کتوستروئیدی تشکیل نمیدهد (به کادر بالینی 19-2 مراجعه کنید).
CLINICAL BOX 19-2
Variations in the Rate of Hepatic Metabolism
The rate of hepatic inactivation of glucocorticoids is depressed in liver disease and, interestingly, during surgery and other stresses. Thus, in stressed humans, the plasma-free cortisol level rises higher than it does with maximal ACTH stimulation in the absence of stress.
جعبه بالینی 19-2
تغییرات در میزان متابولیسم کبدی
میزان غیرفعال شدن گلوکوکورتیکوئیدها در کبد در بیماریهای کبدی و جالب اینکه در حین جراحی و سایر استرسها کاهش مییابد. بنابراین، در انسانهای تحت استرس، سطح کورتیزول بدون پلاسما بالاتر از حداکثر تحریک ACTH در غیاب استرس افزایش مییابد.
ALDOSTERONE
Aldosterone is bound to protein to only a slight extent, and its half-life is short (about 20 min). The amount secreted is small (Table 19-1), and the total plasma aldosterone level in humans is normally about 0.006 μg/dL (0.17 nmol/L), compared with a cortisol level (bound and free) of about 13.5 μg/dL (375 nmol/L). Much of the aldosterone is converted in the liver to the tetrahydroglucuronide derivative, but some is changed in the liver and in the kidneys to an 18-glucuronide. This glucuronide, which is unlike the breakdown products of other steroids, is converted to free aldosterone by hydrolysis at pH 1.0, and it is therefore often referred to as the “acid-labile conjugate.” Less than 1% of the secreted aldosterone appears in the urine in the free form. Another 5% is in the form of the acid-labile conjugate, and up to 40% is in the form of the tetrahydroglucuronide.
آلدوسترون
آلدوسترون فقط تا حدودی به پروتئین متصل میشود و نیمه عمر آن کوتاه است (حدود 20 دقیقه). مقدار ترشح شده اندک است (جدول 1-19)، و سطح کل آلدوسترون پلاسما در انسان به طور معمول حدود 0.006 میکروگرم در دسی لیتر (0.17 نانومول در لیتر)، در مقایسه با سطح کورتیزول (محصول و آزاد) حدود 13.5 میکروگرم در دسی لیتر (375 نانومول در لیتر) است. بیشتر آلدوسترون در کبد به مشتق تتراهیدروگلوکورونید تبدیل میشود، اما مقداری در کبد و در کلیهها به یک گلوکورونید 18 تبدیل میشود. این گلوکورونید، که بر خلاف محصولات تجزیه سایر استروئیدها است، با هیدرولیز در pH 1.0 به آلدوسترون آزاد تبدیل میشود و بنابراین اغلب به عنوان “مجموعه ناپایدار اسیدی” نامیده میشود. کمتر از 1 درصد از آلدوسترون ترشح شده در ادرار به شکل آزاد ظاهر میشود. 5٪ دیگر به شکل مزدوج حساس به اسید و تا 40٪ به شکل تتراهیدروگلوکورونید است.
17-KETOSTEROIDS
The major adrenal androgen is the 17-ketosteroid dehydroepiandrosterone, although androstenedione is also secreted. The 11-hydroxy derivative of androstenedione and the 17-ketosteroids formed from cortisol and cortisone by side chain cleavage in the liver are the only 17-ketosteroids that have an =O or an-OH group in the 11 position (“11-oxy-17-ketosteroids”). Testosterone is also converted to a 17-ketosteroid. Because the daily 17-ketosteroid excretion in normal adults is 15 mg in men and 10 mg in women, about two-thirds of the urinary ketosteroids in men are secreted by the adrenal or formed from cortisol in the liver and about one-third are of testicular origin.
17-کتوستروئیدها
آندروژن اصلی آدرنال، دهیدرواپی آندروسترون 17 کتواستروئیدی است، اگرچه آندروستندیون نیز ترشح میشود. مشتق 11 هیدروکسی آندروستندیون و 17 کتوستروئیدهای تشکیل شده از کورتیزول و کورتیزون توسط شکاف زنجیره جانبی در کبد، تنها 17 کتوستروئیدهایی هستند که دارای یک گروه O= یا an-OH در موقعیت 11 هستند (“11-oxy-17-ketosteroids”). تستوسترون نیز به 17 کتوستروئید تبدیل میشود. از آنجایی که دفع روزانه 17 کتوستروئید در بزرگسالان طبیعی 15 میلی گرم در مردان و 10 میلی گرم در زنان است، حدود دو سوم کتوستروئیدهای ادراری در مردان از غده فوق کلیوی ترشح میشود یا از کورتیزول در کبد تشکیل میشود و حدود یک سوم منشاء بیضه دارند.
Etiocholanolone, one of the metabolites of the adrenal androgens and testosterone, can cause fever when it is unconjugated (see Chapter 17). Certain individuals have episodic bouts of fever due to periodic accumulation in the blood of unconjugated etiocholanolone (“etiocholanolone fever”).
اتیوکولانولون، یکی از متابولیتهای آندروژن آدرنال و تستوسترون، میتواند باعث ایجاد تب در صورت عدم کونژوگه شود (به فصل 17 مراجعه کنید). برخی از افراد به دلیل تجمع دورهای در خون اتیوکولانولون غیرکونژوگه (“تب اتیوکولانولون”) دچار حملات اپیزودیک تب میشوند.
EFFECTS OF ADRENAL ANDROGENS & ESTROGENS
ANDROGENS
Androgens are the hormones that exert masculinizing effects and they promote protein anabolism and growth (see Chapter 23). Testosterone from the testes is the most active androgen and the adrenal androgens have less than 20% of its activity. Secretion of the adrenal androgens is controlled acutely by ACTH and not by gonadotropins. However, the concentration of DHEAS increases until it peaks at about 225 mg/dL in the early 20s, and then falls to very low values in old age (Figure 19-12). These long-term changes are not due to changes in ACTH secretion and appear to be due instead to a rise and then a gradual fall in the lyase activity of 17a-hydroxylase.
اثرات آندروژن و استروژن آدرنال
آندروژنها
آندروژنها هورمونهایی هستند که اثرات مردانه دارند و آنابولیسم و رشد پروتئین را تقویت میکنند (به فصل 23 مراجعه کنید). تستوسترون از بیضهها فعال ترین آندروژن است و آندروژنهای آدرنال کمتر از 20 درصد فعالیت آن را دارند. ترشح آندروژنهای آدرنال به طور حاد توسط ACTH کنترل میشود نه توسط گنادوتروپینها. با این حال، غلظت DHEAS تا زمانی که در اوایل دهه 20 به اوج خود در حدود 225 میلی گرم در دسی لیتر میرسد افزایش مییابد و سپس در سنین بالا به مقادیر بسیار پایین میرسد (شکل 19-12). این تغییرات طولانی مدت به دلیل تغییر در ترشح ACTH نیست و به نظر میرسد در عوض به دلیل افزایش و سپس کاهش تدریجی فعالیت لیاز 17a-hydroxylase باشد.

FIGURE 19-12 Change in serum dehydroepiandrosterone sulfate (DHEAS) with age. The middle line is the mean, and the dashed lines identify +1.96 standard deviations. (Reproduced, with permission, from Smith MR, et al: A radioimmunoassay for the estimation of serum dehydroepiandrosterone sulfate in normal and pathological sera. Clin Chim Acta 1975; Nov 15; 65(1):5–13.)
شکل 19-12 تغییر در دهیدرواپی آندروسترون سولفات سرم (DHEAS) با افزایش سن. خط وسط میانگین است و خطوط چین دار انحراف استاندارد +1.96 را نشان میدهد. (بازتولید شده، با اجازه، از Smith MR، و همکاران: رادیوایمونواسی برای تخمین دهیدرواپی آندروسترون سولفات سرم در سرمهای نرمال و پاتولوژیک. Clin Chim Acta 1975؛ نوامبر 15؛ 65 (1): 5-13.)
All but about 0.3% of the circulating DHEA is conjugated to sulfate (DHEAS). The secretion of adrenal androgens is nearly as great in castrated males and females as it is in normal males, so it is clear that these hormones exert very little masculinizing effect when secreted in normal amounts. However, they can produce appreciable masculinization when secreted in excessive amounts. In adult males, excess adrenal androgens merely accentuate existing characteristics, but in prepubertal boys they can cause precocious development of the secondary sex characteristics without testicular growth (precocious pseudopuberty). In females they cause female pseudo- hermaphroditism and the adrenogenital syndrome. Some health practitioners recommend injections of dehydroepiandrosterone to combat the effects of aging (see Chapter 1), but results to date are controversial at best.
همه به جز حدود 0.3٪ از DHEA در گردش به سولفات (DHEAS) کونژوگه میشود. ترشح آندروژنهای آدرنال در مردان و زنان اخته شده تقریباً به اندازه مردان عادی است، بنابراین واضح است که این هورمونها وقتی در مقادیر طبیعی ترشح میشوند، اثر مردانهسازی بسیار کمیدارند. با این حال، زمانی که در مقادیر بیش از حد ترشح شوند، میتوانند مردانگی قابل توجهی ایجاد کنند. در مردان بالغ، آندروژنهای اضافی آدرنال صرفاً ویژگیهای موجود را برجسته میکنند، اما در پسران قبل از بلوغ میتوانند باعث رشد زودرس ویژگیهای جنسی ثانویه بدون رشد بیضه شوند (بلوغ کاذب زودرس). در زنان باعث ایجاد هرمافرودیتیسم کاذب و سندرم آدرنوژنیتال میشوند. برخی از پزشکان بهداشت تزریق دهیدرواپی آندروسترون را برای مبارزه با اثرات پیری توصیه میکنند (به فصل 1 مراجعه کنید)، اما نتایج تا به امروز در بهترین حالت بحث برانگیز است.
ESTROGENS
The adrenal androgen androstenedione is converted to testosterone and to estrogens (aromatized) in fat and other peripheral tissues. This is an important source of estrogens in postmenopausal women and men (see Chapters 22 and 23).
استروژنها
آندروژن آدرنال آندروستندیون در چربی و سایر بافتهای محیطی به تستوسترون و به استروژن (معطر شده) تبدیل میشود. این منبع مهمیاز استروژن در زنان و مردان یائسه است (فصل 22 و 23 را ببینید).
PHYSIOLOGIC EFFECTS OF GLUCOCORTICOIDS
ADRENAL INSUFFICIENCY
In untreated adrenal insufficiency, Na* loss and shock occurs due to the lack of mineralocorticoid activity, as well as abnormalities of water, carbohydrate, protein, and fat metabolism due to the lack of glucocorticoids. These metabolic abnormalities are eventually fatal despite mineralocorticoid treatment. Small amounts of glucocorticoids correct the metabolic abnormalities, in part directly and in part by permitting other reactions to occur. It is important to separate these physiologic actions of glucocorticoids from the quite different effects produced by large amounts of the hormones.
اثرات فیزیولوژیکی گلوکوکورتیکوئیدها
نارسایی آدرنال
در نارسایی غدد فوق کلیوی درمان نشده، از دست دادن +Na و شوک به دلیل فقدان فعالیت مینرالوکورتیکوئیدها و همچنین ناهنجاریهای متابولیسم آب، کربوهیدرات، پروتئین و چربی به دلیل کمبود گلوکوکورتیکوئیدها رخ میدهد. این ناهنجاریهای متابولیک در نهایت با وجود درمان با مینرالوکورتیکوئید کشنده هستند. مقادیر کمیاز گلوکوکورتیکوئیدها ناهنجاریهای متابولیک را اصلاح میکنند، تا حدی به طور مستقیم و تا حدی با اجازه دادن به سایر واکنشها. مهم است که این اعمال فیزیولوژیک گلوکوکورتیکوئیدها را از اثرات کاملاً متفاوت تولید شده توسط مقادیر زیادی از هورمونها جدا کنیم.
MECHANISM OF ACTION
The multiple effects of glucocorticoids are triggered by binding to glucocorticoid receptors, and the steroid-receptor complexes act as transcription factors that promote the transcription of certain segments of DNA (see Chapter 1). This, in turn, leads via the appropriate mRNAs to synthesis of enzymes that alter cell function. In addition, it seems likely that glucocorticoids have nongenomic actions.
مکانیسم عمل
اثرات متعدد گلوکوکورتیکوئیدها با اتصال به گیرندههای گلوکوکورتیکوئیدی ایجاد میشود و کمپلکسهای گیرنده استروئیدی به عنوان فاکتورهای رونویسی عمل میکنند که رونویسی بخشهای خاصی از DNA را ترویج میکنند (به فصل 1 مراجعه کنید). این به نوبه خود، از طریق mRNAهای مناسب منجر به سنتز آنزیمهایی میشود که عملکرد سلول را تغییر میدهند. علاوه بر این، به نظر میرسد که گلوکوکورتیکوئیدها دارای اثرات غیر ژنومیک هستند.
EFFECTS ON INTERMEDIARY METABOLISM
The actions of glucocorticoids on the intermediary metabolism of carbohydrate, protein, and fat are discussed in Chapter 24. They include increased protein catabolism and increased hepatic glycogenesis and gluconeogenesis. Glucose-6- phosphatase activity is increased, and the plasma glucose level rises. Glucocorticoids exert an anti-insulin action in peripheral tissues and make diabetes worse. However, the brain and the heart are spared, so the increase in plasma glucose provides extra glucose to these vital organs. In diabetics, glucocorticoids raise plasma lipid levels and increase ketone body formation, but in normal individuals, the increase in insulin secretion provoked by the rise in plasma glucose obscures these actions. In adrenal insufficiency, the plasma glucose level is normal as long as an adequate caloric intake is maintained, but fasting causes hypoglycemia that can be fatal. The adrenal cortex is not essential for the ketogenic response to fasting.
اثرات بر متابولیسم واسطه
اعمال گلوکوکورتیکوئیدها بر متابولیسم واسطه کربوهیدرات، پروتئین و چربی در فصل 24 مورد بحث قرار گرفته است. آنها شامل افزایش کاتابولیسم پروتئین و افزایش گلیکوژنز کبدی و گلوکونئوژنز هستند. فعالیت گلوکز-6- فسفاتاز افزایش مییابد و سطح گلوکز پلاسما افزایش مییابد. گلوکوکورتیکوئیدها در بافتهای محیطی اثر ضد انسولین دارند و دیابت را بدتر میکنند. با این حال، مغز و قلب در امان هستند، بنابراین افزایش گلوکز پلاسما، گلوکز اضافی را برای این اندامهای حیاتی فراهم میکند. در افراد دیابتی، گلوکوکورتیکوئیدها سطح لیپیدهای پلاسما را افزایش میدهند و تشکیل بدن کتون را افزایش میدهند، اما در افراد عادی، افزایش ترشح انسولین ناشی از افزایش گلوکز پلاسما این اعمال را مبهم میکند. در نارسایی آدرنال، سطح گلوکز پلاسما تا زمانی که دریافت کالری کافی حفظ شود، طبیعی است، اما روزه داری باعث هیپوگلیسمیمیشود که میتواند کشنده باشد. قشر آدرنال برای پاسخ کتوژنیک به روزه ضروری نیست.
PERMISSIVE ACTION
Small amounts of glucocorticoids must be present for a number of metabolic reactions to occur, although the glucocorticoids do not produce the reactions by themselves. This effect is called their permissive action. Permissive effects include the requirement for glucocorticoids to be present for glucagon and catecholamines to exert their calorigenic effects (see above and Chapter 24), for catecholamines to exert their lipolytic effects, and for catecholamines to produce pressor responses and bronchodilation.
عمل مجاز
مقادیر کمیاز گلوکوکورتیکوئیدها باید برای تعدادی از واکنشهای متابولیک وجود داشته باشد، اگرچه گلوکوکورتیکوئیدها به خودی خود واکنش ایجاد نمیکنند. این اثر را عمل مجاز آنها مینامند. اثرات مجاز شامل الزام وجود گلوکوکورتیکوئیدها برای گلوکاگون و کاتکول آمینها برای اعمال اثرات کالری زایی خود (نگاه کنید به بالا و فصل 24)، برای کاتکول آمینها برای اعمال اثرات لیپولیتیک خود، و برای کاتکول آمینها برای ایجاد پاسخهای فشار دهنده و اتساع برونش است.
EFFECTS ON ACTH SECRETION
Glucocorticoids inhibit ACTH secretion, which represents a negative feedback response on the pituitary. ACTH secretion is increased in adrenalectomized animals. The consequences of the negative feedback action of cortisol on ACTH secretion are discussed below in the section on regulation of glucocorticoid secretion.
اثرات بر ترشح ACTH
گلوکوکورتیکوئیدها ترشح ACTH را مهار میکنند که نشان دهنده یک پاسخ بازخورد منفی در هیپوفیز است. ترشح ACTH در حیوانات آدرنالکتومیافزایش مییابد. پیامدهای اثر بازخورد منفی کورتیزول بر ترشح ACTH در زیر در بخش تنظیم ترشح گلوکوکورتیکوئید مورد بحث قرار گرفته است.
VASCULAR REACTIVITY
In adrenally insufficient animals, vascular smooth muscle becomes unresponsive to norepinephrine and epinephrine. The capillaries dilate and, terminally, become permeable to colloidal dyes. Failure to respond to the norepinephrine liberated at noradrenergic nerve endings probably impairs vascular compensation for the hypovolemia of adrenal insufficiency and promotes vascular collapse. Glucocorticoids restore vascular reactivity.
واکنش عروقی
در حیواناتی که غدد فوق کلیوی کافی ندارند، عضله صاف عروق به نوراپی نفرین و اپی نفرین پاسخ نمیدهد. مویرگها گشاد میشوند و در نهایت به رنگهای کلوئیدی نفوذپذیر میشوند. پاسخ ندادن به نوراپی نفرین آزاد شده در انتهای عصب نورآدرنرژیک احتمالاً جبران عروقی برای هیپوولمینارسایی آدرنال را مختل میکند و باعث فروپاشی عروقی میشود. گلوکوکورتیکوئیدها واکنش پذیری عروق را بازیابی میکنند.
EFFECTS ON THE NERVOUS SYSTEM
Changes in the nervous system in adrenal insufficiency that are reversed only by glucocorticoids include the appearance of electroencephalographic waves slower than the normal ẞ rhythm, and personality changes. The latter, which are mild, include irritability, apprehension, and inability to concentrate.
اثرات بر روی سیستم عصبی
تغییرات سیستم عصبی در نارسایی آدرنال که فقط توسط گلوکوکورتیکوئیدها معکوس میشود شامل ظهور امواج الکتروانسفالوگرافی کندتر از ریتم طبیعی و تغییرات شخصیتی است. حالت دوم که خفیف است شامل تحریک پذیری، دلهره و ناتوانی در تمرکز است.
EFFECTS ON WATER METABOLISM
Adrenal insufficiency is characterized by an inability to excrete a water load, causing the possibility of water intoxication. Only glucocorticoids repair this deficit. In patients with adrenal insufficiency who have not received glucocorticoids, glucose infusion may cause high fever (“glucose fever”) followed by collapse and death. Presumably, the glucose is metabolized, the water dilutes the plasma, and the resultant osmotic gradient between the plasma and the cells causes the cells of the thermoregulatory centers in the hypothalamus to swell to such an extent that their function is disrupted.
اثرات بر متابولیسم آب
نارسایی آدرنال با ناتوانی در دفع آب مشخص میشود که باعث ایجاد مسمومیت با آب میشود. فقط گلوکوکورتیکوئیدها این کمبود را ترمیم میکنند. در بیماران مبتلا به نارسایی آدرنال که گلوکوکورتیکوئید دریافت نکرده اند، انفوزیون گلوکز ممکن است باعث تب بالا (“تب گلوکز”) و به دنبال آن فروپاشی و مرگ شود. احتمالاً گلوکز متابولیزه میشود، آب پلاسما را رقیق میکند و شیب اسمزی حاصل بین پلاسما و سلولها باعث میشود که سلولهای مراکز تنظیم کننده حرارت در هیپوتالاموس به حدی متورم شوند که عملکرد آنها مختل شود.
The cause of defective water excretion in adrenal insufficiency is unsettled. Plasma vasopressin levels are elevated in adrenal insufficiency and reduced by glucocorticoid treatment. The glomerular filtration rate is low, and this probably contributes to the reduction in water excretion. The selective effect of glucocorticoids on the abnormal water excretion is consistent with this possibility, because even though the mineralocorticoids improve filtration by restoring plasma volume, the glucocorticoids raise the glomerular filtration rate to a much greater degree.
علت دفع معیوب آب در نارسایی آدرنال مشخص نیست. سطح وازوپرسین پلاسما در نارسایی آدرنال افزایش یافته و با درمان گلوکوکورتیکوئید کاهش مییابد. سرعت فیلتراسیون گلومرولی کم است و این احتمالاً به کاهش دفع آب کمک میکند. اثر انتخابی گلوکوکورتیکوئیدها بر دفع غیرطبیعی آب با این احتمال سازگار است، زیرا حتی اگر مینرالوکورتیکوئیدها با بازگرداندن حجم پلاسما، فیلتراسیون را بهبود میبخشند، گلوکوکورتیکوئیدها میزان فیلتراسیون گلومرولی را به میزان بسیار بیشتری افزایش میدهند.
EFFECTS ON THE BLOOD CELLS & LYMPHATIC ORGANS
Glucocorticoids decrease the number of circulating eosinophils by increasing their sequestration in the spleen and lungs. Glucocorticoids also lower the number of basophils in the circulation and increase the number of neutrophils, platelets, and red blood cells (Table 19-4).
اثرات بر سلولهای خونی و اندامهای لنفاوی
گلوکوکورتیکوئیدها تعداد ائوزینوفیلهای در گردش را با افزایش تجمع آنها در طحال و ریهها کاهش میدهند. گلوکوکورتیکوئیدها همچنین تعداد بازوفیلها را در گردش خون کاهش میدهند و تعداد نوتروفیلها، پلاکتها و گلبولهای قرمز خون را افزایش میدهند (جدول 19-4).
TABLE 19-4 Typical effects of cortisol on the white and red blood cell counts in humans (cells/μL).
جدول 19-4 اثرات معمول کورتیزول بر تعداد گلبولهای سفید و قرمز خون در انسان (سلولها/μL).

Glucocorticoids decrease the circulating lymphocyte count and the size of the lymph nodes and thymus by inhibiting lymphocyte mitotic activity. They reduce secretion of cytokines by inhibiting the effect of NF-κB on the nucleus. The reduced secretion of the cytokine IL-2 leads to reduced proliferation of lymphocytes (see Chapter 3), and these cells undergo apoptosis.
گلوکوکورتیکوئیدها تعداد لنفوسیتهای در گردش و اندازه غدد لنفاوی و تیموس را با مهار فعالیت میتوز لنفوسیت کاهش میدهند. آنها با مهار اثر NF-kB بر روی هسته، ترشح سیتوکینها را کاهش میدهند. کاهش ترشح سیتوکین IL-2 منجر به کاهش تکثیر لنفوسیتها میشود (به فصل 3 مراجعه کنید)، و این سلولها تحت آپوپتوز قرار میگیرند.
RESISTANCE TO STRESS
The term stress as used in biology has been defined as any change in the environment that changes or threatens to change an existing optimal steady state. Most, if not all, of these stresses activate processes at the molecular, cellular, or systemic level that tend to restore the previous state, that is, they are homeostatic reactions. Some, but not all, of the stresses stimulate ACTH secretion. The increase in ACTH secretion is essential for survival when the stress is severe. If animals are then hypophysectomized, or adrenalectomized but treated with maintenance doses of glucocorticoids, they die when exposed to the same stress.
مقاومت در برابر استرس
اصطلاح استرس همانطور که در زیست شناسی استفاده میشود به عنوان هر تغییری در محیط تعریف شده است که وضعیت پایدار بهینه موجود را تغییر میدهد یا تهدید میکند. بیشتر، اگر نه همه، این تنشها، فرآیندهایی را در سطح مولکولی، سلولی یا سیستمیک فعال میکنند که تمایل به بازگرداندن حالت قبلی دارند، یعنی واکنشهای هموستاتیک هستند. برخی از تنشها، اما نه همه، ترشح ACTH را تحریک میکنند. وقتی استرس شدید است، افزایش ترشح ACTH برای بقا ضروری است. اگر حیوانات سپس هیپوفیزیکتومییا آدرنالکتومیشوند، اما با دوزهای نگهدارنده گلوکوکورتیکوئیدها درمان شوند، زمانی که در معرض همان استرس قرار بگیرند میمیرند.
The reason an elevated circulating ACTH, and hence glucocorticoid level, is essential for resisting stress remains for the most part unknown. Most of the stressful stimuli that increase ACTH secretion also activate the sympathetic nervous system, and part of the function of circulating glucocorticoids may be maintenance of vascular reactivity to catecholamines. Glucocorticoids are also necessary for the catecholamines to exert their full FFA-mobilizing action, and the FFAs are an important emergency energy supply. However,sympathectomized animals tolerate a variety of stresses with relative impunity. Another theory holds that glucocorticoids prevent other stress-induced changes from becoming excessive. At present, all that can be said is that stress causes increases in plasma glucocorticoids to high “pharmacologic” levels that in the short run are life-saving.
دلیل اینکه افزایش ACTH در گردش خون و در نتیجه سطح گلوکوکورتیکوئید برای مقاومت در برابر استرس ضروری است تا حد زیادی ناشناخته باقی مانده است. بیشتر محرکهای استرس زا که ترشح ACTH را افزایش میدهند، سیستم عصبی سمپاتیک را نیز فعال میکنند و بخشی از عملکرد گلوکوکورتیکوئیدهای در گردش ممکن است حفظ واکنش عروقی به کاتکول آمینها باشد. گلوکوکورتیکوئیدها همچنین برای کاتکول آمینها لازم هستند تا فعالیت کامل خود را برای بسیج FFA انجام دهند و FFAها یک منبع انرژی اضطراری مهم هستند. با این حال، حیوانات سمپاتیک شده انواع استرسها را با مصونیت نسبی تحمل میکنند. نظریه دیگری معتقد است که گلوکوکورتیکوئیدها از زیاد شدن سایر تغییرات ناشی از استرس جلوگیری میکنند. در حال حاضر، تنها چیزی که میتوان گفت این است که استرس باعث افزایش گلوکوکورتیکوئیدهای پلاسما به سطوح بالای “داروشناسی” میشود که در کوتاه مدت باعث نجات زندگی میشود.
It should also be noted that the increase in ACTH, which is beneficial in the short term, becomes harmful and disruptive in the long term, causing among other things, the abnormalities of Cushing syndrome.
همچنین باید توجه داشت که افزایش ACTH که در کوتاه مدت مفید است، در درازمدت مضر و مخرب میشود و در کنار سایر موارد باعث ایجاد ناهنجاریهای سندرم کوشینگ میشود.
PHARMACOLOGIC & PATHOLOGIC EFFECTS OF GLUCOCORTICOIDS
CUSHING SYNDROME
The clinical picture produced by prolonged increases in plasma glucocorticoids was described by Harvey Cushing and is called Cushing syndrome (Figure 19- 13). It may be ACTH-independent or ACTH-dependent. The causes of ACTH-independent Cushing syndrome include glucocorticoid-secreting adrenal tumors, adrenal hyperplasia, and prolonged administration of exogenous glucocorticoids for diseases such as rheumatoid arthritis. Rare but interesting ACTH-independent cases have been reported in which adrenocortical cells abnormally express receptors for gastric inhibitory polypeptide (GIP) (see Chapter 25), vasopressin (see Chapter 38), ẞ-adrenergic agonists, IL-1, or gonadotropin-releasing hormone (GnRH; see Chapter 22), causing these peptides to increase glucocorticoid secretion. The causes of ACTH-dependent Cushing syndrome include ACTH-secreting tumors of the anterior pituitary gland and tumors of other organs, usually the lungs, that secrete ACTH (ectopic ACTH syndrome) or corticotropin releasing hormone (CRH). Cushing syndrome due to anterior pituitary tumors is often called Cushing disease because these tumors were the cause of the cases described by Cushing. However, it is confusing to speak of Cushing disease as a subtype of Cushing syndrome, and the distinction seems to be of little more than historical value.
اثرات فارماکولوژیک و پاتولوژیک گلوکوکورتیکوئیدها
سندرم کوشینگ
تصویر بالینی تولید شده در اثر افزایش طولانی مدت گلوکوکورتیکوئیدهای پلاسما توسطهاروی کوشینگ توصیف شد و سندرم کوشینگ نامیده میشود (شکل 19-13). ممکن است مستقل از ACTH یا وابسته به ACTH باشد. علل سندرم کوشینگ مستقل از ACTH شامل تومورهای آدرنال ترشح کننده گلوکوکورتیکوئید، هیپرپلازی آدرنال و تجویز طولانی مدت گلوکوکورتیکوئیدهای اگزوژن برای بیماریهایی مانند آرتریت روماتوئید است. موارد نادر اما جالب مستقل از ACTH گزارش شده است که در آن سلولهای قشر آدرنال به طور غیرطبیعی گیرندههای پلی پپتید مهارکننده معده (GIP) (به فصل 25)، وازوپرسین (به فصل 38 مراجعه کنید)، آگونیستهای آدرنرژیک، IL-1، یا گنادوتروپین-کاهرون (کاهنده گنادوتروپین-relegering)، Chanadotropin-RelegasR (نگاه کنید به GIP) بیان میکنند. این پپتیدها ترشح گلوکوکورتیکوئید را افزایش میدهند. علل سندرم کوشینگ وابسته به ACTH عبارتند از تومورهای ترشح کننده ACTH در غده هیپوفیز قدامیو تومورهای سایر اندامها، معمولاً ریهها، که ACTH (سندرم ACTH نابجا) یا هورمون آزاد کننده کورتیکوتروپین (CRH) ترشح میکنند. سندرم کوشینگ ناشی از تومورهای هیپوفیز قدامیاغلب بیماری کوشینگ نامیده میشود زیرا این تومورها علت موارد توصیف شده توسط کوشینگ بودند. با این حال، صحبت از بیماری کوشینگ به عنوان یک زیرشاخه از سندرم کوشینگ گیج کننده است و به نظر میرسد این تمایز کمیبیشتر از ارزش تاریخی باشد.

FIGURE 19-13 Typical findings in Cushing syndrome. (Reproduced with permission from Forsham PH, Di Raimondo VC: Traumatic Medicine and Surgery for the Attorney. Butterworth; 1960.)
شکل 19-13 یافتههای معمول در سندرم کوشینگ. (تکثیر شده با مجوز Forsham PH، Di Raimondo VC: Traumatic Medicine and Surgery for the Attorney. Butterworth؛ 1960.)
Patients with Cushing syndrome are protein-depleted as a result of excess protein catabolism. The skin and subcutaneous tissues are therefore thin and the muscles are poorly developed. Wounds heal poorly, and minor injuries cause bruises and ecchymoses. The hair is thin and scraggly. Many patients with the disease have some increase in facial hair and acne, but this is caused by the increased secretion of adrenal androgens and often accompanies the increase in glucocorticoid secretion.
بیماران مبتلا به سندرم کوشینگ در نتیجه کاتابولیسم بیش از حد پروتئین دچار کمبود پروتئین میشوند. بنابراین پوست و بافتهای زیر جلدی نازک هستند و ماهیچهها رشد ضعیفی دارند. زخمها به خوبی بهبود نمییابند و آسیبهای جزئی باعث کبودی و اکیموز میشود. موها نازک و نازک است. بسیاری از بیماران مبتلا به این بیماری مقداری افزایش موهای صورت و آکنه دارند، اما این امر به دلیل افزایش ترشح آندروژنهای آدرنال ایجاد میشود و اغلب با افزایش ترشح گلوکوکورتیکوئید همراه است.
Body fat is redistributed in a characteristic way. The extremities are thin, but fat collects in the abdominal wall, face, and upper back, where it produces a “buffalo hump.” As the thin skin of the abdomen is stretched by the increased subcutaneous fat depots, the subdermal tissues rupture to form prominent reddish purple striae. These scars are seen normally whenever a rapid stretching of skin occurs, but in normal individuals the striae are usually inconspicuous and lack the intense purplish color.
چربی بدن به روشی مشخص دوباره توزیع میشود. اندامها نازک هستند، اما چربی در دیواره شکم، صورت و قسمت فوقانی پشت جمع میشود و در آنجا یک «قوز بوفالو» ایجاد میکند. همانطور که پوست نازک شکم توسط افزایش ذخایر چربی زیر جلدی کشیده میشود، بافتهای زیر پوستی پاره میشوند و خطوط بنفش مایل به قرمز برجسته را تشکیل میدهند. این اسکارها معمولاً هر زمان که کشش سریع پوست اتفاق میافتد دیده میشوند، اما در افراد عادی استریاها معمولاً مشخص نیستند و رنگ مایل به ارغوانی ندارند.
Many of the amino acids liberated from catabolized proteins are converted into glucose in the liver and the resultant hyperglycemia and decreased peripheral utilization of glucose may be sufficient to precipitate insulin-resistant diabetes mellitus, especially in patients genetically predisposed to diabetes. Hyperlipidemia and ketosis are associated with the diabetes, but acidosis is usually not severe.
بسیاری از اسیدهای آمینه آزاد شده از پروتئینهای کاتابولیزه شده در کبد به گلوکز تبدیل میشوند و هیپرگلیسمیناشی از آن و کاهش استفاده محیطی از گلوکز ممکن است برای ایجاد دیابت قندی مقاوم به انسولین، به ویژه در بیمارانی که از نظر ژنتیکی مستعد ابتلا به دیابت هستند، کافی باشد. هیپرلیپیدمیو کتوز با دیابت مرتبط هستند، اما اسیدوز معمولاً شدید نیست.
The glucocorticoids are present in such large amounts in Cushing syndrome that they may exert a significant mineralocorticoid action. Deoxycorticosterone secretion is also elevated in cases due to ACTH hypersecretion. The salt and water retention plus the facial obesity cause the characteristic plethoric, rounded “moon-faced” appearance, and there may be significant K* depletion and weakness. About 85% of patients with Cushing syndrome are hypertensive. The hypertension may be due to increased deoxycorticosterone secretion, increased angiotensinogen secretion, or a direct glucocorticoid effect on blood vessels (see Chapter 32).
گلوکوکورتیکوئیدها در مقادیر زیادی در سندرم کوشینگ وجود دارند که ممکن است اثر مینرالوکورتیکوئیدی قابل توجهی داشته باشند. ترشح دئوکسی کورتیکوسترون نیز در مواردی به دلیل ترشح بیش از حد ACTH افزایش مییابد. احتباس نمک و آب به همراه چاقی صورت باعث ایجاد ظاهری گرد و پربار مشخص میشود و ممکن است کاهش و ضعف قابل توجهی K* وجود داشته باشد. حدود 85 درصد از بیماران مبتلا به سندرم کوشینگ فشار خون بالا هستند. فشار خون بالا ممکن است به دلیل افزایش ترشح دئوکسی کورتیکوسترون، افزایش ترشح آنژیوتانسینوژن یا اثر مستقیم گلوکوکورتیکوئیدی بر عروق خونی باشد (به فصل 32 مراجعه کنید).
Glucocorticoid excess leads to bone dissolution by decreasing bone formation and increasing bone resorption. This leads to osteoporosis, a loss of bone mass that leads eventually to collapse of vertebral bodies and other fractures. The mechanisms by which glucocorticoids produce their effects on bone are discussed in Chapter 21.
گلوکوکورتیکوئید اضافی با کاهش تشکیل استخوان و افزایش جذب استخوان منجر به انحلال استخوان میشود. این منجر به پوکی استخوان میشود، از دست دادن توده استخوانی که در نهایت منجر به فروپاشی بدنه مهرهها و سایر شکستگیها میشود. مکانیسمهایی که توسط آن گلوکوکورتیکوئیدها اثرات خود را بر استخوان ایجاد میکنند در فصل 21 مورد بحث قرار میگیرند.
Glucocorticoids in excess accelerate the basic electroencephalographic rhythms and produce mental aberrations ranging from increased appetite, insomnia, and euphoria to frank toxic psychoses. As noted above, glucocorticoid deficiency is also associated with mental symptoms, but the symptoms produced by glucocorticoid excess are more severe.
گلوکوکورتیکوئیدها بیش از حد، ریتمهای اولیه الکتروانسفالوگرافی را تسریع میکنند و انحرافات ذهنی از افزایش اشتها، بی خوابی، و سرخوشی تا روان پریشیهای سمیرا ایجاد میکنند. همانطور که در بالا ذکر شد، کمبود گلوکوکورتیکوئید نیز با علائم ذهنی همراه است، اما علائم ناشی از افزایش گلوکوکورتیکوئید شدیدتر است.
ANTI-INFLAMMATORY & ANTI-ALLERGIC EFFECTS OF GLUCOCORTICOIDS
Glucocorticoids inhibit the inflammatory response to tissue injury. The glucocorticoids also suppress manifestations of allergic disease that are due to the release of histamine from mast cells and basophils. Both of these effects require high levels of circulating glucocorticoids and cannot be produced by administering steroids without producing the other manifestations of glucocorticoid excess. Furthermore, large doses of exogenous glucocorticoids inhibit ACTH secretion to the point that severe adrenal insufficiency can be a dangerous problem when therapy is stopped. However, local administration of glucocorticoids, for example, by injection into an inflamed joint or near an irritated nerve, produces a high local concentration of the steroid, often without enough systemic absorption to cause serious side effects.
اثرات ضد التهابی و ضد آلرژی گلوکوکورتیکوئیدها
گلوکوکورتیکوئیدها پاسخ التهابی به آسیب بافتی را مهار میکنند. گلوکوکورتیکوئیدها همچنین تظاهرات بیماریهای آلرژیک را که به دلیل آزاد شدن هیستامین از ماست سلها و بازوفیلها است، سرکوب میکنند. هر دوی این اثرات به سطوح بالایی از گلوکوکورتیکوئیدهای در گردش نیاز دارند و با تجویز استروئیدها بدون ایجاد سایر تظاهرات اضافی گلوکوکورتیکوئید قابل تولید نیستند. علاوه بر این، دوزهای زیادی از گلوکوکورتیکوئیدهای اگزوژن ترشح ACTH را تا حدی مهار میکنند که نارسایی شدید آدرنال میتواند یک مشکل خطرناک در صورت توقف درمان باشد. با این حال، تجویز موضعی گلوکوکورتیکوئیدها، به عنوان مثال، با تزریق در مفصل ملتهب یا نزدیک یک عصب تحریک شده، غلظت موضعی بالایی از استروئید ایجاد میکند، اغلب بدون جذب سیستمیک کافی برای ایجاد عوارض جانبی جدی.
The actions of glucocorticoids in patients with bacterial infections are dramatic but dangerous. For example, in pneumococcal pneumonia or active tuberculosis, the febrile reaction, the toxicity, and the lung symptoms disappear, but unless antibiotics are given at the same time, the bacteria spread throughout the body. It is important to remember that the symptoms are the warning that disease is present; when these symptoms are masked by treatment with glucocorticoids, there may be serious and even fatal delays in diagnosis and the institution of treatment with antimicrobial drugs.
عملکرد گلوکوکورتیکوئیدها در بیماران مبتلا به عفونتهای باکتریایی چشمگیر اما خطرناک است. به عنوان مثال، در پنومونی پنوموکوکی یا سل فعال، واکنش تب، سمیت و علائم ریوی از بین میرود، اما اگر همزمان آنتی بیوتیک داده نشود، باکتری در سراسر بدن پخش میشود. مهم است که به یاد داشته باشید که علائم هشدار دهنده وجود بیماری هستند. هنگامیکه این علائم با درمان با گلوکوکورتیکوئیدها پوشانده میشود، ممکن است تاخیرهای جدی و حتی کشنده در تشخیص و شروع درمان با داروهای ضد میکروبی ایجاد شود.
The role of NF-κB in the anti-inflammatory and anti-allergic effects of glucocorticoids has been mentioned above and is discussed in Chapter 3. An additional action that combats local inflammation is inhibition of phospholipase A2. This reduces the release of arachidonic acid from tissue phospholipids and consequently reduces the formation of leukotrienes, thromboxanes, prostaglandins, and prostacyclin (see Chapter 32).
نقش NF-kB در اثرات ضد التهابی و ضد حساسیت گلوکوکورتیکوئیدها در بالا ذکر شد و در فصل 3 مورد بحث قرار گرفت. یک اقدام اضافی که با التهاب موضعی مبارزه میکند، مهار فسفولیپاز A2 است. این باعث کاهش آزاد شدن اسید آراشیدونیک از فسفولیپیدهای بافتی و در نتیجه کاهش تشکیل لکوترینها، ترومبوکسانها، پروستاگلاندینها و پروستاسیکلین میشود (به فصل 32 مراجعه کنید).
OTHER EFFECTS
Large doses of glucocorticoids inhibit growth, decrease growth hormone secretion (see Chapter 18), induce PNMT, and decrease thyroid-stimulating hormone secretion. During fetal life, glucocorticoids accelerate the maturation of surfactant in the lungs (see Chapter 34).
سایر اثرات
دوزهای زیاد گلوکوکورتیکوئیدها رشد را مهار میکنند، ترشح هورمون رشد را کاهش میدهند (به فصل 18 مراجعه کنید)، باعث القای PNMT و کاهش ترشح هورمون محرک تیروئید میشوند. در طول زندگی جنینی، گلوکوکورتیکوئیدها بلوغ سورفکتانت را در ریهها تسریع میکنند (به فصل 34 مراجعه کنید).
REGULATION OF GLUCOCORTICOID SECRETION
ROLE OF ACTH
Both basal secretion of glucocorticoids and the increased secretion provoked by stress depend on ACTH from the anterior pituitary (Chapter 18). Angiotensin II also stimulates the adrenal cortex, but its effect is mainly on aldosterone secretion. Large doses of a number of other naturally occurring substances, including vasopressin, serotonin, and vasoactive intestinal polypeptide (VIP), are capable of stimulating the adrenal directly, but there is no evidence that these agents play any role in the physiologic regulation of glucocorticoid secretion.
تنظیم ترشح گلوکوکورتیکوئید
نقش ACTH
هم ترشح پایه گلوکوکورتیکوئیدها و هم افزایش ترشح ناشی از استرس به ACTH از هیپوفیز قدامیبستگی دارد (فصل 18). آنژیوتانسین II همچنین قشر آدرنال را تحریک میکند، اما تأثیر آن عمدتاً بر ترشح آلدوسترون است. دوزهای زیاد تعدادی از مواد طبیعی دیگر، از جمله وازوپرسین، سروتونین، و پلی پپتید وازواکتیو روده ای (VIP)، قادر به تحریک مستقیم آدرنال هستند، اما هیچ مدرکی مبنی بر اینکه این عوامل نقشی در تنظیم فیزیولوژیک ترشح گلوکوکورتیکوئید ایفا میکنند وجود ندارد.
CHEMISTRY & METABOLISM OF ACTH
ACTH is a single-chain polypeptide containing 39 amino acids. Its origin from proopiomelanocortin (POMC) in the pituitary is discussed in Chapter 18. The first 23 amino acids in the chain generally constitute the active “core” of the molecule. Amino acids 24-39 constitute a “tail” that stabilizes the molecule and varies slightly in composition from species to species. The ACTHs that have been isolated are generally active in all species but antigenic in heterologous species.
شیمیو متابولیسم ACTH
ACTH یک پلی پپتید تک زنجیره ای است که حاوی 39 اسید آمینه است. منشا آن از proopiomelanocortin (POMC) در هیپوفیز در فصل 18 مورد بحث قرار گرفته است. 23 اسید آمینه اول در زنجیره به طور کلی “هسته” فعال مولکول را تشکیل میدهند. اسیدهای آمینه 24-39 یک “دم” را تشکیل میدهند که مولکول را تثبیت میکند و از گونه ای به گونه دیگر در ترکیب کمیمتفاوت است. ACTHهایی که جدا شده اند به طور کلی در همه گونهها فعال هستند اما در گونههای هترولوگ آنتی ژن هستند.
ACTH is inactivated in blood in vitro more slowly than in vivo; its half-life in the circulation in humans is about 10 min. A large part of an injected dose of ACTH is found in the kidneys, but neither nephrectomy nor evisceration appreciably enhances its in vivo activity, and the site of its inactivation is not known.
ACTH در خون در شرایط آزمایشگاهی کندتر از in vivo غیر فعال میشود. نیمه عمر آن در گردش خون در انسان حدود 10 دقیقه است. بخش بزرگی از دوز تزریقی ACTH در کلیهها یافت میشود، اما نه نفرکتومیو نه بیرون زدگی به طور قابل ملاحظه ای فعالیت آن را در داخل بدن افزایش نمیدهند و محل غیرفعال شدن آن مشخص نیست.
EFFECT OF ACTH ON THE ADRENAL
After hypophysectomy, glucocorticoid synthesis and output decline within 1 h to very low levels, although some hormone is still secreted. Within a short time after an injection of ACTH (in dogs, less than 2 min), glucocorticoid output is increased. With low doses of ACTH, the relationship between the log of the dose and the increase in glucocorticoid secretion is linear. However, the maximal rate at which glucocorticoids can be secreted is rapidly reached, and this “ceiling on output” also exists in humans. The effects of ACTH on adrenal morphology and the mechanism by which it increases steroid secretion have been discussed above.
اثر ACTH بر غده فوق کلیوی
پس از هیپوفیزکتومی، سنتز گلوکوکورتیکوئید و برون ده در عرض 1 ساعت به سطوح بسیار پایین کاهش مییابد، اگرچه مقداری هورمون هنوز ترشح میشود. در مدت کوتاهی پس از تزریق ACTH (در سگها، کمتر از 2 دقیقه)، برون ده گلوکوکورتیکوئید افزایش مییابد. با دوزهای پایین ACTH، رابطه بین لاگ دوز و افزایش ترشح گلوکوکورتیکوئید خطی است. با این حال، حداکثر سرعت ترشح گلوکوکورتیکوئیدها به سرعت به دست میآید و این “سقف خروجی” در انسان نیز وجود دارد. اثرات ACTH بر مورفولوژی آدرنال و مکانیسم افزایش ترشح استروئید در بالا مورد بحث قرار گرفته است.
ADRENAL RESPONSIVENESS
ACTH not only produces prompt increases in glucocorticoid secretion but also increases the sensitivity of the adrenal to subsequent doses of ACTH. Conversely, single doses of ACTH do not increase glucocorticoid secretion in chronically hypophysectomized animals and patients with hypopituitarism, and repeated injections or prolonged infusions of ACTH are necessary to restore normal adrenal responses to ACTH. Decreased responsiveness is also produced by doses of glucocorticoids that inhibit ACTH secretion. The decreased adrenal responsiveness to ACTH is detectable within 24 h after hypophysectomy and increases progressively with time (Figure 19-14). It is marked when the adrenal is atrophic but develops before visible changes occur in adrenal size or morphology.
پاسخ آدرنال
ACTH نه تنها باعث افزایش سریع ترشح گلوکوکورتیکوئید میشود، بلکه حساسیت غده فوق کلیوی به دوزهای بعدی ACTH را نیز افزایش میدهد. برعکس، دوزهای منفرد ACTH باعث افزایش ترشح گلوکوکورتیکوئید در حیوانات هیپوفیزکتومیمزمن و بیماران مبتلا به هیپوفیز نمیشود و تزریق مکرر یا انفوزیون طولانی مدت ACTH برای بازگرداندن پاسخ طبیعی آدرنال به ACTH ضروری است. کاهش پاسخ دهی نیز با دوزهای گلوکوکورتیکوئیدی که ترشح ACTH را مهار میکنند ایجاد میشود. کاهش پاسخ آدرنال به ACTH ظرف 24 ساعت پس از هیپوفیسکتومیقابل تشخیص است و به تدریج با گذشت زمان افزایش مییابد (شکل 19-14). زمانی مشخص میشود که آدرنال آتروفیک باشد اما قبل از اینکه تغییرات قابل مشاهده در اندازه یا مورفولوژی آدرنال رخ دهد، ایجاد میشود.

FIGURE 19–14 Loss of ACTH responsiveness when ACTH secretion is decreased in humans. The 1- to 24-amino-acid sequence of ACTH was infused intravenously (IV) in a dose of 250 μg over 8 h. CST, long-term corticosteroid therapy; DX, dexamethasone 0.75 mg every 8 h for 3 days; API, anterior pituitary insufficiency; N, normal subjects. (Reproduced with permission from Kolanowski J, et al: Adrenocortical response upon repeated stimulation with corticotropin in patients lacking endogenous corticotropin secretion. Acta Endocrinol [Kbh] 1977;85:595.)
شکل 19-14 از دست دادن پاسخ ACTH زمانی که ترشح ACTH در انسان کاهش مییابد. توالی 1 تا 24 آمینو اسیدی ACTH به صورت داخل وریدی (IV) در دوز 250 میکروگرم در طی 8 ساعت تزریق شد. CST، درمان طولانی مدت کورتیکواستروئید؛ DX، دگزامتازون 0.75 میلی گرم هر 8 ساعت به مدت 3 روز. API، نارسایی هیپوفیز قدامی؛ N، افراد عادی. (بازتولید شده با اجازه Kolanowski J، و همکاران: پاسخ قشر آدرنال بر تحریک مکرر با کورتیکوتروپین در بیماران فاقد ترشح درون زا کورتیکوتروپین. Acta Endocrinol [Kbh] 1977؛ 85:595.)
CIRCADIAN RHYTHM
ACTH is secreted in irregular bursts throughout the day and plasma cortisol tends to rise and fall in response to these bursts (Figure 19-15). In humans, the bursts are most frequent in the early morning, and about 75% of the daily production of cortisol occurs between 4:00 AM and 10:00 AM. The bursts are least frequent in the evening. This diurnal (circadian) rhythm in ACTH secretion is present in patients with adrenal insufficiency receiving constant doses of glucocorticoids. It is not due to the stress of getting up in the morning, traumatic as that may be, because the increased ACTH secretion occurs before waking up. If the “day” is lengthened experimentally to more than 24 h, that is, if the individual is isolated and the day’s activities are spread over more than 24h, the adrenal cycle also lengthens, but the increase in ACTH secretion still occurs during the period of sleep. The biologic clock responsible for the diurnal ACTH rhythm is located in the suprachiasmatic nuclei of the hypothalamus (see Chapter 17).
ریتم شبانه روزی
ACTH به صورت انفجارهای نامنظم در طول روز ترشح میشود و کورتیزول پلاسما تمایل به افزایش و کاهش در پاسخ به این انفجارها دارد (شکل 19-15). در انسان، انفجارها در اوایل صبح بیشتر اتفاق میافتد و حدود 75 درصد از تولید روزانه کورتیزول بین ساعت 4 صبح تا 10 صبح رخ میدهد. انفجارها در عصر کمترین فراوانی را دارند. این ریتم روزانه (سیرکادین) در ترشح ACTH در بیماران مبتلا به نارسایی آدرنال که دوزهای ثابت گلوکوکورتیکوئید دریافت میکنند وجود دارد. این به دلیل استرس ناشی از بیدار شدن در صبح نیست، حتی ممکن است آسیب زا باشد، زیرا افزایش ترشح ACTH قبل از بیدار شدن از خواب رخ میدهد. اگر “روز” به طور تجربی به بیش از 24 ساعت طولانی شود، یعنی اگر فرد منزوی شود و فعالیتهای روز بیش از 24 ساعت پخش شود، چرخه آدرنال نیز طولانی تر میشود، اما افزایش ترشح ACTH همچنان در طول دوره خواب رخ میدهد. ساعت بیولوژیکی مسئول ریتم روزانه ACTH در هستههای فوق کیاسماتیک هیپوتالاموس قرار دارد (به فصل 17 مراجعه کنید).

FIGURE 19-15 Fluctuations in plasma ACTH and glucocorticoids throughout the day in a normal girl (age 16). The ACTH was measured by immunoassay and the glucocorticoids as 11-oxysteroids (11-OHCS). Note the greater ACTH and glucocorticoid rises in the morning, before awakening. ACTH, adrenocorticotropic hormone. (Reproduced, with permission, from Krieger DT, et al: Characterization of the normal temporal pattern of plasma corticosteroid levels. J Clin Endocrinol Metab 1971; Feb; 32(2):266–284.)
شکل 19-15 نوسانات ACTH پلاسما و گلوکوکورتیکوئیدها در طول روز در یک دختر عادی (سن 16 سالگی). ACTH با روش ایمونواسی و گلوکوکورتیکوئیدها به عنوان 11-اکسی استروئید (11-OHCS) اندازه گیری شد. توجه داشته باشید که ACTH و گلوکوکورتیکوئید بیشتر در صبح، قبل از بیدار شدن از خواب افزایش مییابد. ACTH، هورمون آدرنوکورتیکوتروپیک. (بازتولید شده، با اجازه، از کریگر DT، و همکاران: خصوصیات الگوی زمانی طبیعی سطوح کورتیکواستروئید پلاسما. J Clin Endocrinol Metab 1971؛ فوریه؛ 32(2):266-284.)
THE RESPONSE TO STRESS
The morning plasma ACTH concentration in a healthy resting human is about 25 pg/mL (5.5 pmol/L). ACTH and cortisol values in various abnormal conditions are summarized in Figure 19-16. During severe stress, the amount of ACTH secreted exceeds the amount necessary to produce maximal glucocorticoid output. However, prolonged exposure to ACTH in conditions such as the ectopic ACTH syndrome increases the adrenal maximum.
پاسخ به استرس
غلظت ACTH پلاسمایی صبحگاهی در یک انسان سالم در حال استراحت حدود 25 pg/mL (5.5 pmol/L) است. مقادیر ACTH و کورتیزول در شرایط غیر طبیعی مختلف در شکل 19-16 خلاصه شده است. در طول استرس شدید، مقدار ACTH ترشح شده از مقدار لازم برای تولید حداکثر خروجی گلوکوکورتیکوئید بیشتر است. با این حال، مواجهه طولانی مدت با ACTH در شرایطی مانند سندرم ACTH نابجا، حداکثر آدرنال را افزایش میدهد.
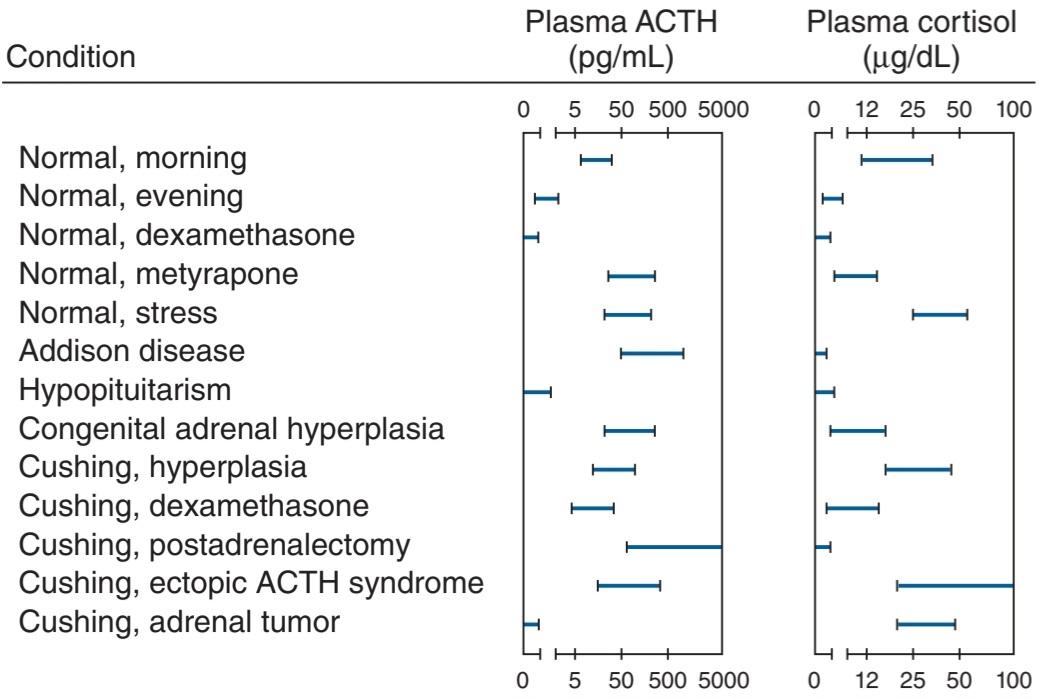
FIGURE 19-16 Plasma concentrations of ACTH and cortisol in various clinical states. ACTH, adrenocorticotropic hormone. (Reproduced with permission from Williams RH [editor]: Textbook of Endocrinology, 5th ed. Saunders, 1974.)
شکل 19-16 غلظت پلاسمایی ACTH و کورتیزول در حالات مختلف بالینی. ACTH، هورمون آدرنوکورتیکوتروپیک. (تکثیر شده با اجازه ویلیامز RH [ویرایشگر]: کتاب درسی غدد درون ریز، ویرایش پنجم ساندرز، 1974.)
Increases in ACTH secretion to meet emergency situations are mediated almost exclusively through the hypothalamus via release of CRH. This polypeptide is produced by neurons in the paraventricular nuclei. It is secreted in the median eminence and transported in the portal-hypophysial vessels to the anterior pituitary, where it stimulates ACTH secretion (see Chapter 18). If the median eminence is destroyed, increased secretion in response to many different stresses is blocked. Afferent nerve pathways from many parts of the brain converge on the paraventricular nuclei. Fibers from the amygdaloid nuclei mediate responses to emotional stresses, and fear, anxiety, and apprehension cause marked increases in ACTH secretion. Input from the suprachiasmatic nuclei provides the drive for the diurnal rhythm. Impulses ascending to the hypothalamus via the nociceptive pathways and the reticular formation trigger increased ACTH secretion in response to injury (Figure 19-16). The baroreceptors exert an inhibitory input via the nucleus of the tractus solitarius.
افزایش ترشح ACTH برای مواجهه با شرایط اضطراری تقریباً منحصراً از طریق هیپوتالاموس از طریق آزادسازی CRH انجام میشود. این پلی پپتید توسط سلولهای عصبی موجود در هستههای پارا بطنی تولید میشود. در برجستگی میانی ترشح میشود و در عروق پورتال-هیپوفیزال به هیپوفیز قدامیمنتقل میشود، جایی که ترشح ACTH را تحریک میکند (به فصل 18 مراجعه کنید). اگر برجستگی میانی از بین برود، افزایش ترشح در پاسخ به بسیاری از تنشهای مختلف مسدود میشود. مسیرهای عصبی آوران از بسیاری از قسمتهای مغز روی هستههای پارا بطنی همگرا میشوند. فیبرهای هستههای آمیگدال واکنشها به استرسهای عاطفی را واسطه میکنند و ترس، اضطراب و دلهره باعث افزایش قابل توجه ترشح ACTH میشوند. ورودی از هستههای فوق کیاسماتیک محرک ریتم روزانه را فراهم میکند. تکانههایی که از طریق مسیرهای درد به هیپوتالاموس صعود میکنند و تشکیل شبکه ای باعث افزایش ترشح ACTH در پاسخ به آسیب میشود (شکل 19-16). بارورسپتورها یک ورودی بازدارنده از طریق هسته tractus solitarius اعمال میکنند.
GLUCOCORTICOID FEEDBACK
Free glucocorticoids inhibit ACTH secretion, and the degree of pituitary inhibition is proportional to the circulating glucocorticoid level. The inhibitory effect is exerted at both the pituitary and the hypothalamic levels. The inhibition is due primarily to an action on DNA, and maximal inhibition takes several hours to develop, although more rapid “fast feedback” also occurs. The ACTH- inhibiting activity of the various steroids parallels their glucocorticoid potency. A drop in resting corticoid levels stimulates ACTH secretion, and in chronic adrenal insufficiency the rate of ACTH synthesis and secretion is markedly increased.
بازخورد گلوکوکورتیکوئید
گلوکوکورتیکوئیدهای آزاد ترشح ACTH را مهار میکنند و میزان مهار هیپوفیز متناسب با سطح گلوکوکورتیکوئید در گردش است. اثر مهاری در هر دو سطح هیپوفیز و هیپوتالاموس اعمال میشود. این مهار عمدتاً به دلیل یک عمل روی DNA است و حداکثر بازداری چندین ساعت طول میکشد تا ایجاد شود، اگرچه “بازخورد سریع” سریعتر نیز رخ میدهد. فعالیت مهارکننده ACTH استروئیدهای مختلف به موازات قدرت گلوکوکورتیکوئید آنها است. کاهش سطح کورتیکوئید در حالت استراحت باعث تحریک ترشح ACTH میشود و در نارسایی مزمن آدرنال سرعت سنتز و ترشح ACTH به طور قابل توجهی افزایش مییابد.
Thus, the rate of ACTH secretion is determined by two opposing forces: the sum of the neural and possibly other stimuli converging through the hypothalamus to increase ACTH secretion, and the magnitude of the braking action of glucocorticoids on ACTH secretion, which is proportional to their level in the circulating blood (Figure 19–17).
بنابراین، سرعت ترشح ACTH توسط دو نیروی متضاد تعیین میشود: مجموع محرکهای عصبی و احتمالاً سایر محرکهایی که از هیپوتالاموس برای افزایش ترشح ACTH همگرا میشوند، و میزان اثر ترمز گلوکوکورتیکوئیدها بر ترشح ACTH، که متناسب با سطح آنها در خون در گردش است (شکل 17-19).

FIGURE 19-17 Feedback control of the secretion of cortisol and other glucocorticoids via the hypothalamic-pituitary-adrenal axis. The dashed arrows indicate inhibitory effects and the solid arrows indicate stimulating effects. ACTH, adrenocorticotropic hormone; CRH, corticotropin-releasing hormone; NTS, nucleus tractus solitarius.
شکل 19-17 کنترل بازخورد ترشح کورتیزول و سایر گلوکوکورتیکوئیدها از طریق محور هیپوتالاموس-هیپوفیز-آدرنال. فلشهای چین دار نشان دهنده اثرات بازدارنده و فلشهای جامد نشان دهنده اثرات تحریک کننده است. ACTH، هورمون آدرنوکورتیکوتروپیک؛ CRH، هورمون آزاد کننده کورتیکوتروپین؛ NTS، Nucleus tractus Solitarius.
The dangers involved when prolonged treatment with anti-inflammatory doses of glucocorticoids is stopped deserve emphasis. Not only is the adrenal atrophic and unresponsive after such treatment, but even if its responsiveness is restored by injecting ACTH, the pituitary may be unable to secrete normal amounts of ACTH for as long as a month. The cause of the deficiency is presumably diminished ACTH synthesis. Thereafter, ACTH secretion slowly increases to supranormal levels. These in turn stimulate the adrenal, and glucocorticoid output rises, with feedback inhibition gradually reducing the elevated ACTH levels to normal (Figure 19-18). The complications of sudden cessation of steroid therapy can usually be avoided by slowly decreasing the steroid dose over a long period of time.
خطرات ناشی از توقف درمان طولانی مدت با دوزهای ضد التهابی گلوکوکورتیکوئیدها، شایسته تاکید است. نه تنها آدرنال آتروفیک است و پس از چنین درمانی پاسخ نمیدهد، بلکه حتی اگر پاسخگویی آن با تزریق ACTH بازیابی شود، هیپوفیز ممکن است تا یک ماه نتواند مقادیر طبیعی ACTH ترشح کند. علت کمبود احتمالاً کاهش سنتز ACTH است. پس از آن، ترشح ACTH به آرامیبه سطوح فوق طبیعی افزایش مییابد. اینها به نوبه خود غده فوق کلیوی را تحریک میکنند و برون ده گلوکوکورتیکوئید افزایش مییابد، با مهار بازخوردی که به تدریج سطوح ACTH بالا را به حالت طبیعی کاهش میدهد (شکل 19-18). عوارض قطع ناگهانی استروئید درمانی را معمولاً میتوان با کاهش آهسته دوز استروئید در یک دوره زمانی طولانی اجتناب کرد.

FIGURE 19-18 Pattern of plasma ACTH and cortisol values in patients recovering from prior long-term daily treatment with large doses of glucocorticoids. ACTH, adrenocorticotropic hormone. (Used with permission of R Ney.)
شکل 19-18 الگوی مقادیر ACTH و کورتیزول پلاسما در بیمارانی که از درمان روزانه طولانی مدت قبلی با دوزهای زیاد گلوکوکورتیکوئیدها بهبود مییابند. ACTH، هورمون آدرنوکورتیکوتروپیک. (با اجازه R Ney استفاده میشود.)
EFFECTS OF MINERALOCORTICOIDS
ACTIONS
Aldosterone and other steroids with mineralocorticoid activity increase the reabsorption of Na* from the urine, sweat, saliva, and the contents of the colon. Thus, mineralocorticoids cause retention of Na+ in the ECF. This expands ECF volume. In the kidneys, they act primarily on the principal cells (P cells) of the collecting ducts (see Chapter 37). Under the influence of aldosterone, increased amounts of Na+ are in effect exchanged for K* and H+ in the renal tubules, producing a K+ diuresis (Figure 19-19) and an increase in urine acidity.
اثرات مینرالوکورتیکوئیدها
اقدامات
آلدوسترون و سایر استروئیدها با فعالیت مینرالوکورتیکوئیدی باعث افزایش بازجذب +Na از ادرار، عرق، بزاق و محتویات روده بزرگ میشوند. بنابراین، مینرالوکورتیکوئیدها باعث حفظ +Na در ECF میشوند. این باعث افزایش حجم ECF میشود. در کلیهها، آنها عمدتاً روی سلولهای اصلی (سلولهای P) مجاری جمع کننده عمل میکنند (به فصل 37 مراجعه کنید). تحت تأثیر آلدوسترون، مقادیر افزایش یافته +Na در واقع با +K و +H در لولههای کلیوی مبادله میشود و باعث ایجاد دیورز +K میشود (شکل 19-19) و افزایش اسیدیته ادرار.
FIGURE 19-19 Effect of aldosterone (5 μg as a single dose injected into the aorta) on electrolyte excretion in an adrenalectomized dog. The scale for creatinine clearance is on the right.
شکل 19-19 اثر آلدوسترون (5 میکروگرم به عنوان یک دوز منفرد تزریق شده به آئورت) بر دفع الکترولیت در سگ آدرنالکتومیشده. مقیاس کلیرانس کراتینین در سمت راست است.
MECHANISM OF ACTION
Like many other steroids, aldosterone binds to a cytoplasmic receptor, and the receptor-hormone complex moves to the nucleus where it alters the transcription of mRNAs. This in turn increases the production of proteins that alter cell function. The aldosterone-stimulated proteins have two effects a rapid effect, to increase the activity of epithelial sodium channels (ENaCs) by increasing the insertion of these channels into the cell membrane from a cytoplasmic pool; and a slower effect to increase the synthesis of ENaCs. Among the genes activated by aldosterone is the gene for serum- and glucocorticoid-regulated kinase (sgk), a serine-threonine protein kinase. The gene for sgk is an early response gene, and sgk increases ENaC activity. Aldosterone also increases the mRNAs for the three subunits that make up ENaCs. The fact that sgk is activated by glucocorticoids as well as aldosterone is not a problem because glucocorticoids are inactivated at mineralocorticoid receptor sites. However, aldosterone activates the genes for other proteins in addition to sgk and ENaCs and inhibits others. Therefore, the exact mechanism by which aldosterone-induced proteins increase Na* reabsorption is still unsettled.
مکانیسم عمل
مانند بسیاری از استروئیدهای دیگر، آلدوسترون به یک گیرنده سیتوپلاسمیمتصل میشود و مجموعه گیرنده-هورمون به سمت هسته حرکت میکند که در آنجا رونویسی mRNAها را تغییر میدهد. این به نوبه خود باعث افزایش تولید پروتئینهایی میشود که عملکرد سلول را تغییر میدهند. پروتئینهای تحریک شده با آلدوسترون دو اثر دارند: یک اثر سریع، افزایش فعالیت کانالهای سدیم اپیتلیال (ENaCs) با افزایش ورود این کانالها به غشای سلولی از یک استخر سیتوپلاسمی. و یک اثر کندتر برای افزایش سنتز ENaCs. از جمله ژنهای فعال شده توسط آلدوسترون، ژن کیناز تنظیم شده با سرم و گلوکوکورتیکوئید (sgk)، یک پروتئین کیناز سرین ترئونین است. ژن sgk یک ژن پاسخ اولیه است و sgk فعالیت ENaC را افزایش میدهد. آلدوسترون همچنین mRNAها را برای سه زیرواحد تشکیل دهنده ENaC افزایش میدهد. این واقعیت که sgk توسط گلوکوکورتیکوئیدها و همچنین آلدوسترون فعال میشود مشکلی نیست زیرا گلوکوکورتیکوئیدها در مکانهای گیرنده مینرالوکورتیکوئید غیرفعال میشوند. با این حال، آلدوسترون علاوه بر sgk و ENaCs، ژنهای سایر پروتئینها را فعال میکند و سایر پروتئینها را مهار میکند. بنابراین، مکانیسم دقیقی که توسط آن پروتئینهای ناشی از آلدوسترون باعث افزایش بازجذب +Na میشود هنوز مشخص نیست.
Evidence is accumulating that aldosterone also binds to the cell membrane and by a rapid, nongenomic action increases the activity of membrane Na+-K+ exchangers. This produces an increase in intracellular Na+, and the second messenger involved is probably IP3. In any case, the principal effect of aldosterone on Na+ transport takes 10-30 min to develop and peaks even later (Figure 19-19), indicating that it depends on the synthesis of new proteins by a genomic mechanism.
شواهدی در حال انباشته شدن است مبنی بر اینکه آلدوسترون نیز به غشای سلولی متصل میشود و با یک عمل سریع و غیر ژنومیک، فعالیت مبدلهای +Na+-K غشا را افزایش میدهد. این باعث افزایش +Na داخل سلولی میشود و پیام رسان دوم احتمالاً IP3 است. در هر صورت، اثر اصلی آلدوسترون بر انتقال Na+ 10 تا 30 دقیقه طول میکشد تا ایجاد شود و حتی بعداً به اوج خود میرسد (شکل 19-19)، که نشان میدهد که بستگی به سنتز پروتئینهای جدید توسط یک مکانیسم ژنومیدارد.
RELATION OF MINERALOCORTICOID TO GLUCOCORTICOID RECEPTORS
It is intriguing that in vitro, the mineralocorticoid receptor (MR) has an appreciably higher affinity for glucocorticoids than the glucocorticoid receptor does, and glucocorticoids are present in large amounts in vivo. This raises the question of why glucocorticoids do not bind to the mineralocorticoid receptors in the kidneys and other locations and produce mineralocorticoid effects. At least in part, the answer is that the kidneys and other mineralocorticoid-sensitive tissues also contain the enzyme 11ẞ-hydroxysteroid dehydrogenase type 2. This enzyme leaves aldosterone untouched, but it converts cortisol to cortisone(Figure 19-11) and corticosterone to its 11-oxy derivative. These 11-oxy derivatives do not bind to the receptor (Clinical Box 19-3).
ارتباط مینرالوکورتیکوئید با گیرندههای گلوکوکورتیکوئید
جالب است که در شرایط آزمایشگاهی، گیرنده مینرالوکورتیکوئیدی (MR) نسبت به گیرنده گلوکوکورتیکوئید میل ترکیبی بسیار بالاتری برای گلوکوکورتیکوئیدها دارد و گلوکوکورتیکوئیدها در مقادیر زیادی در داخل بدن وجود دارند. این سوال را مطرح میکند که چرا گلوکوکورتیکوئیدها به گیرندههای مینرالوکورتیکوئیدی در کلیهها و سایر مکانها متصل نمیشوند و اثرات مینرالوکورتیکوئیدی ایجاد نمیکنند. حداقل تا حدودی، پاسخ این است که کلیهها و سایر بافتهای حساس به مینرالوکورتیکوئید نیز حاوی آنزیم 11-هیدروکسی استروئید دهیدروژناز نوع 2 هستند. این آنزیم آلدوسترون را دست نخورده باقی میگذارد، اما کورتیزول را به کورتیزون (شکل 19-11) و کورتیکوسترون را به مشتقات آن تبدیل میکند. این مشتقات 11-اکسی به گیرنده متصل نمیشوند (باکس 19-3).
CLINICAL BOX 19-3
Apparent Mineralocorticoid Excess
If 11ẞ-hydroxysteroid dehydrogenase type 2 is inhibited or absent, cortisol has marked mineralocorticoid effects. The resulting syndrome is called apparent mineralocorticoid excess (AME). Patients with this condition have the clinical picture of hyperaldosteronism because cortisol is acting on their mineralocorticoid receptors, and their plasma aldosterone level as well as their plasma renin activity is low. The condition can be due to congenital absence of the enzyme.
جعبه بالینی 19-3
افزایش ظاهری مینرالوکورتیکوئید
اگر 11-هیدروکسی استروئید دهیدروژناز نوع 2 مهار یا وجود نداشته باشد، کورتیزول دارای اثرات معدنی کورتیکوئیدی مشخص است. سندرم ایجاد شده، بیش از حد معدنی کورتیکوئید آشکار (AME) نامیده میشود. بیماران مبتلا به این بیماری تصویر بالینی هیپرآلدوسترونیسم را دارند زیرا کورتیزول روی گیرندههای معدنی کورتیکوئید آنها اثر میگذارد و سطح آلدوسترون پلاسما و همچنین فعالیت رنین پلاسما آنها پایین است. این وضعیت میتواند به دلیل عدم وجود آنزیم مادرزادی باشد.
THERAPEUTIC HIGHLIGHTS
Prolonged ingestion of licorice can also cause an increase in blood pressure. Outside of the United States, licorice contains glycyrrhetinic acid, which inhibits 11ẞ-hydroxysteroid dehydrogenase type 2. Individuals who eat large amounts of licorice have an increase in MR-activated sodium absorption via the epithelial sodium channel ENaC in the renal collecting duct, and blood pressure can rise.
نکات برجسته درمانی
مصرف طولانی مدت شیرین بیان همچنین میتواند باعث افزایش فشار خون شود. در خارج از ایالات متحده، شیرین بیان حاوی اسید گلیسیرتینیک است که 11-هیدروکسی استروئید دهیدروژناز نوع 2 را مهار میکند. افرادی که مقادیر زیادی شیرین بیان میخورند، جذب سدیم فعال شده با MR از طریق کانال سدیم اپیتلیال ENaC در مجرای جمع کننده کلیه افزایش مییابد و فشار خون میتواند افزایش یابد.
OTHER STEROIDS THAT AFFECT NA+ EXCRETION
Aldosterone is the principal mineralocorticoid secreted by the adrenals, although corticosterone is secreted in sufficient amounts to exert a minor mineralocorticoid effect (Tables 19-1 and 19-2). Deoxycorticosterone, which is secreted in appreciable amounts only in abnormal situations, has about 3% of the activity of aldosterone. Large amounts of progesterone and some other steroids cause natriuresis, but there is little evidence that they play any normal role in the control of Na* excretion.
سایر استروئیدهایی که بر دفع +Na تأثیر میگذارند
آلدوسترون مینرالوکورتیکوئید اصلی ترشح شده توسط آدرنال است، اگرچه کورتیکوسترون به مقدار کافی برای اعمال یک اثر مینرالوکورتیکوئیدی جزئی ترشح میشود (جدول 19-1 و 19-2). دئوکسی کورتیکوسترون که فقط در شرایط غیر طبیعی به مقدار قابل توجهی ترشح میشود، حدود 3 درصد از فعالیت آلدوسترون را دارد. مقادیر زیاد پروژسترون و برخی دیگر از استروئیدها باعث ناتریورز میشوند، اما شواهد کمیوجود دارد که نشان دهد آنها نقش طبیعی در کنترل دفع +Na دارند.
EFFECT OF ADRENALECTOMY
In adrenal insufficiency, Na* is lost in the urine; K* is retained, and the plasma K* rises. When adrenal insufficiency develops rapidly, the amount of Na* lost from the ECF exceeds the amount excreted in the urine, indicating that Na* also must be entering cells. When the posterior pituitary is intact, salt loss exceeds water loss, and the plasma Na+ falls (Table 19-5). However, the plasma volume is also reduced, resulting in hypotension, circulatory insufficiency and, eventually, fatal shock. These changes can be prevented to a degree by increasing dietary NaCl intake. Rats survive indefinitely on extra salt alone, but in dogs and most humans, the amount of supplementary salt needed is so large that it is almost impossible to prevent eventual collapse and death unless mineralocorticoid treatment is also instituted (see Clinical Box 19-4).
اثر آدرنالکتومی
در نارسایی آدرنال، +Na در ادرار از بین میرود. +K حفظ میشود و +K پلاسما بالا میرود. هنگامیکه نارسایی آدرنال به سرعت ایجاد میشود، مقدار +Na از دست رفته از ECF از مقدار دفع شده در ادرار بیشتر میشود، که نشان میدهد +Na نیز باید وارد سلولها شود. هنگامیکه هیپوفیز خلفی دست نخورده باشد، از دست دادن نمک از تلفات آب بیشتر میشود و +Na پلاسما کاهش مییابد (جدول 19-5). با این حال، حجم پلاسما نیز کاهش مییابد و منجر به افت فشار خون، نارسایی گردش خون و در نهایت شوک کشنده میشود. با افزایش دریافت نمک طعام در رژیم غذایی میتوان تا حدودی از این تغییرات جلوگیری کرد. موشها بهطور نامحدود تنها با نمک اضافی زنده میمانند، اما در سگها و بیشتر انسانها، مقدار نمک مکمل مورد نیاز آنقدر زیاد است که جلوگیری از فروپاشی و مرگ نهایی تقریباً غیرممکن است مگر اینکه درمان با مینرالوکورتیکوئید نیز انجام شود (به کادر بالینی 19-4 مراجعه کنید).
TABLE 19-5 Typical plasma electrolyte levels in normal humans and patients with adrenocortical diseases.
جدول 19-5 سطوح الکترولیت پلاسما معمولی در انسانهای عادی و بیماران مبتلا به بیماریهای قشر آدرنال.
CLINICAL BOX 19-4
Secondary Effects of Excess Mineralocorticoids
A prominent feature of prolonged mineralocorticoid excess (Table 20-5) is K+ depletion due to prolonged K+ diuresis. H+ is also lost in the urine. Na+ is retained initially, but the plasma Na+ is elevated only slightly if at all, because water is retained with the osmotically active sodium ions. Consequently, ECF volume is expanded and the blood pressure rises. When the ECF expansion passes a certain point, Na* excretion is usually increased in spite of the continued action of mineralocorticoids on the renal tubules. This escape phenomenon (Figure 19-20) is probably due to increased secretion of atrial natriuretic peptide (ANP) (see Chapter 38). Because of increased excretion of Na* when the ECF volume is expanded, mineralocorticoids do not produce edema in normal individuals and patients with hyperaldosteronism. However, escape may not occur in certain disease states, and in these situations, continued expansion of ECF volume leads to edema (see Chapters 37 and 38).
جعبه بالینی 19-4
اثرات ثانویه مینرالوکورتیکوئیدهای اضافی
یکی از ویژگیهای بارز افزایش طولانی مدت مینرالوکورتیکوئید (جدول 20-5) کاهش +K به دلیل دیورز طولانی مدت +K است. +H نیز در ادرار از بین میرود. +Na در ابتدا حفظ میشود، اما +Na پلاسما فقط کمیافزایش مییابد، زیرا آب با یونهای سدیم فعال اسمزی حفظ میشود. در نتیجه، حجم ECF افزایش مییابد و فشار خون افزایش مییابد. هنگامیکه انبساط ECF از یک نقطه خاص عبور میکند، علیرغم ادامه فعالیت مینرالوکورتیکوئیدها بر روی لولههای کلیوی، معمولاً دفع +Na افزایش مییابد. این پدیده فرار (شکل 19-20) احتمالاً به دلیل افزایش ترشح پپتید ناتریورتیک دهلیزی (ANP) است (به فصل 38 مراجعه کنید). به دلیل افزایش دفع +Na هنگام افزایش حجم ECF، مینرالوکورتیکوئیدها در افراد عادی و بیماران مبتلا به هیپرآلدوسترونیسم ادم ایجاد نمیکنند. با این حال، فرار ممکن است در برخی از حالات بیماری رخ ندهد، و در این شرایط، افزایش مداوم حجم ECF منجر به ادم میشود (به فصلهای 37 و 38 مراجعه کنید).

FIGURE 19-20 “Escape” from the sodium-retaining effect of desoxycorticosterone acetate (DOCA) in an adrenalectomized patient. ECF, extracellular fluid volume; PV, plasma volume; RCV, red cell volume; TBV, total blood volume. (Used with permission of EG Biglieri.)
شکل 19-20 “فرار” از اثر نگهدارنده سدیم دزوکسی کورتیکوسترون استات (DOCA) در یک بیمار آدرنالکتومیشده. ECF، حجم مایع خارج سلولی؛ PV، حجم پلاسما؛ RCV، حجم گلبول قرمز؛ TBV، حجم کل خون. (با اجازه EG Biglieri استفاده میشود.)
REGULATION OF ALDOSTERONE SECRETION
STIMULI
The principal conditions that increase aldosterone secretion are summarized in Table 19-6. Some of them also increase glucocorticoid secretion; others selectively affect the output of aldosterone. The primary regulatory factors involved are ACTH from the pituitary, renin from the kidney via angiotensin II, and a direct stimulatory effect on the adrenal cortex of a rise in plasma K+ concentration.
تنظیم ترشح آلدوسترون
محرک
شرایط اصلی افزایش ترشح آلدوسترون در جدول 19-6 خلاصه شده است. برخی از آنها نیز ترشح گلوکوکورتیکوئید را افزایش میدهند. برخی دیگر به طور انتخابی بر خروجی آلدوسترون تأثیر میگذارند. عوامل تنظیم کننده اولیه درگیر عبارتند از ACTH از هیپوفیز، رنین از کلیه از طریق آنژیوتانسین II، و یک اثر تحریکی مستقیم بر روی قشر آدرنال ناشی از افزایش غلظت +K پلاسما.
TABLE 19-6 Conditions that increase aldosterone secretion.
جدول 19-6 شرایطی که باعث افزایش ترشح آلدوسترون میشود.

EFFECT OF ACTH
When first administered, ACTH stimulates the output of aldosterone as well as that of glucocorticoids and sex hormones. Although the amount of ACTH required to increase aldosterone output is somewhat greater than the amount that stimulates maximal glucocorticoid secretion (Figure 19-21), it is well within the range of endogenous ACTH secretion. The effect is transient, and even if ACTH secretion remains elevated, aldosterone output declines in 1 or 2 days. On the other hand, the output of the mineralocorticoid deoxycorticosterone remains elevated. The decline in aldosterone output is partly due to decreased renin secretion secondary to hypervolemia, but it is possible that some other factor also decreases the conversion of corticosterone to aldosterone. After hypophysectomy, the basal rate of aldosterone secretion is normal. The increase normally produced by surgical and other stresses is absent, but the increase produced by dietary salt restriction is unaffected for some time. Later on, atrophy of the zona glomerulosa complicates the picture in long-standing hypopituitarism, and this may lead to salt loss and hypoaldosteronism.
اثر ACTH
هنگامیکه ACTH برای اولین بار تجویز میشود، خروجی آلدوسترون و همچنین گلوکوکورتیکوئیدها و هورمونهای جنسی را تحریک میکند. اگرچه مقدار ACTH مورد نیاز برای افزایش برون ده آلدوسترون تا حدودی بیشتر از مقداری است که حداکثر ترشح گلوکوکورتیکوئید را تحریک میکند (شکل 19-21)، اما به خوبی در محدوده ترشح ACTH درون زا قرار دارد. این اثر گذرا است و حتی اگر ترشح ACTH همچنان بالا بماند، خروجی آلدوسترون در 1 یا 2 روز کاهش مییابد. از سوی دیگر، خروجی مینرالوکورتیکوئید دئوکسی کورتیکوسترون بالا باقی میماند. کاهش برون ده آلدوسترون تا حدی به دلیل کاهش ترشح رنین ثانویه به هیپرولمیاست، اما ممکن است عامل دیگری نیز باعث کاهش تبدیل کورتیکوسترون به آلدوسترون شود. پس از هیپوفیزکتومی، میزان پایه ترشح آلدوسترون طبیعی است. افزایشی که معمولاً در اثر فشارهای جراحی و سایر استرسها ایجاد میشود وجود ندارد، اما افزایش تولید شده توسط محدودیت نمک رژیم غذایی برای مدتی تحت تأثیر قرار نمیگیرد. بعداً، آتروفی زونا گلومرولوزا تصویر را در کمکاری هیپوفیز طولانی مدت پیچیده میکند و این ممکن است منجر به از دست دادن نمک و هیپوآلدوسترونیسم شود.

FIGURE 19-21 Changes in adrenal venous output of steroids produced by ACTH in nephrectomized hypophysectomized dogs. ACTH, adrenocorticotropic hormone.
شکل 19-21 تغییرات در برون ده وریدی آدرنال استروئیدهای تولید شده توسط ACTH در سگهای نفرکتومیشده هیپوفیزکتومیشده. ACTH، هورمون آدرنوکورتیکوتروپیک.
Normally, glucocorticoid treatment does not suppress aldosterone secretion. However, an interesting recently described syndrome is glucocorticoid- remediable aldosteronism (GRA). This is an autosomal dominant disorder in which the increase in aldosterone secretion produced by ACTH is no longer transient. The hypersecretion of aldosterone and the accompanying hypertension are remedied when ACTH secretion is suppressed by administering glucocorticoids. The genes encoding aldosterone synthase and 11ẞ-hydroxylase are 95% identical and are close together on chromosome 8. In individuals with GRA, there is unequal crossing over so that the 5’regulatory region of the 11ẞ- hydroxylase gene is fused to the coding region of the aldosterone synthase gene. The product of this hybrid gene is an ACTH-sensitive aldosterone synthase.
به طور معمول، درمان با گلوکوکورتیکوئید ترشح آلدوسترون را سرکوب نمیکند. با این حال، یک سندرم جالب که اخیراً توصیف شده است، آلدوسترونیسم قابل درمان با گلوکوکورتیکوئید (GRA) است. این یک اختلال اتوزومال غالب است که در آن افزایش ترشح آلدوسترون تولید شده توسط ACTH دیگر گذرا نیست. ترشح بیش از حد آلدوسترون و پرفشاری خون همراه با سرکوب ترشح ACTH با تجویز گلوکوکورتیکوئیدها برطرف میشود. ژنهای کد کننده آلدوسترون سنتاز و 11-هیدروکسیلاز 95 درصد یکسان هستند و در کروموزوم 8 به هم نزدیک هستند. در افراد مبتلا به GRA، تلاقی نابرابر وجود دارد به طوری که ناحیه تنظیم کننده 5 ژن 11-هیدروکسیلاز به ناحیه کد کننده سنتز ژن آلدوسترون ترکیب میشود. محصول این ژن هیبریدی یک آلدوسترون سنتاز حساس به ACTH است.
EFFECTS OF ANGIOTENSIN II & RENIN
The octapeptide angiotensin II is formed in the body from angiotensin I, which is liberated by the action of renin on circulating angiotensinogen (see Chapter 38). Injections of angiotensin II stimulate adrenocortical secretion and, in small doses, affect primarily the secretion of aldosterone. The sites of action of angiotensin II are both early and late in the steroid biosynthetic pathway. The early action is on the conversion of cholesterol to pregnenolone, and the late action is on the conversion of corticosterone to aldosterone (Figure 19-8). Angiotensin II does not increase the secretion of deoxycorticosterone, which is controlled by ACTH.
اثرات آنژیوتانسین II و رنین
اکتاپپتید آنژیوتانسین II در بدن از آنژیوتانسین I تشکیل میشود که با اثر رنین بر روی آنژیوتانسینوژن در گردش آزاد میشود (به فصل 38 مراجعه کنید). تزریق آنژیوتانسین II ترشح قشر آدرنال را تحریک میکند و در دوزهای کوچک عمدتاً بر ترشح آلدوسترون تأثیر میگذارد. محلهای اثر آنژیوتانسین II هم در مراحل اولیه و هم در اواخر مسیر بیوسنتزی استروئیدی است. اقدام اولیه بر روی تبدیل کلسترول به پرگننولون و اقدام دیرهنگام بر روی تبدیل کورتیکوسترون به آلدوسترون است (شکل 19-8). آنژیوتانسین II باعث افزایش ترشح دئوکسی کورتیکوسترون نمیشود که توسط ACTH کنترل میشود.
Renin is secreted from the juxtaglomerular cells that surround the renal afferent arterioles as they enter the glomeruli (see Chapter 38). Aldosterone secretion is regulated via the renin-angiotensin system in a feedback manner (Figure 19-22). A drop in ECF volume or intra-arterial vascular volume leads to a reflex increase in renal nerve discharge and decreases renal arterial pressure. Both changes increase renin secretion, and the angiotensin II formed by the action of renin increases the rate of secretion of aldosterone. The aldosterone causes Na+ and, secondarily, water retention, expanding ECF volume, and shutting off the stimulus that initiated increased renin secretion.
رنین از سلولهای juxtaglomerular که شریانهای آوران کلیه را احاطه کرده اند، هنگام ورود به گلومرولها ترشح میشود (به فصل 38 مراجعه کنید). ترشح آلدوسترون از طریق سیستم رنین-آنژیوتانسین به روش بازخوردی تنظیم میشود (شکل 19-22). کاهش حجم ECF یا حجم عروق داخل شریانی منجر به افزایش رفلکس در تخلیه عصب کلیوی و کاهش فشار شریانی کلیه میشود. هر دو تغییر باعث افزایش ترشح رنین میشوند و آنژیوتانسین II که در اثر عمل رنین ایجاد میشود، سرعت ترشح آلدوسترون را افزایش میدهد. آلدوسترون باعث +Na و در مرحله دوم، احتباس آب، افزایش حجم ECF و قطع محرکی میشود که باعث افزایش ترشح رنین میشود.
FIGURE 19-22 Feedback mechanism regulating aldosterone secretion. The dashed arrow indicates inhibition.
شکل 19-22 مکانیسم بازخورد تنظیم کننده ترشح آلدوسترون. فلش چین نشان دهنده بازداری است.
Hemorrhage stimulates ACTH and renin secretion. Like hemorrhage, standing and constriction of the thoracic inferior vena cava decrease intrarenal arterial pressure. Dietary sodium restriction also increases aldosterone secretion via the renin-angiotensin system (Figure 19-23). Such restriction reduces ECF volume, but aldosterone and renin secretion are increased before any consistent decrease in blood pressure takes place. Consequently, the initial increase in renin secretion produced by dietary sodium restriction is probably due to a reflex increase in the activity of the renal nerves. The increase in circulating angiotensin II produced salt depletion upregulates the angiotensin II receptors in the adrenal cortex and hence increases the response to angiotensin II, whereas it down-regulates the angiotensin II receptors in the blood vessels.
خونریزی باعث تحریک ترشح ACTH و رنین میشود. مانند خونریزی، ایستادن و انقباض ورید اجوف تحتانی قفسه سینه باعث کاهش فشار شریانی داخل کلیه میشود. محدودیت سدیم در رژیم غذایی نیز ترشح آلدوسترون را از طریق سیستم رنین-آنژیوتانسین افزایش میدهد (شکل 19-23). چنین محدودیتی حجم ECF را کاهش میدهد، اما ترشح آلدوسترون و رنین قبل از کاهش مداوم فشار خون افزایش مییابد. در نتیجه، افزایش اولیه ترشح رنین تولید شده توسط محدودیت سدیم در رژیم غذایی احتمالاً به دلیل افزایش رفلکس در فعالیت اعصاب کلیوی است. افزایش کاهش نمک تولید شده در گردش آنژیوتانسین II، گیرندههای آنژیوتانسین II را در قشر آدرنال تنظیم میکند و در نتیجه پاسخ به آنژیوتانسین II را افزایش میدهد، در حالی که گیرندههای آنژیوتانسین II را در رگهای خونی کاهش میدهد.

FIGURE 19-23 Effect of low-, normal-, and high-sodium diets on sodium metabolism and plasma renin activity, aldosterone, vasopressin, and ANP in normal humans. (Data from Sagnella GA, et al: Plasma atrial natriuretic peptide: Its relationship to changes in sodium in-take, plasma renin activity, and aldosterone in man. Clin Sci 1987; 72:25.)
شکل 19-23 اثر رژیمهای کم، نرمال و پر سدیم بر متابولیسم سدیم و فعالیت رنین پلاسما، آلدوسترون، وازوپرسین و ANP در انسانهای عادی. (دادههای Sagnella GA و همکاران: پپتید ناتریورتیک دهلیزی پلاسما: ارتباط آن با تغییرات سدیم دریافتی، فعالیت رنین پلاسما و آلدوسترون در انسان. Clin Sci 1987؛ 72:25.)
ELECTROLYTES & OTHER FACTORS
An acute decline in plasma Na+ of about 20 mEq/L stimulates aldosterone secretion, but changes of this magnitude are rare. However, the plasma K+ level need increase only 1 mEq/L to stimulate aldosterone secretion, and transient increases of this magnitude may occur after a meal, particularly if it is rich in K+. Like angiotensin II, K+ stimulates the conversion of cholesterol to pregnenolone and the conversion of deoxycorticosterone to aldosterone. It appears to act by depolarizing the cell, which opens voltage-gated Ca2+ channels, increasing intracellular Ca2+. The sensitivity of the zona glomerulosa to angiotensin II and consequently to a low-sodium diet is decreased by a low-potassium diet.
الکترولیتها و سایر عوامل
کاهش حاد سدیم پلاسما به میزان 20 میلی اکی والان در لیتر باعث تحریک ترشح آلدوسترون میشود، اما تغییرات در این بزرگی نادر است. با این حال، سطح +K پلاسما برای تحریک ترشح آلدوسترون تنها به 1 میلی اکی والان در لیتر نیاز دارد و افزایش گذرا در این مقدار ممکن است بعد از غذا رخ دهد، به خصوص اگر غنی از پتاسیم باشد. مانند آنژیوتانسین +II، K تبدیل کلسترول به پرگننولون و تبدیل دئوکسی کورتیکوسترون به آلدوسترون را تحریک میکند. به نظر میرسد که با دپلاریز کردن سلول عمل میکند، که کانالهای +Ca2 دارای ولتاژ را باز میکند و +Ca2 داخل سلولی را افزایش میدهد. حساسیت زونا گلومرولوزا به آنژیوتانسین II و در نتیجه به رژیم غذایی کم سدیم با رژیم غذایی کم پتاسیم کاهش مییابد.
In normal individuals, plasma aldosterone concentrations increase during the portion of the day that the individual is carrying on activities in the upright position. This increase is due to a decrease in the rate of removal of aldosterone from the circulation by the liver and an increase in aldosterone secretion due to a postural increase in renin secretion. Individuals who are confined to bed show a circadian rhythm of aldosterone and renin secretion, with the highest values in the early morning before awakening. Atrial natriuretic peptide (ANP) inhibits renin secretion and decreases the responsiveness of the zona glomerulosa to angiotensin II (see Chapter 38). The mechanisms by which ACTH, angiotensin II, and K+ stimulate aldosterone secretion are summarized in Table 19-7.
در افراد عادی، غلظت آلدوسترون پلاسما در بخشی از روز که فرد در وضعیت عمودی فعالیت میکند، افزایش مییابد. این افزایش به دلیل کاهش سرعت حذف آلدوسترون از گردش خون توسط کبد و افزایش ترشح آلدوسترون به دلیل افزایش وضعیتی در ترشح رنین است. افرادی که در رختخواب هستند، ریتم شبانه روزی ترشح آلدوسترون و رنین را نشان میدهند که بالاترین مقدار آن در اوایل صبح قبل از بیدار شدن است. پپتید ناتریورتیک دهلیزی (ANP) ترشح رنین را مهار میکند و پاسخ زونا گلومرولوزا به آنژیوتانسین II را کاهش میدهد (به فصل 38 مراجعه کنید). مکانیسمهایی که ACTH، آنژیوتانسین II و +K ترشح آلدوسترون را تحریک میکنند در جدول 19-7 خلاصه شده است.
TABLE 19-7 Second messengers involved in the regulation of aldosterone secretion.
جدول 19-7 پیام رسانهای دوم که در تنظیم ترشح آلدوسترون نقش دارند.
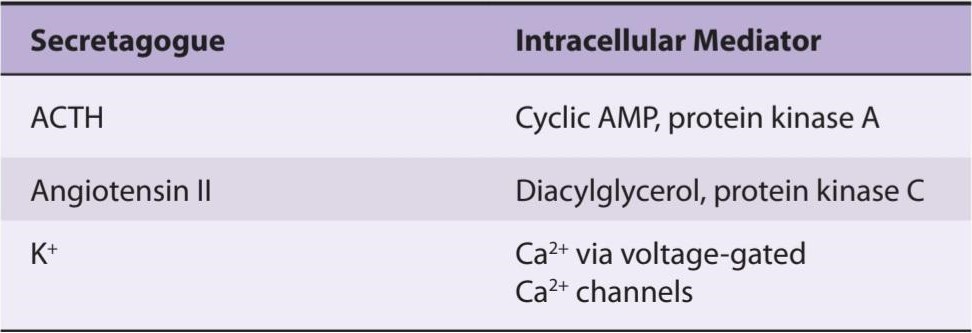
ROLE OF MINERALOCORTICOIDS IN THE REGULATION OF SALT BALANCE
Variations in aldosterone secretion is only one of many factors affecting Na+ excretion. Other major factors include the glomerular filtration rate, ANP, the presence or absence of osmotic diuresis, and changes in tubular reabsorption of Na+ independent of aldosterone. It takes some time for aldosterone to act. When one rises from the supine to the standing position, aldosterone secretion increases and Na+ is retained from the urine. However, the decrease in Na+ excretion develops too rapidly to be explained solely by increased aldosterone secretion. The primary function of the aldosterone-secreting mechanism is the defense of intravascular volume, but it is only one of the homeostatic mechanisms involved in this regulation.
نقش مینرالوکورتیکوئیدها در تنظیم تعادل نمک
تغییرات در ترشح آلدوسترون تنها یکی از عوامل موثر بر دفع +Na است. سایر عوامل اصلی عبارتند از: نرخ فیلتراسیون گلومرولی، ANP، وجود یا عدم وجود دیورز اسمزی، و تغییرات در بازجذب لولهای +Na مستقل از آلدوسترون. مدتی طول میکشد تا آلدوسترون عمل کند. هنگامیکه فرد از حالت خوابیده به پشت به حالت ایستاده بالا میرود، ترشح آلدوسترون افزایش مییابد و +Na از ادرار باقی میماند. با این حال، کاهش دفع +Na خیلی سریع ایجاد میشود که نمیتوان آن را صرفاً با افزایش ترشح آلدوسترون توضیح داد. عملکرد اولیه مکانیسم ترشح آلدوسترون، دفاع از حجم داخل عروقی است، اما این تنها یکی از مکانیسمهای هموستاتیکی است که در این تنظیم نقش دارد.
SUMMARY OF THE EFFECTS OF ADRENOCORTICAL HYPER- & HYPOFUNCTION IN HUMANS
Recapitulating the manifestations of excess and deficiency of the adrenocortical hormones in humans is a convenient way to summarize the multiple and complex actions of these steroids. A characteristic clinical syndrome is associated with excess secretion of each of the types of hormones.
خلاصه ای از اثرات بیش از حد و کم کاری غده آدرنوکورتیکال در انسان
مروری بر تظاهرات بیش از حد و کمبود هورمونهای قشر آدرنال در انسان، راه مناسبی برای خلاصه کردن اعمال متعدد و پیچیده این استروئیدها است. یک سندرم بالینی مشخصه با ترشح بیش از حد هر یک از انواع هورمونها همراه است.
Excess androgen secretion causes masculinization (adrenogenital syndrome) and precocious pseudopuberty or female pseudohermaphroditism.
ترشح بیش از حد آندروژن باعث مردانه شدن (سندرم آدرنوژنیتال) و بلوغ کاذب زودرس یا شبه هرمافرودیتیسم زنانه میشود.
Excess glucocorticoid secretion produces a moon-faced, plethoric appearance, with trunk obesity, purple abdominal striae, hypertension, osteoporosis, protein depletion, mental abnormalities and, frequently, diabetes mellitus (Cushing syndrome). Excess mineralocorticoid secretion leads to K+ depletion and Na+ retention, usually without edema but with weakness, hypertension, tetany, polyuria, and hypokalemic alkalosis (hyperaldosteronism). This condition may be due to primary adrenal disease (primary hyperaldosteronism; Conn syndrome) such as an adenoma of the zona glomerulosa, unilateral or bilateral adrenal hyperplasia, adrenal carcinoma, or by GRA. In patients with primary hyperaldosteronism, renin secretion is depressed. Secondary hyperaldosteronism with high plasma renin activity is caused by cirrhosis, heart failure, and nephrosis. Increased renin secretion is also found in individuals with the salt-losing form of the adrenogenital syndrome (see above), because their ECF volume is low. In patients with elevated renin secretion due to renal artery constriction, aldosterone secretion is increased; in those in whom renin secretion is not elevated, aldosterone secretion is normal. The relationship of aldosterone to hypertension is discussed in Chapter 32.
ترشح بیش از حد گلوکوکورتیکوئید، ظاهری پر از ماه، با چاقی تنه، استریاهای بنفش شکمی، فشار خون بالا، پوکی استخوان، کاهش پروتئین، ناهنجاریهای ذهنی و اغلب دیابت شیرین (سندرم کوشینگ) ایجاد میکند. ترشح بیش از حد مینرالوکورتیکوئید منجر به کاهش پتاسیم و احتباس +Na میشود، معمولاً بدون ادم اما با ضعف، فشار خون بالا، کزاز، پلی اوری و آلکالوز هیپوکالمیک (هیپرآلدوسترونیسم). این وضعیت ممکن است به دلیل بیماری اولیه آدرنال (هیپرآلدوسترونیسم اولیه، سندرم کان) مانند آدنوم زونا گلومرولوزا، هیپرپلازی یک طرفه یا دو طرفه آدرنال، کارسینوم آدرنال یا توسط GRA باشد. در بیماران مبتلا به هیپرآلدوسترونیسم اولیه، ترشح رنین کاهش مییابد. هیپرآلدوسترونیسم ثانویه با فعالیت رنین بالا در پلاسما ناشی از سیروز، نارسایی قلبی و نفروز است. افزایش ترشح رنین نیز در افراد مبتلا به سندرم آدرنوژنیتال از دست دادن نمک مشاهده میشود (به بالا مراجعه کنید)، زیرا حجم ECF آنها کم است. در بیماران با ترشح رنین بالا به دلیل انقباض شریان کلیوی، ترشح آلدوسترون افزایش مییابد. در کسانی که ترشح رنین در آنها بالا نیست، ترشح آلدوسترون طبیعی است. رابطه آلدوسترون با فشار خون بالا در فصل 32 مورد بحث قرار گرفته است.
Primary adrenal insufficiency due to disease processes that destroy the adrenal cortex is called Addison disease. The condition used to be a relatively common complication of tuberculosis, but now it is usually due to autoimmune inflammation of the adrenal. Patients lose weight, are tired, and become chronically hypotensive. They have small hearts, probably because the hypotension decreases the work of the heart. Eventually, severe hypotension and shock (addisonian crisis) develop. This is due not only to mineralocorticoid deficiency but to glucocorticoid deficiency as well. Fasting causes fatal hypoglycemia, and any stress causes collapse. Water is retained, and there is always the danger of water intoxication. Circulating ACTH levels are elevated. The diffuse tanning of the skin and the spotty pigmentation characteristic of chronic glucocorticoid deficiency are due, at least in part, to the melanocyte- stimulating hormone (MSH) activity of the ACTH in the blood. Pigmentation of skin creases on the hands and the gums are common. Minor menstrual abnormalities occur in women, but the deficiency of adrenal sex hormones usually has little effect in the presence of normal testes or ovaries.
نارسایی اولیه آدرنال ناشی از فرآیندهای بیماری که باعث تخریب قشر آدرنال میشود، بیماری آدیسون نامیده میشود. این بیماری قبلاً یک عارضه نسبتاً شایع سل بود، اما اکنون معمولاً به دلیل التهاب خود ایمنی غده فوق کلیوی است. بیماران وزن کم میکنند، خسته هستند و به طور مزمن دچار افت فشار خون میشوند. آنها قلبهای کوچکی دارند، احتمالاً به این دلیل که افت فشار خون کار قلب را کاهش میدهد. در نهایت، افت فشار خون شدید و شوک (بحران آدیسونی) ایجاد میشود. این نه تنها به دلیل کمبود مینرالوکورتیکوئید بلکه به دلیل کمبود گلوکوکورتیکوئید نیز میباشد. روزه باعث هیپوگلیسمیکشنده میشود و هر استرسی باعث فروپاشی میشود. آب حفظ میشود و همیشه خطر مسمومیت با آب وجود دارد. سطح ACTH در گردش افزایش یافته است. برنزه شدن پوست و رنگدانههای خالدار مشخصه کمبود مزمن گلوکوکورتیکوئید، حداقل تا حدی به دلیل فعالیت هورمون محرک ملانوسیت (MSH) ACTH در خون است. رنگدانه شدن چینهای پوستی روی دستها و لثهها شایع است. ناهنجاریهای جزئی قاعدگی در زنان رخ میدهد، اما کمبود هورمونهای جنسی آدرنال معمولاً در حضور بیضهها یا تخمدانهای طبیعی تأثیر کمیدارد.
Secondary adrenal insufficiency is caused by pituitary diseases that decrease ACTH secretion, and tertiary adrenal insufficiency is caused by hypothalamic disorders disrupting CRH secretion. Both are usually milder than primary adrenal insufficiency because electrolyte metabolism is affected to a lesser degree. In addition, there is no pigmentation because in both of these conditions, plasma ACTH is low, not high.
نارسایی ثانویه آدرنال ناشی از بیماریهای هیپوفیز است که باعث کاهش ترشح ACTH میشود و نارسایی ثانویه آدرنال به دلیل اختلالات هیپوتالاموس که ترشح CRH را مختل میکند، ایجاد میشود. هر دو معمولاً خفیف تر از نارسایی اولیه آدرنال هستند زیرا متابولیسم الکترولیتها به میزان کمتری تحت تأثیر قرار میگیرد. علاوه بر این، هیچ رنگدانه ای وجود ندارد زیرا در هر دوی این شرایط، ACTH پلاسما کم است، نه زیاد.
Cases of isolated aldosterone deficiency have also been reported in patients with renal disease and a low circulating renin level (hyporeninemic hypoaldosteronism). In addition, pseudohypoaldosteronism is produced when there is resistance to the action of aldosterone. Patients with these syndromes have marked hyperkalemia, salt wasting, and hypotension, and they may develop metabolic acidosis.
مواردی از کمبود جدا شده آلدوسترون نیز در بیماران مبتلا به بیماری کلیوی و سطح رنین در گردش پایین (هیپوآلدوسترونیسم هیپورنیمیک) گزارش شده است. علاوه بر این، هیپوآلدوسترونیسم کاذب زمانی ایجاد میشود که مقاومت در برابر عمل آلدوسترون وجود داشته باشد. بیماران مبتلا به این سندرمها هیپرکالمیمشخص، هدر رفتن نمک و افت فشار خون دارند و ممکن است دچار اسیدوز متابولیک شوند.
CHAPTER SUMMARY
خلاصه فصل
• The adrenal gland consists of the adrenal medulla that secretes dopamine and the catecholamines epinephrine and norepinephrine, and the adrenal cortex that secretes steroid hormones.
• غده آدرنال شامل بصل الکلی است که دوپامین و کاتکول آمینهای اپی نفرین و نوراپی نفرین ترشح میکند و قشر آدرنال که هورمونهای استروئیدی ترشح میکند.
• Norepinephrine and epinephrine act on two classes of receptors, a- and ẞ- adrenergic receptors, and exert metabolic effects that include glycogenolysis in liver and skeletal muscle, mobilization of FFA, increased plasma lactate, and stimulation of the metabolic rate.
• نوراپی نفرین و اپی نفرین روی دو دسته از گیرندههای گیرندههای a- و ẞ- آدرنرژیک عمل میکنند و اثرات متابولیکی از جمله گلیکوژنولیز در کبد و عضله اسکلتی، تحرک FFA، افزایش لاکتات پلاسما و تحریک سرعت متابولیک را اعمال میکنند.
• The hormones of the adrenal cortex are derivatives of cholesterol and include the mineralocorticoid aldosterone, the glucocorticoids cortisol and corticosterone, and the androgens dehydroepiandrosterone (DHEA) and androstenedione.
• هورمونهای قشر آدرنال مشتقات کلسترول هستند و شامل مینرالوکورتیکوئید آلدوسترون، گلوکوکورتیکوئیدهای کورتیزول و کورتیکوسترون و آندروژنهای دهیدرواپی آندروسترون (DHEA) و آندروستندیون میباشند.
• Androgens are the hormones that exert masculinizing effects, and they promote protein anabolism and growth. The adrenal androgen androstenedione is converted to testosterone and to estrogens (aromatized) in fat and other peripheral tissues. This is an important source of estrogens in men and postmenopausal women.
• آندروژنها هورمونهایی هستند که اثرات مردانه دارند و باعث آنابولیسم و رشد پروتئین میشوند. آندروژن آدرنال آندروستندیون در چربی و سایر بافتهای محیطی به تستوسترون و به استروژن (معطر شده) تبدیل میشود. این منبع مهم استروژن در مردان و زنان یائسه است.
• The mineralocorticoid aldosterone has effects on Na+ and K+ excretion and glucocorticoids affect glucose and protein metabolism.
• مینرالوکورتیکوئید آلدوسترون بر دفع +Na و +K و گلوکوکورتیکوئیدها بر متابولیسم گلوکز و پروتئین تأثیر میگذارد.
• Glucocorticoid secretion is dependent on ACTH from the anterior pituitary and is increased by stress. Angiotensin II increases the secretion of aldosterone.
• ترشح گلوکوکورتیکوئید به ACTH از هیپوفیز قدامیوابسته است و در اثر استرس افزایش مییابد. آنژیوتانسین II باعث افزایش ترشح آلدوسترون میشود.
MULTIPLE-CHOICE QUESTIONS
For all questions, select the single best answer unless otherwise directed.
سوالات چند گزینه ای
برای همه سؤالات، بهترین پاسخ را انتخاب کنید، مگر اینکه دستور دیگری داده شود.
1. Which of the following is produced only by large amounts of glucocorticoids?
A. Normal responsiveness of fat depots to norepinephrine
B. Maintenance of normal vascular reactivity
C. Increased excretion of a water load
D. Inhibition of the inflammatory response
E. Inhibition of ACTH secretion
1. کدام یک از موارد زیر فقط توسط مقادیر زیاد گلوکوکورتیکوئیدها تولید میشود؟
الف. پاسخ طبیعی ذخایر چربی به نوراپی نفرین
ب. حفظ واکنش پذیری عروقی طبیعی
ج. افزایش دفع یک بار آبی
د. مهار پاسخ التهابی
ی. مهار ترشح ACTH
2. Which of the following are incorrectly paired?
A. Gluconeogenesis: Cortisol
B. Free fatty acid mobilization: Dehydroepiandrosterone
C. Muscle glycogenolysis: Epinephrine
D. Kaliuresis : Aldosterone
E. Hepatic glycogenesis: Insulin
2- کدام یک از موارد زیر به اشتباه جفت شده اند؟
الف. گلوکونئوژنز: کورتیزول
ب. بسیج اسیدهای چرب آزاد: دهیدرواپی آندروسترون
ج. گلیکوژنولیز عضلانی: اپی نفرین
د. Kaliuresis: آلدوسترون
ی. گلیکوژنز کبدی: انسولین
3. Which of the following hormones has the shortest plasma half-life?
A. Corticosterone
B. Renin
C. Dehydroepiandrosterone
D. Aldosterone
E. Norepinephrine
3- کدام یک از هورمونهای زیر نیمه عمر پلاسمایی کمتری دارد؟
الف. کورتیکوسترون
ب. رنین
ج. دهیدرواپی آندروسترون
د. آلدوسترون
ی. نوراپی نفرین
4. Mole for mole, which of the following has the greatest effect on Na* excretion?
A. Progesterone
B. Cortisol
C. Vasopressin
D. Aldosterone
E. Dehydroepiandrosterone
4. خال برای خال، کدام یک از موارد زیر بیشترین تأثیر را در دفع +Na دارد؟
الف. پروژسترون
ب. کورتیزول
ج. وازوپرسین
د. آلدوسترون
ی. دهیدرواپی آندروسترون
5. Mole for mole, which of the following has the greatest effect on plasma osmolality?
A. Progesterone
B. Cortisol
C. Vasopressin
D. Aldosterone
E. Dehydroepiandrosterone
5. خال برای خال، کدام یک از موارد زیر بیشترین تأثیر را بر اسمولالیته پلاسما دارد؟
الف. پروژسترون
ب. کورتیزول
ج. وازوپرسین
د. آلدوسترون
ی. دهیدرواپی آندروسترون
6. The secretion of which of the following would be least affected by a decrease in extracellular fluid volume?
A. CRH
B. Arginine vasopressin
C. Dehydroepiandrosterone
D. Estrogens
E. Aldosterone
6. ترشح کدام یک از موارد زیر کمتر تحت تأثیر کاهش حجم مایع خارج سلولی قرار میگیرد؟
الف. CRH
ب. وازوپرسین آرژنین
ج. دهیدرواپی آندروسترون
د. استروژنها
ی. آلدوسترون
7. A young man presents with a blood pressure of 175/110 mm Hg. He is found to have a high circulating aldosterone but a low circulating cortisol. Glucocorticoid treatment lowers his circulating aldosterone and lowers his blood pressure to 140/85 mm Hg. He probably has an abnormal
A. 17α-hydroxylase.
B. 21ẞ-hydroxylase.
C. 3ẞ-hydroxysteroid dehydrogenase.
D. aldosterone synthase.
E. cholesterol desmolase.
7. مرد جوانی با فشار خون 175/110 میلی متر جیوه مراجعه میکند. مشخص شد که او دارای آلدوسترون در گردش بالا اما کورتیزول در گردش پایین است. درمان با گلوکوکورتیکوئید آلدوسترون در گردش او را کاهش میدهد و فشار خون او را به 140/85 میلی متر جیوه کاهش میدهد. او احتمالا یک بیماری غیر طبیعی دارد
الف. 17α-هیدروکسیلاز.
ب. 21ẞ-هیدروکسیلاز.
ج. 3ẞ-هیدروکسی استروئید دهیدروژناز.
د. آلدوسترون سنتاز.
ی. کلسترول دسمولاز.
8. A 32-year-old woman presents with a blood pressure of 155/96 mm Hg. In response to questioning, she admits that she loves licorice and eats some at least three times a week. She probably has a low level of
A. type 2 11ẞ-hydroxysteroid dehydrogenase activity.
B. ACTH.
C. 11ẞ-hydroxylase activity.
D. glucuronyl transferase.
E. norepinephrine.
8. خانمی32 ساله با فشار خون 155/96 میلی متر جیوه مراجعه میکند. او در پاسخ به سوال اعتراف میکند که عاشق شیرین بیان است و حداقل سه بار در هفته مقداری از آن را میخورد. احتمالا سطح پایینی داره
الف. نوع 2 فعالیت 11ẞ-هیدروکسی استروئید دهیدروژناز.
ب. ACTH.
ج. فعالیت 11-هیدروکسیلاز.
د. گلوکورونیل ترانسفراز.
ی. نوراپی نفرین.
9. In its action in cells, aldosterone
A. increases transport of ENaCs from the cytoplasm to the cell membrane.
B. does not act on the cell membrane.
C. binds to a receptor excluded from the nucleus.
D. may activate a heat shock protein.
E. also binds to glucocorticoid receptors.
9. در عمل خود در سلولها، آلدوسترون
الف. انتقال ENaCs را از سیتوپلاسم به غشای سلولی افزایش میدهد.
ب. روی غشای سلولی اثر نمیکند.
ج. به یک گیرنده خارج از هسته متصل میشود.
د. ممکن است پروتئین شوک حرارتی را فعال کند.
ی. همچنین به گیرندههای گلوکوکورتیکوئید متصل میشود.
کلیک کنید «منابع»
CHAPTER RESOURCES
Goldstein JL, Brown MS: The cholesterol quartet. Science 2001;292:1510.
Goodman HM (editor): Handbook of Physiology, Section 7: The Endocrine System. Oxford University Press, 2000.
Larsen PR, Kronenberg HM, Melmed S, et al. (editors). Williams Textbook of Endocrinology, 9th ed. Saunders, 2003.
Stocco DM: A review of the characteristics of the protein required for the acute regulation of steroid hormone biosynthesis: The case for the steroidogenic acute regulatory (StAR) protein. Proc Soc Exp Biol Med 1998;217:123.
White PC: Disorders of aldosterone biosynthesis and action. N Engl J Med 1994;331:250.
🚀 با ما همراه شوید!
تازهترین مطالب و آموزشهای مغز و اعصاب را از دست ندهید. با فالو کردن کانال تلگرام آیندهنگاران مغز، از ما حمایت کنید!

 ورود/ ثبت نام با جیمیل
ورود/ ثبت نام با جیمیل
